Comparison of Drought and Heat Resistance Strategies among Six Populations of Solanum chilense and Two Cultivars of Solanum lycopersicum

 , and
, and
Abstract
:1. Introduction
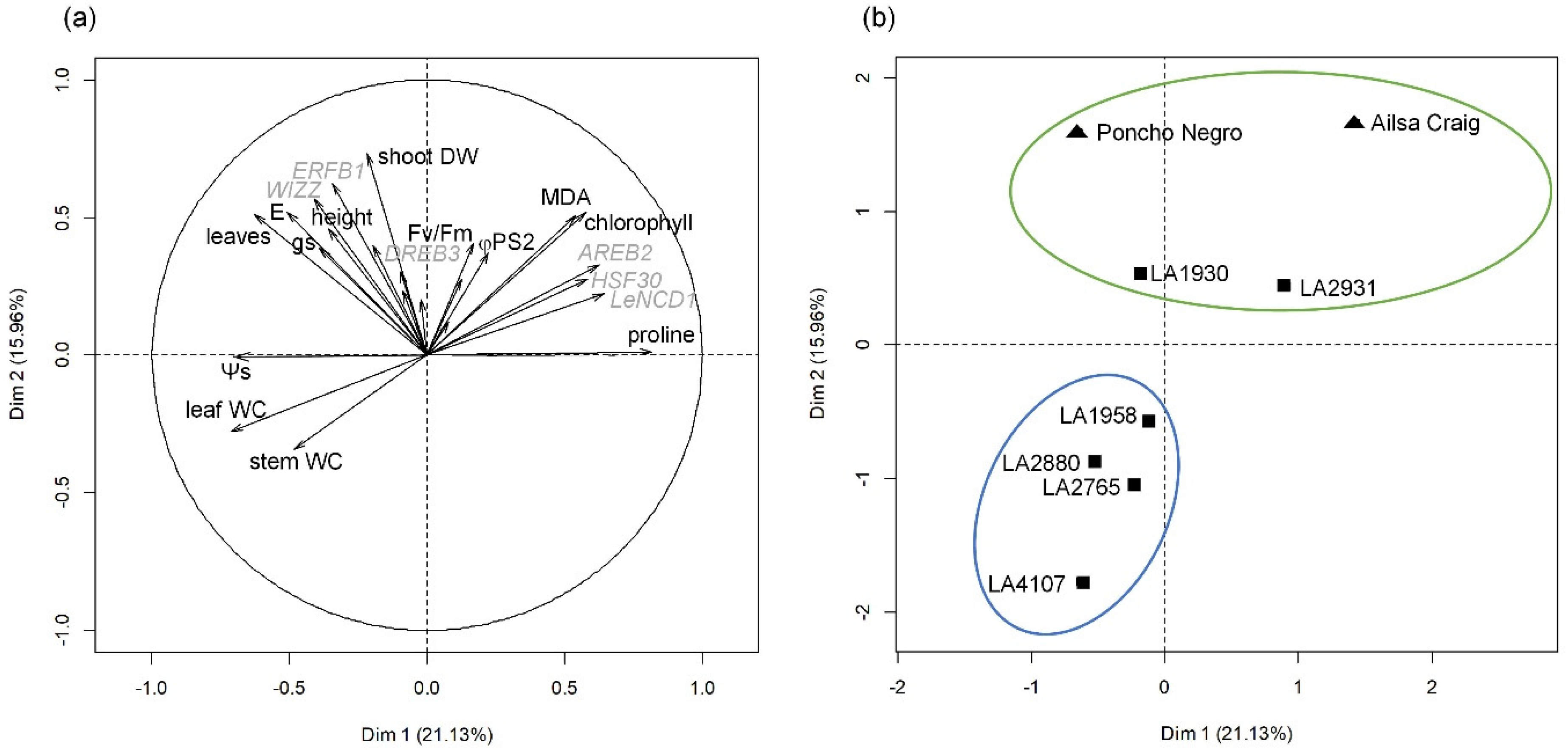
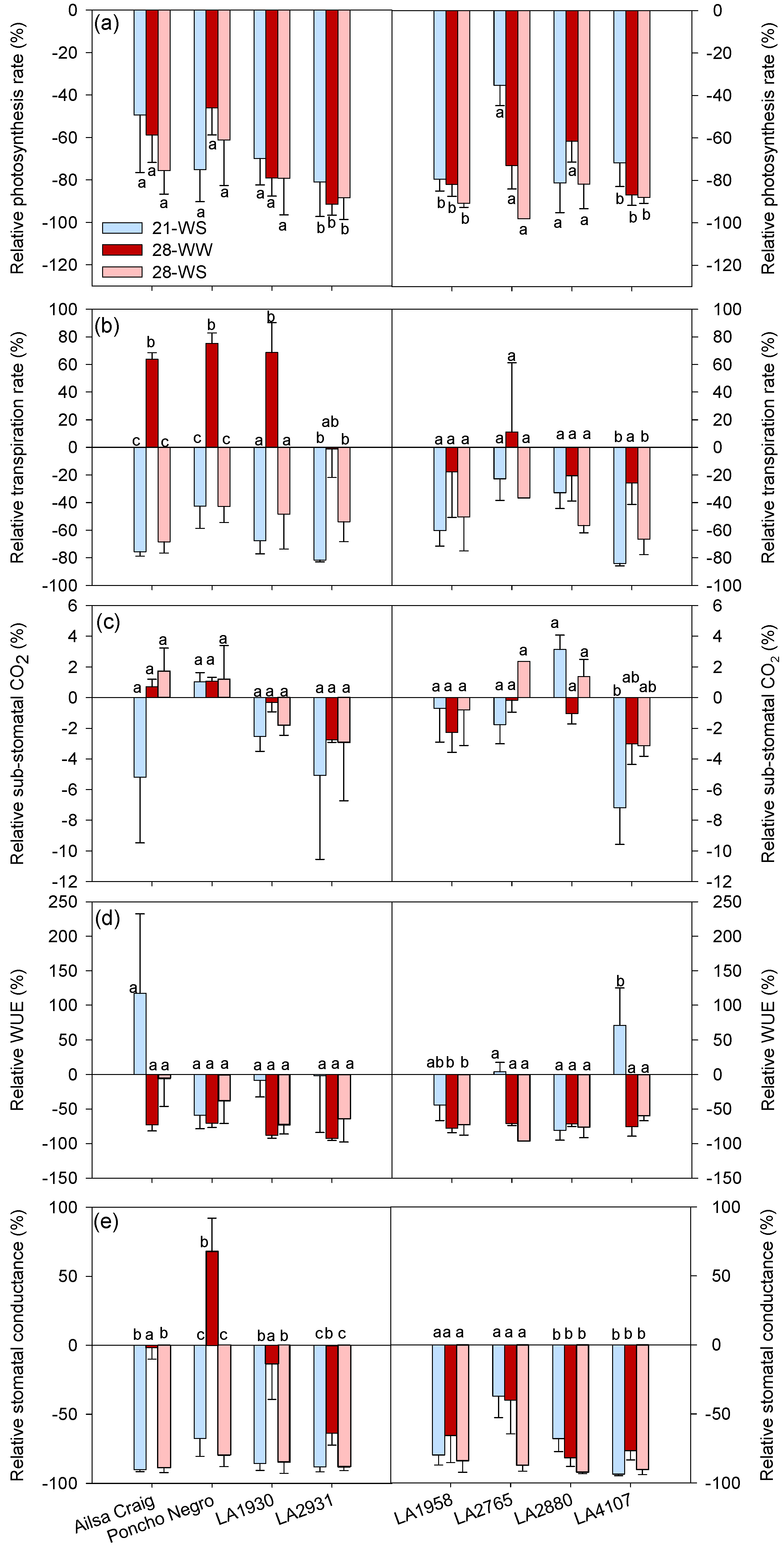
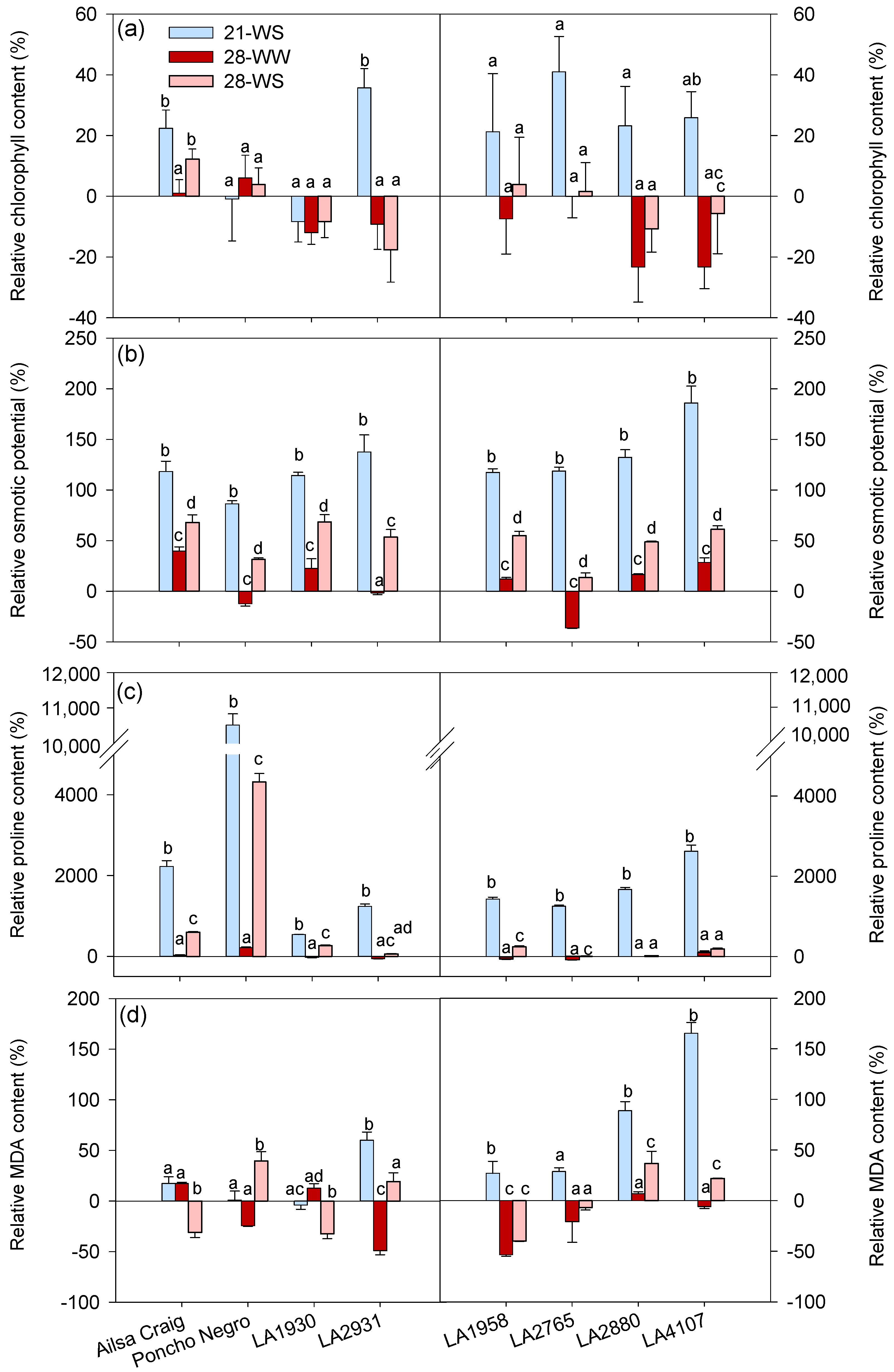
2. Results
2.1. Plant Growth
2.2. Plant Physiology
2.3. Expression of Water Stress and Heat Markers
3. Discussion
4. Materials and Methods
4.1. Plant Culture and Growth Conditions
4.2. Plant Growth and Water Status Parameters
4.3. Photosynthesis Parameters
4.4. Malondialdehyde (MDA) and Proline Extraction and Quantification
4.5. Quantitative Real-Time PCR and Data Evaluation
4.6. Statistical Treatment
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- FAOSTAT. Available online: http://www.fao.org/faostat/en/#data/QC (accessed on 4 March 2021).
- Amuji, C.F.; Beaumont, L.J.; Atwell, B.J. The effect of co-occurring heat and water stress on reproductive traits and yield of tomato (Solanum lycopersicum). Hortic. J. 2020, 89, 530–536. [Google Scholar] [CrossRef]
- IPCC. Climate Change 2014: Synthesis Report: Contribution of Working Groups I, II and III to the Fifth Assessment Report of the Intergovernmental Panel on Climate Change; Core Writing Team, Pachauri, R.K., Meyer, L.A., Eds.; IPCC: Geneva, Switzerland, 2014. [Google Scholar]
- Zhou, R.; Yu, X.; Wen, J.; Jensen, N.B.; dos Santos, T.M.; Wu, Z.; Rosenqvist, E.; Ottosen, C.-O. Interactive effects of elevated CO2 concentration and combined heat and drought stress on tomato photosynthesis. BMC Plant Biol. 2020, 20, 260. [Google Scholar] [CrossRef]
- Roy, S.J.; Tucker, E.J.; Tester, M. Genetic analysis of abiotic stress tolerance in crops. Curr. Opin. Plant Biol. 2011, 14, 232–239. [Google Scholar] [CrossRef]
- Prasch, C.M.; Sonnewald, U. Simultaneous application of heat, drought, and virus to Arabidopsis plants reveals significant shifts in signaling networks. Plant Physiol. 2013, 162, 1849–1866. [Google Scholar] [CrossRef]
- Zhou, R.; Kong, L.; Wu, Z.; Rosenqvist, E.; Wang, Y.; Zhao, L.; Zhao, T.; Ottosen, C.-O. Physiological response of tomatoes at drought, heat and their combination followed by recovery. Physiol. Plant. 2019, 165, 144–154. [Google Scholar] [CrossRef] [PubMed]
- Duan, H.; Wu, J.; Huang, G.; Zhou, S.; Liu, W.; Liao, Y.; Yang, X.; Xiao, Z.; Fan, H. Individual and interactive effects of drought and heat on leaf physiology of seedlings in an economically important crop. AoB Plants 2016, 9. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nankishore, A.; Farrell, A.D. The response of contrasting tomato genotypes to combined heat and drought stress. J. Plant Physiol. 2016, 202, 75–82. [Google Scholar] [CrossRef]
- Zhou, R.; Yu, X.; Ottosen, C.-O.; Rosenqvist, E.; Zhao, L.; Wang, Y.; Yu, W.; Zhao, T.; Wu, Z. Drought stress had a predominant effect over heat stress on three tomato cultivars subjected to combined stress. BMC Plant Biol. 2017, 17, 24. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fischer, I.; Camus-Kulandaivelu, L.; Allal, F.; Stephan, W. Adaptation to drought in two wild tomato species: The evolution of the Asr gene family. New Phytol. 2011, 190, 1032–1044. [Google Scholar] [CrossRef] [PubMed]
- Zsögön, A.; Čermák, T.; Naves, E.R.; Notini, M.M.; Edel, K.H.; Weinl, S.; Freschi, L.; Voytas, D.F.; Kudla, J.; Peres, L.E.P. De novo domestication of wild tomato using genome editing. Nat. Biotechnol. 2018, 36, 1211–1216. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Peralta, I.E.; Spooner, D.M.; Knapp, S. Taxonomy of wild tomatoes and their relatives (Solanum Sect. lycopersicoides, Sect. juglandifolia, Sect. lycopersicon; Solanaceae). Syst. Bot. Monogr. 2008, 84, 1–186. [Google Scholar]
- Aflitos, S.; Schijlen, E.; de Jong, H.; de Ridder, D.; Smit, S.; Finkers, R.; Wang, J.; Zhang, G.; Li, N.; Mao, L.; et al. Exploring genetic variation in the tomato (Solanum Section lycopersicon) clade by whole-genome sequencing. Plant J. 2014, 80, 136–148. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Alaguero-Cordovilla, A.; Gran-Gómez, F.J.; Tormos-Moltó, S.; Pérez-Pérez, J.M. Morphological characterization of root system architecture in diverse tomato genotypes during early growth. Int. J. Mol. Sci 2018, 19, 3888. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nakazato, T.; Warren, D.L.; Moyle, L.C. Ecological and geographic modes of species divergence in wild tomatoes. Am. J. Bot. 2010, 97, 680–693. [Google Scholar] [CrossRef]
- Böndel, K.B.; Nosenko, T.; Wolfgang, S. Signatures of natural selection in abiotic stress-responsive genes of Solanum chilense. R. Soc. Open Sci. 2018, 5, 171198. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bretó, M.P.; Asins, M.J.; Carbonell, E.A. Genetic variability in Lycopersicon species and their genetic relationships. Theor. Appl. Genet. 1993, 86, 113–120. [Google Scholar] [CrossRef] [PubMed]
- Stam, R.; Nosenko, T.; Hörger, A.C.; Stephan, W.; Seidel, M.; Kuhn, J.M.M.; Haberer, G.; Tellier, A. The de novo reference genome and transcriptome assemblies of the wild tomato species Solanum chilense highlights birth and death of NLR genes between tomato species. G3 Genes Genomes Genet. 2019, 9, 3933–3941. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Beddows, I.; Reddy, A.; Kloesges, T.; Rose, L.E. population genomics in wild tomatoes-the interplay of divergence and admixture. Genome Biol. Evol. 2017, 9, 3023–3038. [Google Scholar] [CrossRef] [Green Version]
- Raduski, A.R.; Igić, B. Biosystematic studies on the status of Solanum chilense. Am. J. Bot. 2021, 108, 520–537. [Google Scholar] [CrossRef]
- Chetelat, R.T.; Pertuzé, R.A.; Faúndez, L.; Graham, E.B.; Jones, C.M. Distribution, ecology and reproductive biology of wild tomatoes and related nightshades from the Atacama desert region of Northern Chile. Euphytica 2009, 167, 77–93. [Google Scholar] [CrossRef] [Green Version]
- Tapia, G.; Méndez, J.; Inostroza, L. Different combinations of morpho-physiological traits are responsible for tolerance to drought in wild tomatoes Solanum chilense and Solanum peruvianum. Plant Biol. 2016, 18, 406–416. [Google Scholar] [CrossRef]
- Böndel, K.B.; Lainer, H.; Nosenko, T.; Mboup, M.; Tellier, A.; Stephan, W. North-South colonization associated with local adaptation of the wild tomato species Solanum chilense. Mol. Biol. Evol. 2015, 32, 2932–2943. [Google Scholar] [CrossRef] [Green Version]
- Martínez, J.P.; Antúnez, A.; Araya, H.; Pertuzé, R.; Fuentes, L.; Lizana, X.C.; Lutts, S. Salt stress differently affects growth, water status and antioxidant enzyme activities in Solanum lycopersicum and its wild relative Solanum chilense. Aust. J. Bot. 2014, 62, 359–368. [Google Scholar] [CrossRef] [Green Version]
- Kashyap, S.P.; Kumari, N.; Mishra, P.; Prasad Moharana, D.; Aamir, M.; Singh, B.; Prasanna, H.C. Transcriptional regulation-mediating ROS homeostasis and physio-biochemical changes in wild tomato (Solanum chilense) and cultivated tomato (Solanum lycopersicum) under high salinity. Saudi J. Biol. Sci. 2020, 27, 1999–2009. [Google Scholar] [CrossRef]
- Gharbi, E.; Martínez, J.-P.; Benahmed, H.; Fauconnier, M.-L.; Lutts, S.; Quinet, M. Salicylic Acid Differently impacts ethylene and polyamine synthesis in the glycophyte Solanum lycopersicum and the wild-related halophyte Solanum chilense exposed to mild salt stress. Physiol. Plant. 2016, 158, 152–167. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gharbi, E.; Martínez, J.-P.; Benahmed, H.; Lepoint, G.; Vanpee, B.; Quinet, M.; Lutts, S. Inhibition of ethylene synthesis reduces salt-tolerance in tomato wild relative species Solanum chilense. J. Plant Physiol. 2017, 210, 24–37. [Google Scholar] [CrossRef] [PubMed]
- Gharbi, E.; Martínez, J.-P.; Benahmed, H.; Hichri, I.; Dobrev, P.I.; Motyka, V.; Quinet, M.; Lutts, S. Phytohormone profiling in relation to osmotic adjustment in NaCl-treated plants of the halophyte tomato wild relative species Solanum chilense comparatively to the cultivated glycophyte Solanum lycopersicum. Plant Sci. 2017, 258, 77–89. [Google Scholar] [CrossRef]
- Martínez, J.-P.; Antúnez, A.; Pertuzé, R.; Acosta, M.D.P.; Palma, X.; Fuentes, L.; Ayala, A.; Araya, H.; Lutts, S. Effects of saline water on water status, yield and fruit quality of wild (Solanum chilense) and domesticated (Solanum lycopersicum Var. cerasiforme) tomatoes. Exp. Agric. 2012, 48, 573–586. [Google Scholar] [CrossRef]
- Martínez, J.P.; Fuentes, R.; Farías, K.; Lizana, C.; Alfaro, J.F.; Fuentes, L.; Calabrese, N.; Bigot, S.; Quinet, M.; Lutts, S. Effects of salt stress on fruit antioxidant capacity of wild (Solanum chilense) and domesticated (Solanum lycopersicum Var. cerasiforme) tomatoes. Agronomy 2020, 10, 1481. [Google Scholar] [CrossRef]
- Kashyap, S.P.; Prasanna, H.C.; Kumari, N.; Mishra, P.; Singh, B. Understanding salt tolerance mechanism using transcriptome profiling and de novo assembly of wild tomato Solanum chilense. Sci. Rep. 2020, 10, 15835. [Google Scholar] [CrossRef] [PubMed]
- Godoy, F.; Olivos-Hernández, K.; Stange, C.; Handford, M. Abiotic stress in crop species: Improving tolerance by applying plant metabolites. Plants 2021, 10, 186. [Google Scholar] [CrossRef] [PubMed]
- Smith, D.T.; Potgieter, A.B.; Chapman, S.C. Scaling up high-throughput phenotyping for abiotic stress selection in the field. Theor. Appl. Genet. 2021, 134, 1845–1866. [Google Scholar] [CrossRef] [PubMed]
- Krishna, R.; Karkute, S.G.; Ansari, W.A.; Jaiswal, D.K.; Verma, J.P.; Singh, M. Transgenic tomatoes for abiotic stress tolerance: Status and way ahead. 3 Biotech 2019, 9, 143. [Google Scholar] [CrossRef] [PubMed]
- Snowdon, R.J.; Wittkop, B.; Chen, T.-W.; Stahl, A. Crop adaptation to climate change as a consequence of long-term breeding. Theor. Appl. Genet. 2021, 134, 1613–1623. [Google Scholar] [CrossRef]
- Bodner, G.; Nakhforoosh, A.; Kaul, H.-P. Management of crop water under drought: A review. Agron. Sustain. Dev. 2015, 35, 401–442. [Google Scholar] [CrossRef]
- Ullah, U.; Ashraf, M.; Shahzad, S.M.; Siddiqui, A.R.; Piracha, M.A.; Suleman, M. Growth behavior of tomato (Solanum lycopersicum L.) Under drought stress in the presence of silicon and plant growth promoting rhizobacteria. Soil Environ. 2016, 35, 65–75. [Google Scholar]
- Ayenan, M.A.T.; Danquah, A.; Hanson, P.; Ampomah-Dwamena, C.; Sodedji, F.A.K.; Asante, I.K.; Danquah, E.Y. accelerating breeding for heat tolerance in tomato (Solanum lycopersicum L.): An integrated approach. Agronomy 2019, 9, 720. [Google Scholar] [CrossRef] [Green Version]
- Arena, C.; Conti, S.; Francesca, S.; Melchionna, G.; Hájek, J.; Barták, M.; Barone, A.; Rigano, M.M. Eco-physiological screening of different tomato genotypes in response to high temperatures: A combined field-to-laboratory approach. Plants 2020, 9, 508. [Google Scholar] [CrossRef] [Green Version]
- Zhou, R.; Kjær, K.H.; Rosenqvist, E.; Yu, X.; Wu, Z.; Ottosen, C.-O. Physiological response to heat stress during seedling and anthesis stage in tomato genotypes differing in heat tolerance. J. Agron. Crop. Sci. 2017, 203, 68–80. [Google Scholar] [CrossRef]
- Zhou, R.; Kong, L.; Yu, X.; Ottosen, C.-O.; Zhao, T.; Jiang, F.; Wu, Z. Oxidative damage and antioxidant mechanism in tomatoes responding to drought and heat stress. Acta Physiol. Plant. 2019, 41, 20. [Google Scholar] [CrossRef]
- Gong, P.; Zhang, J.; Li, H.; Yang, C.; Zhang, C.; Zhang, X.; Khurram, Z.; Zhang, Y.; Wang, T.; Fei, Z.; et al. Transcriptional profiles of drought-responsive genes in modulating transcription signal transduction, and biochemical pathways in Tomato. J. Exp. Bot. 2010, 61, 3563–3575. [Google Scholar] [CrossRef] [PubMed]
- Klay, I.; Gouia, S.; Liu, M.; Mila, I.; Khoudi, H.; Bernadac, A.; Bouzayen, M.; Pirrello, J. Ethylene Response factors (ERF) are differentially regulated by different abiotic stress types in tomato plants. Plant Sci. 2018, 274, 137–145. [Google Scholar] [CrossRef] [PubMed]
- Xia, H.; Camus-Kulandaivelu, L.; Stephan, W.; Tellier, A.; Zhang, Z. Nucleotide Diversity Patterns of local adaptation at drought-related candidate genes in wild tomatoes. Mol. Ecol. 2010, 19, 4144–4154. [Google Scholar] [CrossRef] [PubMed]
- Sato, S.; Peet, M.M.; Thomas, J.F. Physiological factors limit fruit set of tomato (Lycopersicon esculentum Mill.) under chronic, mild heat stress. Plant Cell Environ. 2000, 23, 719–726. [Google Scholar] [CrossRef]
- Nosenko, T.; Böndel, K.B.; Kumpfmüller, G.; Stephan, W. Adaptation to low temperatures in the wild tomato species Solanum chilense. Mol. Ecol. 2016, 25, 2853–2869. [Google Scholar] [CrossRef]
- Zhou, J.; Reddy, S.; Zhou, S.; Sauvé, R.J.; Bhatti, S.; Fish, T.; Thannhauser, T.W. Effect of heat stress on leaf proteome and enzyme activity in Solanum chilense. Plant Stress 2012, 6, 8–13. [Google Scholar]
- Rezaei, E.E.; Webber, H.; Gaiser, T.; Naab, J.; Ewert, F. Heat stress in cereals: Mechanisms and modelling. Eur. J. Agron. 2015, 64, 98–113. [Google Scholar] [CrossRef]
- Peguero-Pina, J.J.; Vilagrosa, A.; Alonso-Forn, D.; Ferrio, J.P.; Sancho-Knapik, D.; Gil-Pelegrín, E. Living in drylands: Functional adaptations of trees and shrubs to cope with high temperatures and water scarcity. Forests 2020, 11, 1028. [Google Scholar] [CrossRef]
- Singh, D.; Laxmi, A. Transcriptional regulation of drought response: A tortuous network of transcriptional factors. Front. Plant Sci. 2015, 6, 895. [Google Scholar] [CrossRef] [Green Version]
- Yáñez, M.; Cáceres, S.; Orellana, S.; Bastías, A.; Verdugo, I.; Ruiz-Lara, S.; Casaretto, J.A. An Abiotic stress-responsive BZIP transcription factor from wild and cultivated tomatoes regulates stress-related genes. Plant Cell Rep. 2009, 28, 1497–1507. [Google Scholar] [CrossRef]
- Gerszberg, A.; Hnatuszko-Konka, K. Tomato Tolerance to abiotic stress: A review of most often engineered target sequences. Plant Growth Regul. 2017, 83, 175–198. [Google Scholar] [CrossRef] [Green Version]
- Karkute, S.G.; Ansari, W.A.; Singh, A.K.; Singh, P.M.; Rai, N.; Bahadur, A.; Singh, J. Characterization of high-temperature stress-tolerant tomato (Solanum lycopersicum L.) genotypes by biochemical analysis and expression profiling of heat-responsive genes. 3 Biotech 2021, 11, 45. [Google Scholar] [CrossRef] [PubMed]
- Lutts, S.; Majerus, V.; Kinet, J.-M. NaCl effects on proline metabolism in rice (Oryza sativa) seedlings. Physiol. Plant. 1999, 105, 450–458. [Google Scholar] [CrossRef]
- Maxwell, K.; Johnson, G.N. Chlorophyll Fluorescence--a Practical Guide. J. Exp. Bot. 2000, 51, 659–668. [Google Scholar] [CrossRef] [PubMed]
- Heath, R.L.; Packer, L. Photoperoxidation in isolated chloroplasts. I. Kinetics and stoichiometry of fatty acid peroxidation. Arch. Biochem. Biophys. 1968, 125, 189–198. [Google Scholar] [CrossRef]
- Bates, L.S.; Waldren, R.P.; Teare, I.D. Rapid determination of free proline for water-stress studies. Plant Soil 1973, 39, 205–207. [Google Scholar] [CrossRef]
- Untergasser, A.; Nijveen, H.; Rao, X.; Bisseling, T.; Geurts, R.; Leunissen, J.A.M. Primer3Plus, an enhanced web interface to Primer3. Nucleic Acids Res. 2007, 35, W71–W74. [Google Scholar] [CrossRef] [Green Version]
- R: The R Project for Statistical Computing. Available online: https://www.r-project.org/ (accessed on 23 September 2020).








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Accession | 21-WW | 21-WS | 28-WW | 28-WS |
|---|---|---|---|---|
| Number of ramifications | ||||
| Ailsa Craig | 4.5 ± 0.3 ab | 3.0 ± 0.5 b | 5.2 ± 0.6 a | 3.3 ± 0.3 b |
| Poncho Negro | 3.2 ± 0.5 ab | 0.8 ± 0.2 c | 4.0 ± 0.5 a | 2.5 ± 0.2 b |
| LA1930 | 4.8 ± 2.8 ab | 0.8 ± 0.6 b | 11.2 ± 3.0 a | 5.3 ± 3.0 ab |
| LA2931 | 8.2 ± 1.1 a | 9.2 ± 0.7 b | 15.5 ± 3.0 b | 11.2 ± 2.4 b |
| LA1958 | 8.3 ± 2.9 a | 5.0 ± 1.6 a | 7.7 ± 1.2 a | 9.5 ± 1.8 a |
| LA2765 | 8.0 ± 2.9 a | 9.8 ± 1.8 a | 7.3 ± 2.0 a | 9.0 ± 1.2 a |
| LA2880 | 5.7 ± 1.8 b | 4.4 ± 1.0 b | 11.5 ± 2.0 a | 6.3 ± 0.8 b |
| LA4107 | 9.7 ± 2.7 a | 10.8 ± 1.2 a | 10.8 ± 1.6 a | 8.6 ± 1.3 a |
| Number of inflorescences on the main stem | ||||
| Ailsa Craig | 5.50 ± 0.34 a | 3.00 ± 0.45 b | 4.83 ± 0.17 a | 3.33 ± 0.33 b |
| Poncho Negro | 2.50 ± 0.67 a | 1.33 ± 0.49 a | 2.67 ± 0.33 a | 1.67 ± 0.42 a |
| LA1930 | 0.80 ± 0.37 a | 0.00 ± 0.00 b | 0.00 ± 0.00 b | 0.17 ± 0.17 b |
| LA2931 | 1.40 ± 0.75 a | 3.00 ± 0.58 a | 2.67 ± 0.88 a | 2.83 ± 1.01 a |
| LA1958 | 1.80 ± 0.92 a | 1.00 ± 0.55 a | 0.17 ± 0.17 a | 1.83 ± 0.60 a |
| LA2765 | 2.50 ± 0.50 a | 2.33 ± 0.67 a | 1.50 ± 0.96 a | 1.37 ± 0.56 a |
| LA2880 | 1.75 ± 0.75 a | 2.60 ± 0.75 a | 0.20 ± 0.20 a | 0.83 ± 0.65 a |
| LA4107 | 2.67 ± 0.95 ab | 3.33 ± 0.49 a | 1.00 ± 0.45 b | 0.50 ± 0.50 b |
| Accession | 21-WW | 21-WS | 28-WW | 28-WS |
|---|---|---|---|---|
| Efficiency of photosystem 2 (φPS2) | ||||
| Ailsa Craig | 0.78 ± 0.02 a | 0.82 ± 0 a | 0.81 ± 0.01 a | 0.80 ± 0.01 a |
| Poncho Negro | 0.61 ± 0.14 a | 0.59 ± 0.11 a | 0.76 ± 0.02 a | 0.79 ± 0.01 a |
| LA1930 | 0.81 ± 0.01 a | 0.76 ± 0.05 a | 0.81 ± 0.01 a | 0.75 ± 0.05 a |
| LA2931 | 0.73 ± 0.04 a | 0.83 ± 0.01 a | 0.74 ± 0.04 a | 0.77 ± 0.02 a |
| LA1958 | 0.75 ± 0.04 a | 0.78 ± 0.05 a | 0.77 ± 0.04 a | 0.70 ± 0.08 a |
| LA2765 | 0.71 ± 0.09 a | 0.74 ± 0.08 a | 0.72 ± 0.07 a | 0.77 ± 0.02 a |
| LA2880 | 0.57 ± 0.10 a | 0.52 ± 0.13 a | 0.73 ± 0.02 a | 0.79 ± 0.02 a |
| LA4107 | 0.74 ± 0.06 a | 0.84 ± 0.01 a | 0.74 ± 0.04 a | 0.73 ± 0.03 a |
| Non-photochemical quenching (NPQ) | ||||
| Ailsa Craig | 0.32 ± 0.09 a | 0.23 ± 0.02 a | 0.23 ± 0.01 a | 0.19 ± 0.06 a |
| Poncho Negro | 0.30 ± 0.03 ab | 0.47 ± 0.08 a | 0.28 ± 0.05 ab | 0.20 ± 0.02 b |
| LA1930 | 0.25 ± 0.05 a | 0.23 ± 0.02 a | 0.26 ± 0.03 a | 0.30 ± 0.06 a |
| LA2931 | 0.27 ± 0.05 a | 0.25 ± 0.01 a | 0.43 ± 0.17 a | 0.16 ± 0.02 a |
| LA1958 | 0.27 ± 0.04 a | 0.30 ± 0.03 a | 0.32 ± 0.06 a | 0.20 ± 0.03 a |
| LA2765 | 0.24 ± 0.04 a | 0.37 ± 0.07 a | 0.18 ± 0.04 b | 0.16 ± 0.04 b |
| LA2880 | 0.34 ± 0.02 a | 0.32 ± 0.10 a | 0.32 ± 0.10 a | 0.20 ± 0.03 a |
| LA4107 | 0.27 ± 0.05 a | 0.20 ± 0.03 a | 0.24 ± 0.05 a | 0.17 ± 0.06 a |
| Accession | 21-WW | 21-WS | 28-WW | 28-WS |
|---|---|---|---|---|
| Leaf water content (%) | ||||
| Ailsa Craig | 90.30 ± 0.71 a | 83.56 ± 2.54 a | 89.22 ± 0.28 a | 83.86 ± 1.39 a |
| Poncho Negro | 89.88 ± 0.98 a | 83.13 ± 2.92 a | 89.82 ± 1.31 a | 81.15 ± 2.43 a |
| LA1930 | 88.88 ± 0.55 a | 82.61 ± 0.94 b | 91.35 ± 0.40 a | 91.79 ± 1.37 a |
| LA2931 | 89.06 ± 0.84 a | 77.54 ± 2.52 b | 89.61 ± 0.21 a | 90.82 ± 1.77 a |
| LA1958 | 90.70 ± 0.95 a | 88.31 ± 3.94 a | 90.50 ± 0.47 a | 91.46 ± 0.99 a |
| LA2765 | 90.02 ± 1.50 a | 83.87 ± 1.01 a | 90.11 ± 0.78 a | 86.46 ± 2.34 a |
| LA2880 | 87.12 ± 1.61 a | 85.47 ± 4.08 a | 93.18 ± 2.95 a | 91.97 ± 1.52 a |
| LA4107 | 93.11 ± 0.87 a | 82.16 ± 0.35 b | 91.10 ± 0.20 a | 92.37 ± 2.22 a |
| Stem water content (%) | ||||
| Ailsa Craig | 92.06 ± 0.32 a | 86.97 ± 0.10 b | 89.03 ± 0.30 a | 87.23 ± 1.32 b |
| Poncho Negro | 91.94 ± 0.61 a | 90.12 ± 0.73 a | 86.99 ± 0.52 b | 85.45 ± 1.56 b |
| LA1930 | 85.80 ± 1.12 a | 86.37 ± 1.12 a | 89.54 ± 0.64 a | 89.97 ± 2.02 a |
| LA2931 | 86.60 ± 0.80 a | 67.01 ± 15.36 a | 78.60 ± 12.35 a | 90.47 ± 2.32 a |
| LA1958 | 90.35 ± 1.34 a | 89.83 ± 3.23 a | 90.58 ± 0.78 a | 88.76 ± 0.19 a |
| LA2765 | 91.58 ± 2.09 a | 2.81 ± 1.62 a | 90.44 ± 1.61 a | 87.21 ± 1.73 a |
| LA2880 | 92.48 ± 2.35 a | 86.33 ± 4.65 a | 92.76 ± 3.46 a | 91.90 ± 2.22 a |
| LA4107 | 93.42 ± 1.112 a | 83.54 ± 0.35 b | 94.02 ± 2.06 a | 91.96 ± 1.92 a |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Blanchard-Gros, R.; Bigot, S.; Martinez, J.-P.; Lutts, S.; Guerriero, G.; Quinet, M. Comparison of Drought and Heat Resistance Strategies among Six Populations of Solanum chilense and Two Cultivars of Solanum lycopersicum. Plants 2021, 10, 1720. https://0-doi-org.brum.beds.ac.uk/10.3390/plants10081720
Blanchard-Gros R, Bigot S, Martinez J-P, Lutts S, Guerriero G, Quinet M. Comparison of Drought and Heat Resistance Strategies among Six Populations of Solanum chilense and Two Cultivars of Solanum lycopersicum. Plants. 2021; 10(8):1720. https://0-doi-org.brum.beds.ac.uk/10.3390/plants10081720
Chicago/Turabian StyleBlanchard-Gros, Rémi, Servane Bigot, Juan-Pablo Martinez, Stanley Lutts, Gea Guerriero, and Muriel Quinet. 2021. "Comparison of Drought and Heat Resistance Strategies among Six Populations of Solanum chilense and Two Cultivars of Solanum lycopersicum" Plants 10, no. 8: 1720. https://0-doi-org.brum.beds.ac.uk/10.3390/plants10081720