
Genome-Wide Analysis of the IQM Gene Family in Rice (Oryza sativa L.)

Guangzhou Key Laboratory for Functional Study on Plant Stress-Resistant Genes, School of Life Sciences, Guangzhou University, Guangzhou 510006, China
*
Author to whom correspondence should be addressed.
Plants 2021, 10(9), 1949; https://0-doi-org.brum.beds.ac.uk/10.3390/plants10091949
Submission received: 4 August 2021
/
Revised: 6 September 2021
/
Accepted: 14 September 2021
/
Published: 18 September 2021
Abstract
:Members of the IQM (IQ-Motif Containing) gene family are involved in plant growth and developmental processes, biotic and abiotic stress response. To systematically analyze the IQM gene family and their expression profiles under diverse biotic and abiotic stresses, we identified 8 IQM genes in the rice genome. In the current study, the whole genome identification and characterization of OsIQMs, including the gene and protein structure, genome localization, phylogenetic relationship, gene expression and yeast two-hybrid were performed. Eight IQM genes were classified into three subfamilies (I–III) according to the phylogenetic analysis. Gene structure and protein motif analyses showed that these IQM genes are relatively conserved within each subfamily of rice. The 8 OsIQM genes are distributed on seven out of the twelve chromosomes, with three IQM gene pairs involved in segmental duplication events. The evolutionary patterns analysis revealed that the IQM genes underwent a large-scale event within the last 20 to 9 million years. In addition, quantitative real-time PCR analysis of eight OsIQMs genes displayed different expression patterns at different developmental stages and in different tissues as well as showed that most IQM genes were responsive to PEG, NaCl, jasmonic acid (JA), abscisic acid (ABA) treatment, suggesting their crucial roles in biotic, and abiotic stress response. Additionally, a yeast two-hybrid assay showed that OsIQMs can interact with OsCaMs, and the IQ motif of OsIQMs is required for OsIQMs to combine with OsCaMs. Our results will be valuable to further characterize the important biological functions of rice IQM genes.
1. Introduction
As one of the most important intracellular second messenger, Ca2+ plays a prominent role in many essential biological processes of plants, such as numerous developmental processes and responses to various kinds of biotic and abiotic stresses [1]. The spatial and transient changes of cytoplasmic Ca2+ levels in response to various stresses were detected and decoded by specific Ca2+ sensors, which then transduce these changes of Ca2+ levels into a series of downstream effects [2,3].
To date, most of the Ca2+ sensors have been identified to contain conserved Ca2+-binding helix-loop-helix EF-hand motif [4]. Based on the number of EF hand motifs, four types of Ca2+ sensors were identified in higher plants: calmodulins (CaM), CaM-like proteins (CML), calcineurin B-like proteins (CBL) and calcium-dependent protein kinases (CDPK) [5,6]. Among these Ca2+ sensors, CaM is a highly conserved small, acidic protein and has been getting the most attention in Ca2+ signal transduction [7]. CaM has no enzymatic activity of its own; however, CaM produces a change in conformation after binding Ca2+, and then it can activate a wide range of target proteins named calmodulin-binding proteins (CaMBPs) involved in diverse cellular processes [7,8].
The CaMBPs include transcription factors, metabolic enzymes, cytoskeleton, chaperones and kinases to involve in plant development, metabolic regulation, stress response, defense reactions [1,9]. The recognition motifs for the interaction between CaM and CaMBPs contain three conserved types: one is an IQ motif [ILV] QxxxRxxxx [R, K] or (IQxxxRGxxxR), which binds CaM in a Ca2+-independent manner, others are the 1-5-10 motif ([FILVW] × 3[FILV] × [FILVW]) and 1-8-14 motif ([FILVW] × 6[FAILVW] × 5[FILVW]), which are thought to mediate CaM retention in a Ca2+-dependent manner [10]. In plants, five classes of the IQ motif-containing protein family, namely, the IQ67-domain containing protein (IQD) family, the myosin protein family, the calmodulin-binding transcription activator (CAMTA) family, the cyclic nucleotide-gated channels (CNGC) family and the IQ motif containing protein (IQM) family, have been identified [11]. The members of these five families had divergence in the number and distribution of IQ motifs. Compared to other families, the functional studies of the IQM family are needed to explore more. To date, the plant-specific IQM gene family have been comprehensively analyzed only in Arabidopsis genomes. In Arabidopsis, six members named IQM1 to IQM6 were identified [11]. Across all IQM gene family members, only one IQ motif, located in the N-terminal of these proteins, is the common characteristic. Moreover, the IQ motif overlaps with the sequence, which is similar to the pea heavy-metal induced protein 6A (PHMIP 6A) [12]. Meanwhile, there is a segment of trichosanthin that consists of 248 amino-acid residues in the C-terminal [11]. Subsequently, a few functional investigations of IQM proteins have been reported in recent years: in Arabidopsis thaliana, AtIQM1 proteins can interact with AtCaM5 in a Ca2+-dependent manner and modulates stomatal movement by affecting the content of ROS [13]. Furthermore, it was reported that AtIQM1 is a key regulatory factor in plant defense against the necrotrophic pathogen Botrytis cinerea by regulating JA biosynthesis [14]. In addition, AtIQM4 is involved in seed dormancy and germination [15], and the mutation of AtIQM5 delays flowering possibly through modulating the juvenile-to-adult transition [16]. Although, some previous studies had revealed the roles of some IQMs and the underlying mechanisms in Arabidopsis. On the other hand, the understanding and systematic analysis of the IQM family members in the other crops need to be explored. We considered that the IQM gene family might have similar functions in rice as in Arabidopsis. Therefore, to obtain a better understanding of the IQM gene family and to improve future study of this family with regard to resistance to biotic and abiotic stress in rice, the current study performed a comprehensive genome-wide analysis of them. In this study, eight non-redundant IQM encoding genes were identified by genome-wide analysis technology. The phylogenetic relationships, gene structure, conserved motifs, evolutionary patterns and divergence, yeast two-hybrid assay and expression profiling in response to abiotic stress and hormone were explored, providing a theoretical foundation for the downstream functional analysis of the rice IQM gene.
2. Results
2.1. Identification of IQM Gene Family in Rice Genome
To identify the IQM gene family in rice, the previously reported Arabidopsis IQM proteins and their IQ domain were used as the query sequences to search the RAP-DB (http://rapdb.dna.affrc.go.jp/ accessed on 19 July 2020). Here, a total of eight putative OsIQM genes contained the conserved IQ calmodulin-binding motif were identified through SMART. The present study coined these eight OsIQM genes as OsIQM1 to OsIQM8, according to their physical locations (from top to bottom) on chromosomes 1–12 (Table 1). The characteristics of the OsIQM genes, including ORFs, amino acids (aa), isoelectric point (pI), molecular weight (MW), chromosome location and start position, were listed in Table 1. The identified OsIQM gene family members encode proteins ranging from 475 to 650 aa in length and the predicted molecular masses ranged from 52.2 to 73.1 kDa. Additionally, the predicted pI values ranged from 6.19 to 9.28.
2.2. Phylogenetic Analysis of the OsIQM Gene Family
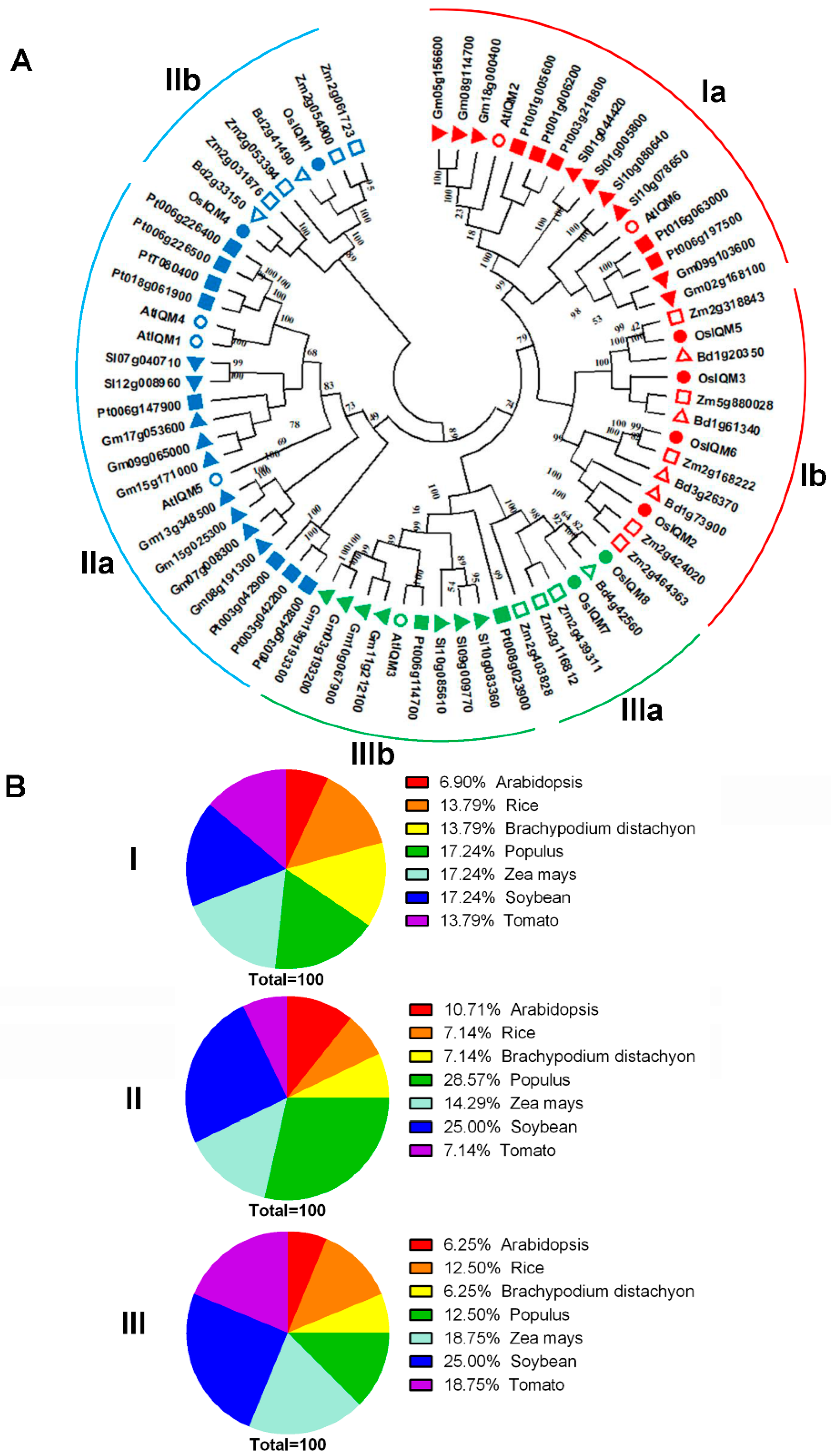
To better understand the molecular evolution and phylogenetic relationship, the current study employed MEGA X software to establish the unrooted phylogenetic tree of IQM members from Brachypodium distachyon, Oryza sativa, Arabidopsis thaliana, Populus trichocarpa, Zea mays, Glycine max and Solanum lycopersicum [17]. A phylogenetic tree from the alignment of full-length IQM protein sequences was constructed using the neighbor-joining method. The NJ phylogenetic tree contained 73 IQM protein sequences from Oryza sativa (8), Brachypodium distachyon (7), Arabidopsis thaliana (6), Populus trichocarpa (15), Zea mays (12), Glycine max (16) and Solanum lycopersicum (9). The results showed that these IQMs could be separated into three different subfamilies (I, II, and III) (Figure 1A). The IQM I and II subfamilies covered mostly IQM genes. By contrast, the IQM III subfamily had the fewest IQM genes (Figure 1A). Generally, subfamily I and II constituted the most clade, containing 29 and 28 IQMs, which accounted for 50% and 48% of the total IQM genes, respectively. We further examined each of the subfamilies and found they all separated into two distinct groups (a and b) of monocot and eudicot (Figure 1A). This result suggested the IQM proteins in monocot and eudicot may produce different functions. Additionally, as shown in Figure 1B, PtIQMs, ZmIQMs and GmIQMs account for 17.24% in subfamily I; PtIQMs account for 28.57% in subfamily II; GmIQMs account for 25% in subfamily III. It revealed that the plant IQM protein sequence distribution predominates with species bias and there were more IQM genes in each subfamily for dicotyledonous than for monocotyledonous.
In the phylogenetic tree, the putative orthologous genes of OsIQMs were found in Zea mays and Brachypodium distachyon, which supported by the three ortholog pairs (OsIQM1/Bd2g41490, OsIQM4/Bd2g33150, OsIQM5/Zm2g318843, OsIQM6/Zm2g168222 and OsIQM8/Bd4g42560) (Figure 1A). There were three monocot ortholog pairs that were found in Oryza sativa/Brachypodium distachyon. Only one eudicot ortholog pairs were observed in Arabidopsis thaliana and Populus trichocarpa (Figure 1A). Generally, the monocot IQM genes share branches with the nearest monocot orthologs, while eudicot IQM genes form sister pairs with the nearest eudicot orthologs. Indeed, the ortholog pair developed by eudicot and monocot was not found in the phylogenetic tree (Figure 1A), indicating the member of each subfamily from eudicot and monocot may not come from a common ancestor.
2.3. Gene Structure and Conserved Motifs of OsIQM Genes
In order to understand the structural diversity of rice IQM genes, we first constructed an unrooted phylogenetic tree based on the alignment of the full-length sequences of the eight OsIQM proteins, and sequences of rice IQM proteins were retrieved from the RAP-DB database (http://rapdb.dna.affrc.go.jp/ accessed on 19 July 2020). The rice IQM family members can be classified into three subfamilies: subfamily I, II and III, with each subfamily containing four, two, and two members, respectively (Figure 2). These results were in good agreement with that described above for the 7 plant species (Figure 1A and Figure 2). These eight OsIQM genes formed four sister pairs, and all of them displayed high bootstrap support. The exon–intron organization map analysis indicated that the eight OsIQM genes contain different numbers of exons, ranging from six to nine (Figure 2). Furthermore, the genes in one subfamily shared similar gene structures in terms of either the intron/exon number or length. However, one sister gene pairs showed greater changes in their structural organizations (OsIQM1/4) and varying numbers of exons and introns.
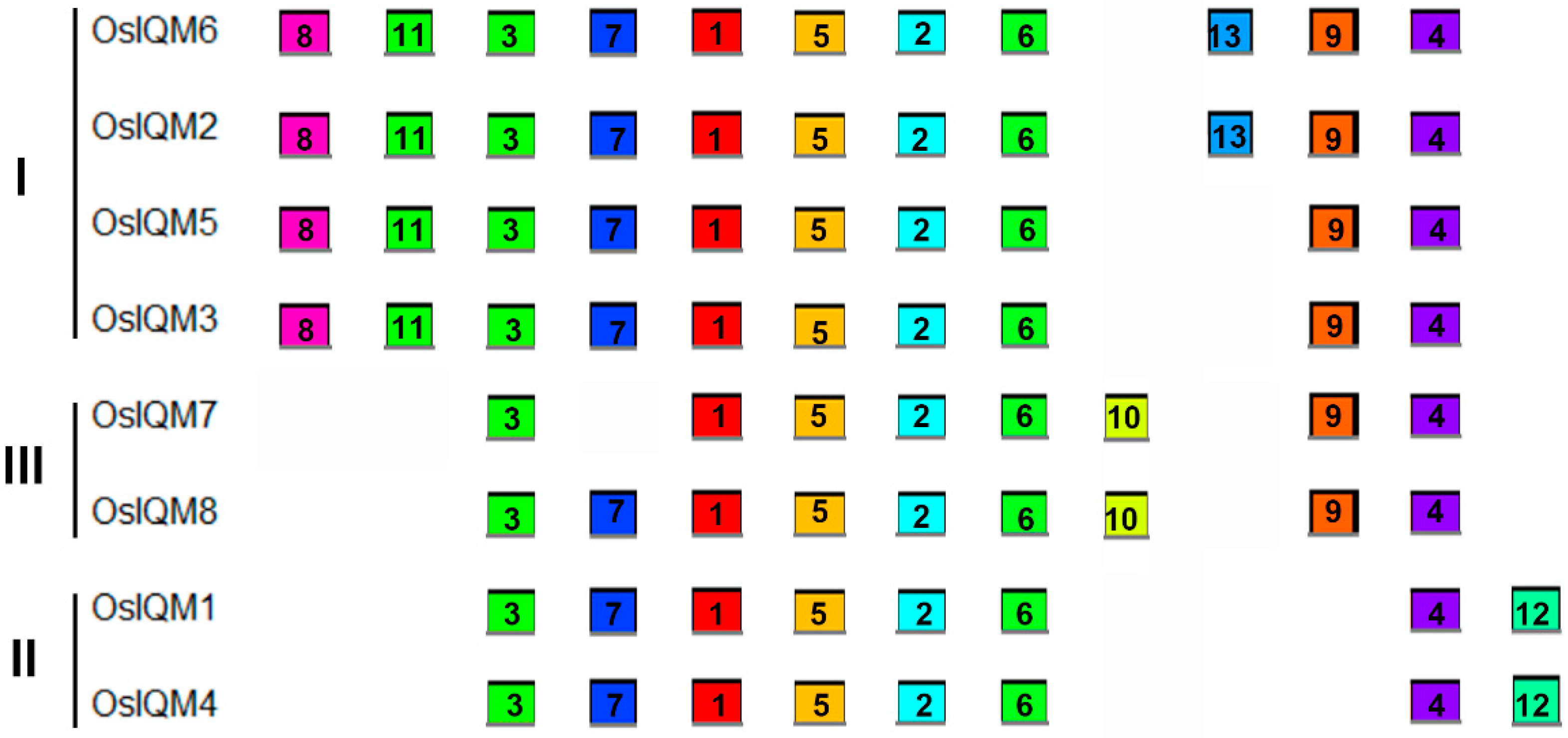
MEME was used to analyze protein motifs in the eight IQM protein sequences in rice (Figure 3). In total, 13 conserved motifs were identified and named motif 1–13. Figure S2 presents the conserved amino acid sequences and the length of each motif. Each of the putative motifs identified by MEME was subsequently verified using the Pfam and SMART. Among them, motif three was found to encode the IQ domain and related to heavy metal-induced protein 6 (HMIP6) of pea [11], motif five was considered to have similarity with trichosanthin [11], while motif six is a representative JmjN motif. However, the biological annotation of the other putative motifs remains unclear and need to be further investigated. IQM proteins in the same subfamily often have a common motif, suggesting functional similarities among these IQM proteins. In addition, some motifs were also detected in the specific subfamily. For example, motifs 12 were only observed in the subfamily II and motifs 10 are only present in subfamily III (Figure 3). These subfamily-specific motifs may produce functional divergence of IQM genes in rice.
2.4. Chromosomal Locations and Gene Duplication
The distribution of OsIQM genes was constructed by their positions in the rice chromosomes. The results show that eight OsIQM genes are unevenly distributed across 7 of the 12 chromosomes (Figure 4A). Both OsIQM2 and OsIQM3 were localized to chromosome 3. OsIQM1, OsIQM4, OsIQM5, OsIQM6, OsIQM7 and OsIQM8 were localized on chromosomes 1, 5, 7, 10, 11 and 12, respectively (Figure 4A). The nearest SSR markers of eight OsIQM genes were mapped to 7 rice chromosomes. Segmental duplication, tandem duplication and retroposition have been suggested to drive the expansion of gene families in genomes [18]. To evaluate the evolutionary patterns of the OsIQM gene family, segmental duplication and tandem duplication analysis were performed. According to the rice segmental duplication database from RGAP, we found three pairs (OsIQM2/OsIQM6, OsIQM3/OsIQM5 and OsIQM7/OsIQM8) that were located in duplicated chromosomal regions, suggesting that six genes came from segmental duplication. In addition, the chromosome distribution analysis indicated that tandem duplications have not been involved in the expansion of the OsIQM gene. These results indicated that the expansion of the OsIQM gene family results from segmental duplication events.
To explore the evolutionary selection of the three pairs of OsIQM genes, the Ka (non-synonymous substitution rate), the Ks (synonymous substitution rate) and the Ka/Ks ratios were calculated for each duplicated OsIQM gene pair in the family. As we know, Ka/Ks ratio < 1 indicates purifying selection which eliminates deleterious mutations and maintains the protein, where the Ka/Ks ratio > 1 indicates positive selection that accelerated evolution by changing the protein [19]. Our study showed that the Ka/Ks ratio for the one segmentally duplicated gene pair (OsIQM7/OsIQM8) was <1, suggesting that the gene pair had mainly had negative or purifying selection after the duplication events. The Ka/Ks ratios of OsIQM2/OsIQM6 and OsIQM3/OsIQM5 were more than 1, suggesting that the two duplicated pairs had evolved under positive selection (Figure 4B). According to the neutral evolutionary rate (λ) of 6.5 × 10−9 substitutions per synonymous site per year for the rice genome, we calculated the dates of the duplication events [20]. The divergence time of duplicated OsIQM gene pairs ranged from 11.9 to 19.8 Mya (Figure 4C). The duplication time within the last 20 to 9 million years for the three pairs occurred after Zizaniinae and Oryzinae were divided and before the Oryza genus branched off from the remaining genera of Oryzeae [21].
2.5. The Subcellular Localization of OsIQMs
In order to gain further insights into these OsIQM proteins, we predict their possible subcellular localization patterns by WoLF PSORT. The WoLF PSORT prediction analysis indicated that OsIQM2–8 are localized in the cell nucleus and OsIQM1 are localized in the chloroplasts (Table 1). To further determine the subcellular localization of OsIQMs, full-length cDNA was fused with Green Fluorescent Protein (GFP) driven by the CMV 35S promoter and transiently expressed in protoplasts of Arabidopsis cells. The results showed the OsIQM1 was localized in both the cytosol and nucleus, while the OsIQM2–8 were mainly localized in the cytosol. (Figure 5). These indicated the localization of OsIQMs is not consistent with the predicted localization using WoLF PSORT programs.
2.6. Protein Interaction Analysis of OsIQMs with OsCaMs
Previous studies have shown that IQM1 can interact with CaM5 in Arabidopsis [13]. To verify whether this interaction also exists in rice, we used yeast two-hybrid analysis to detect the interaction between OsIQMs with OsCaMs. Our results showed that AD-OsCaMs (OsCaM1, 2, 3) and BD-OsIQMs (OsIQM1, 2, 3, 4, 5, 6, 7, 8) co-transform into AH109 yeast cells, and all the transformants were found to grow well on SD/-Trp/-Leu/-Ade/-His medium, indicating that OsIQMs and OsCaMs interacted in yeast (Figure 6A).
2.7. Mutations in the IQ Motif of OsIQMs Result in the Loss of Affinity CaM Binding
In Arabidopsis, the IQ motif in AtIQM1 was identified to bind with CaM [13]. In order to verify whether the IQ motif can bind with CaM in rice, we used yeast two-hybrid analysis to examine the interaction between OsIQMsDelLQ with OsCaM1. Our results showed that when the recombinant plasmid BD-OsIQMsDelLQ and AD-OsCaM1 were transformed into the yeast AH109 strain, we can see all the transformants that were not able to grow on the SD/-Trp/-Leu/-Ade/-His medium (Figure 6B). The results suggested that the IQ motif is required for OsIQMs to combine with OsCaM1.
2.8. Expression Profiling of OsIQM Genes
To examine the transcript of OsIQM genes in the entire rice life cycle, the expression profiles at different developmental stages and organs were analyzed by quantitative real-time PCR (Figure 7). RNA was extracted from various organs and tissues of Zhonghua 11 (ZH11). The expression patterns of the OsIQM1 genes showed high expression in all tested organs and tissues. OsIQM2, OsIQM5 and OsIQM6 showed relatively low expression levels in all tested organs and tissues. OsIQM3 and OsIQM4 displayed higher expression in root, leaf sheaths and culm. OsIQM7 was predominantly expressed in root, and OsIQM8 showed high expression in root and culm.
In Arabidopsis, IQM1 was regulated by disease, ABA and JA [14,15]. However, no IQM genes responsive to stress and hormones have been reported in rice. We used qRT-PCR to analyze the expression levels of OsIQM family genes under stress and hormones treatments. As shown in Figure 8 and Figure S3, the OsIQM family have the same expression level under PEG and NaCl treatment. To realize the expression patterns of the eight OsIQM genes, we further revealed the expression levels of each gene for each period. Of the eight genes, four (OsIQM3, 5, 7 and 8) showed the higher transcript levels in the root treated with PEG, while three (OsIQM2, 3 and 5) showed the higher transcript levels in the shoot under the PEG treatment. In the root, these genes caused a marked change in the transcription levels at 0.5 h.
From the heat map of the qRT-PCR analysis results for the OsIQM genes, almost all genes were found to be ABA and MeJA responsive; however, some differences were observed among these genes (Figure 9 and Figure S4). Following ABA treatment, the transcript levels of three genes (OsIQM1, 3 and 8) were rapidly reduced in root and shoot, and OsIQM4 was downregulated in the shoot but upregulated in the root. OsIQM2 and OsIQM7 genes exhibited minor changes in expression of shoot and root at all time points except OsIQM7 was highly expressed at 0.5 h in the root. For OsIQM5 and OsIQM6, the expression level was initially upregulated to high levels during early treatment, and then downregulated in both root and shoot. Under MeJA treatment, most of the OsIQM genes (OsIQM1, 3, 4, 5, 6, 7 and 8 for the root, OsIQM1, 2, 3, 4, 5, 6 and 8 for the shoot) were inducible by MeJA abiotic stress. Among these, the expression levels of OsIQM1, 3, 4 and 8 were obviously downregulated in both root and shoot, while OsIQM6 exhibited minor changes in downregulated expression. By contrast, OsIQM5 was upregulated during early treatment which peaked at 0.5 h but downregulated at later time points for root, while OsIQM7 was obviously upregulated at all time points. For the shoot, OsIQM5 was obviously downregulated while OsIQM2 was gradually upregulated. These results indicated that certain OsIQM genes play important roles in regulating the responses to PEG, NaCl, ABA and MeJA stress.
2.9. Co-Expression Network Analysis of OsIQMs
To evaluate whether the genes co-expressed with OsIQMs involve in various biological processes, an extensive co-expression analysis of the eight OsIQMs was undertaken using the multiple guide gene search (MR = 10, hierarchy = 1) in the RiceFREND web servers [22]. The results indicated that OsIQM1 was mainly involved in plant–pathogen interaction pathways by WRKY24 (Figure 10). OsIQM2 was involved in nucleotide excision repair pathways (Figure 10). No hits were found for OsIQM3, OsIQM5 and OsIQM7.
3. Discussion
The plant-specific IQM gene family is widespread in many plants and are reported to participate in many crucial biological processes in plants [13,14,15,16]. The systematic and integrative analysis of IQM genes has been studied in Arabidopsis (Zhou et al., 2010). However, little is known about the rice IQM gene family. We focused on rice crop and performed genome-wide analysis to identify the IQM gene family in rice.
3.1. Evolution of the OsIQMs Gene Family in Rice
Genome-wide analysis of a gene family can characterize plant gene functions and facilitate the study of the evolution of genes and genomes. In the present study, based on the amino acid sequence of the conserved domain, eight IQM genes were identified in the rice genome. These were divided into three subfamilies based on their phylogenetic relationships with IQM proteins from Brachypodium distachyon, Oryza sativa, Arabidopsis thaliana, Populus trichocarpa, Zea mays, Glycine max and Solanum lycopersicum (Figure 1A). The phylogenetic analysis revealed the evolutionary relationships between the rice IQM proteins, and also showed that subfamily I contained the largest number (four) of rice IQM protein family members. Most closely related members within the same subfamily shared similarities in gene structure (Figure 2) and motif distribution (Figure 3), implying the reliability of the subfamily classification. Therefore, we speculated that the IQMs in each subfamily may have similar functions.
To further analyze the phylogenetic relationship of the IQM genes, we identified the putative orthologous and paralogous genes from members of our phylogenetic tree. Generally, orthologs are defined as genes from different genomes that were derived from a single ancestral gene and may have similar functions. In our study, the putative orthologous genes of OsIQMs (OsIQM1/Bd2g41490, OsIQM4/Bd2g33150, OsIQM5/Zm2g318843, OsIQM6/Zm2g168222 and OsIQM8/Bd4g42560) were identified (Figure 1A). Notably, the ortholog pair formed by eudicot and monocot was not found in the phylogenetic tree, indicating before the eudicot–monocot split, there were no ancestral IQM genes that existed.
In addition, paralogs are defined as genes that originate from an ancestor gene within the same genome produced by gene duplication events [23,24]. Therefore, gene duplication events play an important role in the evolution of a gene family [25,26]. In our study, we found that all OsIQM gene pairs exhibited segmental duplication events but not tandem duplication events (Figure 4A). This result indicated that segmental duplication played a leading role in the expansion of the rice IQM gene family. To better explain the patterns of expansion, estimates of the evolutionary rates are extremely useful. We estimated the Ks and Ka models of paralogous genes and calculated the approximate date of the duplication event. By calculating the duplication dates for the paralogous pairs, we concluded that all of the segmental duplication events in the OsIQM gene family occurred ranged from 11.9 to 19.8 Mya. Therefore, before the evolutionary expansion of the OsIQM gene family in the rice genome, five OsIQM genes were distributed on four chromosomes. From the separation of Zizaniinae and Oryzinae (approximately 20 million years ago) to the time where Oryza branched-off from the remaining Oryzeae genera (approximately 9 million years ago), OsIQM5 and OsIQM6 arose from OsIQM2 and OsIQM3 on chromosome 3, OsIQM8 arose from OsIQM7 on chromosome 11 through segmental duplication event. After the duplication evolutionary stages, the number of OsIQM gene family increased to eight genes.
3.2. The IQ Motif Plays an Important Role in the Interaction of OsIQM and OsCaM
Previous studies have revealed that five classes of IQ motif-containing protein family (CNGC, IQM, CAMTA, myosin and IQD) had divergence in the number and distribution of IQ motifs [11]. The IQ domain was related to the calmodulin-binding domain, and it was reported that IQD proteins interact with CaMs in maize [27], Chinese cabbage [28], moso bamboo [29] and Arabidopsis [30], IQM proteins interact with CaMs in Arabidopsis [13]. In our study, multiple sequence alignments of the amino acid sequences of eight OsIQM proteins revealed the presence of the IQ domain (Figure S1). Moreover, MEME analysis showed that motif one, motif two, motif three, motif four, motif five and motif six were present in almost all members of the OsIQM family and motif three contained the IQ motif. Next, we used yeast two-hybrid analysis to detect the interaction between OsCaM and OsIQMs. The results showed that OsIQMs interacted with OsCaM, which provide another evidence that OsIQMs protein might function throughout the CaM pathway. Previous studies have shown that AtIQM1 can bind with CaM5 via its IQ-motif. In order to further understand whether the IQ motif plays an important role in the interaction of OsIQM and OsCaM, we deleted the LQ of OsIQM to test the binding activity of OsIQM and OsCaM. The results indicated that the IQ motif is the key domain of OsIQM and determined the binding activity of OsIQM and OsCaM. These preliminary results of the OsIQM family genes may provide a foundation for further studies on the biochemical functions of IQM proteins in rice.
3.3. OsIQMs Respond to Abiotic and Biotic Stress and May Take Part in Stress Resistance Involving Stress-Related Genes
The evolutionary expansion of a gene family may result in functional diversification by gene duplication, and then perform the different expression profile of the gene family member [25,31]. To further investigate the possible functions of the OsIQMs in plant growth and development, the spatio-temporal expression patterns of the eight OsIQMs in ten different tissues and organs were detected by qRT-PCR (Figure 7). Based on our results, the OsIQM genes were differentially expressed at various developmental stages and tissues. OsIQM1 exhibit high expression levels in all tissues. By contrast, OsIQM2, OsIQM5 and OsIQM6 lowly expressed across all tissues, and indicated that these genes may work together with other proteins during plant growth and development. In addition, unlike most duplicated genes, the expression profiles of the duplicated OsIQM genes had distinct tissue specificities.
Jasmonic acid (JA) is an essential plant hormone that regulate plant growth and developmental processes as well as disease response signaling [32]. The catalases are involved in plant stress resistance by eliminating excess H2O2 in Arabidopsis and rice [33,34]. In Arabidopsis, IQM1 can positively regulate JA content and B. cinerea resistance by directly interacted with and promoted CATALASE2 (CAT2) expression and CAT2 enzyme activity, indicating that IQM1 is a key regulatory factor in the signaling of plant disease responses mediated by JA [14]. Therefore, we used qRT-PCR to investigate the expression patterns of OsIQMs family genes under MeJA treatments (Figure 9). The results demonstrated that most OsIQMs genes were either increased or repressed under the MeJA treatments, and we speculated that OsIQMs genes might have a similar biological function in the signaling of plant disease responses. It has been reported that WRKY transcription factors played an important role in abiotic stress responses [35,36] and biotic stress responses [37,38,39,40]. OsWRKY11, 24, 30 and 71 are transcriptional activators that positively regulates disease resistance in rice [35,41,42,43]. In this work, co-expression networks showed that seven OsIQM1 co-expressed with OsWRKY24, implying that OsIQM1 might partner with OsWRKY24 during plant disease response (Figure 10). Meanwhile, the results from the Genevestigator demonstrated that the expression of OsIQM1 was rapidly increased after infection of Magnaporthe oryzae (M. oryzae) (Figure S5). These results further demonstrated that OsIQM1 may be involved in disease resistance in rice, similar to AtIQM1. In addition, the phytohormone, ABA, plays a predominant role in regulating plant adaption to various abiotic stresses such as drought, salt or low temperature [44]. Zhou et al. verify that AtIQM4 plays key roles in modulating the responses to ABA, salt and osmotic stress during seed germination and post-germination growth [15]. Similarly, the expression of most OsIQMs members was regulated by ABA, PEG, NaCl. Thus, it is possible that these OsIQMs respond to abiotic and biotic stress and may be candidate genes for stress resistance in rice. The new information obtained in this study may provide new insights for further functional characterization of OsIQMs.
4. Materials and Methods
4.1. Identification of IQM Genes in Rice
The current study searched rice IQM (OsIQM) proteins using the Basic Local Alignment Search Tool algorithms program (BLASTP) in RAP-DB (http://rapdb.dna.affrc.go.jp/, accessed on 19 July 2020) and NCBI database (http://0-www-ncbi-nlm-nih-gov.brum.beds.ac.uk/, accessed on 19 July 2020), with the Arabidopsis IQM protein sequences used as query sequences. After manually removing the redundancy genes based on cluster-W alignment results, all candidate sequences were further confirmed using SMART (http://smart.embl-heidelberg.de/, accessed on 19 July 2020) for detecting the IQ domain [45]. Sequences of Arabidopsis thaliana IQM proteins were retrieved from the Arabidopsis Information Resource (TAIR, http://www.arabidopsis.org/, accessed on 19 July 2020), Brachypodium distachyon, Populus trichocarpa, Zea mays and Glycine max IQM protein sequences were downloaded from Phytozome v12.1, (http://www.phytozome.net/, accessed on 19 July 2020) and Solanum lycopersicum IQM protein sequences were retrieved from the tomato WGS chromosomes (2.40; SL2.40) (SGN http://solgenomics.net, accessed on 19 July 2020), respectively. Information on rice IQM genes, including the number of amino acids, open reading frame (ORF) lengths and location coordinates were acquired from RAP-DB. The physicochemical parameters of each rice proteins were generated by the program ExPASy (http://web.expasy.org/protparam/, accessed on 11 August 2020). The subcellular localization of the IQM proteins were predicted with the WoLF PSORT program (http://wolfpsort.org, accessed on 11 August 2020) using the amino acid sequences.
4.2. Chromosomal Localization and Gene Duplication
The chromosomal distribution of OsIQM genes was drafted by MapInspect software (http://www.plantbreeding.wur.nl/uk/software_mapinspect.html, accessed on 15 August 2020) according to their position information available in the Rice Genome Annotation Project (RGAP) database (http://rice.plantbiology.msu.edu/index.shtml, accessed on 15 August 2020). The current study analyzed the OsIQM gene duplication events, including tandem and segmental duplications. If the OsIQM gene were placed on duplicated chromosomal blocks available at MSU-RGA with the maximal length distance permitted between collinear gene pairs of 500 kb, they were produced by segmentally duplicated events. Tandem duplicates were determined when genes separated by five or fewer genes in a 100-kb region.
To further analyze the divergence of duplicated genes, the synonymous substitution rate (Ks) and non-synonymous substitution rate (Ka) were calculated using the software DnaSp. Based on a rate of λ (λ = 6.5 × 10−9 for rice) substitutions per synonymous site per year, the divergence time (T) was subsequently calculated using the equation T = Ks/2λ × 10−6 million years ago (Mya).
4.3. Protein Sequence Alignment and Phylogenetic Analysis
Multiple sequence alignment of all IQM protein and conserved domains (IQ domain) was performed using the Clustal X (version 1.83) program. To generate the unrooted phylogenetic tree, the neighbor-joining (NJ) method was used and with the p-distance substitution model in the MEGA X software. For bootstrap analysis we used 1000 replicates and the pairwise deletion option to obtain a support value for each branch.
4.4. Analysis of Gene and Protein Structure
For gene structure analysis, the coding sequence (CDS) and corresponding genomic sequences of OsIQM genes were downloaded using genome browser tool in RAP-DB, and we used the Gene Structures Display Server2.0 (GSDS2.0) (http://gsds.cbi.pku.edu.cn/, accessed on 1 September 2020) to analyze the exon/intron structure for individual IQM genes.
To investigate the conserved motifs of IQM proteins, the complete amino acid sequences were analyzed using MEME (Multiple Expectation Maximization for Motif Elicitation) (http://meme.nbcr.net/meme/cgibin/meme.cgi, accessed on 1 September 2020). The maximum number of motifs were defined as 10, the motif width was set to between 6 and 200 residues, respectively.
4.5. Yeast Two Hybrid Assay
The Matchmaker GAL4 two-hybrid system was used to test the interaction between IQMs and CaM. For yeast two-hybrid assay, the full-length CDS of CaM were separately cloned into pGADT7, the full length of three IQM CDSs was cloned into the pGBKT7 bait vector. They were co-transformed into AH109 yeast cells by the PEG/LiAC method. Interactions in yeast were determined on SD/-His-Trp-Leu-Ade plates. The co-transformants with pGBKT7-53 and pGADT7-T were used as positive control while pGBKT7-Lam and pGADT7-T were used as negative controls.
4.6. Effects of Mutations in IQ Motif of OsIQMs on Its CaM Binding
To confirm the effects of mutation of key amino acid residues in the IQ motif of IQM on its CaM binding, the LQ was deleted through the mutagenesis technology in vitro. The PCR products were subsequently treated with Dpn I (TaKaRa, Japan) to eliminate the methylation. The OsIQMsDelLQ fragment (OsIQM1Del140-141VQ, OsIQM2Del125-126VQ, OsIQM3Del112-113LQ, OsIQM4Del11-112VQ, OsIQM5Del103-104LQ, OsIQM6Del140-141LQ, OsIQM7Del43-44LQ, OsIQM1Del41-42LQ) was inserted into the pGBKT7 vector. Then CaM binding of the mutant proteins was analyzed via yeast two-hybrid assay described above.
4.7. Plant Materials and Growth Conditions
Rice (Oryza sativa L. ssp. japonica cv. Zhonghua 11) seedlings were used as the WT control in this study. In order to evaluate the spatio-temporal expression profiles of OsIQM genes under normal conditions, the rice seedlings were grown in a controlled paddy at the South China Botanical Garden during natural growing seasons (March to July). For various treatments, seeds were surface-sterilized with ethanol (70% v/v) for 1 min and then with 5% NaOCl solution for 50 min, followed by several rinses with sterile water and cultured in Hoagland’s Solution at 28 °C under 14 h light and 10 h dark for 2 weeks. For analysis of gene expression under abiotic stresses, 2-week-old ZH11 seedlings were exposed to the following treatments: salinity (150 mM NaCl) and PEG (20% PEG6000) [46]. For analysis of gene expression with hormone, ABA (50 μM) and MeJA (50 μM) were added. Shoot and root were sampled at 0, 0.5, 2, 6, 12 and 24 h after treatment, then immediately frozen in liquid nitrogen and stored at −80 °C [46].
4.8. RNA Isolation and Quantitative Real-Time PCR
The total RNA was extracted from different rice samples according to the manufacturer’s protocols of RNAiso Plus (Takara, Code No. 9108). M-MLV Reverse Transcriptase (Promega, Cat#M1701) was used for reverse transcription. qRT-PCR was performed on 384-well plates with SYBR Premix Ex Taq II (Takara, RR820A) using a Roche Light Cycler 480 Real-Time PCR system. eEF-1a were used as internal controls for mRNA and the relative expression levels of genes were calculated using the 2−△△CT method. The experiments were performed in triplicate. All primers used are listed in Table S1.
4.9. Subcellular Localization Assays
A green fluorescent protein (GFP) fusion protein was constructed using full-length IQM CDS with a C-terminal fusion of the GFP clone under the control of a CaMV35S promoter [47]. Arabidopsis protoplast isolation and transformation were conducted as previously described [48]. The protoplasts were incubated at 22 °C for 12 h in the dark after transfection, and the subcellular distribution of the GFP fusion protein was examined using a confocal laser scanning microscope. Excitation was achieved using an argon laser at 488 nm (GFP), and the emission of GFP was detected from 492 to 550 nm. The auto-fluorescence of chlorophyll was simultaneously detected between 650 and 730 nm. The images presented are average projections of 8–20 optical sections [47].
4.10. Co-Expression Network Analysis
The OsIQMs was employed as a guide gene in Rice FREND (http://ricefrend.dna.affrc.go.jp/, accessed on 1 September 2020) [22]. In Rice FREND, Mutual Rank (MR) values were assessed between OsIQMs and co-expressed genes.
5. Conclusions
The current study systematically identified and characterized the plant-specific IQM gene family in rice. In this study, eight IQM genes identified in the rice genome were classified into three subfamilies (I–III). Gene structure and protein motif analyses showed that these genes are relatively conserved within each subfamily. Furthermore, quantitative real-time PCR analysis of eight OsIQMs genes showed that most IQM genes were responsive to PEG, NaCl, MeJA and ABA treatment, suggesting their crucial roles in biotic and abiotic stress response. Additionally, yeast two-hybrid assay showed that OsIQMs can interact with OsCaMs, and the IQ motif of OsIQMs is required for OsIQMs to combine with OsCaMs. Our preliminary results may be useful for future elucidating the role of IQMs in rice, bringing us one step closer to understanding the important biological functions of these proteins.
Supplementary Materials
The following are available online at https://0-www-mdpi-com.brum.beds.ac.uk/article/10.3390/plants10091949/s1, Figure S1: amino acid sequence alignments of IQ domains in rice IQM protein sequences. The multiple alignment results indicate the highly conserved IQM domains among the eight identified rice IQM protein sequences. The IQ motif was in the red box. Figure S2: the conserved amino acid sequences and length of each motif. Figure S3: expression patterns of eight OsIQM genes under PEG and NaCl stress, as revealed by qRT-PCR. Figure S4: expression patterns of eight OsIQM genes under MeJA and ABA stress, as revealed by qRT-PCR. Figure S5: the expression of OsIQM1 was rapidly increased after infection of M. oryzae from the Genevestigator. Table S1: all primes in this study.
Author Contributions
C.T. and T.F., designed the research. T.F., T.L. and C.X., performed experiments. T.F., C.T., Y.Z., analyzed data. T.F. and C.T., wrote the manuscript. All authors have read and agreed to the published version of the manuscript.
Funding
This research was funded by the National Natural Science Foundation of China (31601817/311770342), the Natural Science Foundation of Guangdong Province (2020A1515011423).
Institutional Review Board Statement
Not applicable.
Informed Consent Statement
Not applicable.
Data Availability Statement
Not applicable.
Conflicts of Interest
Authors declare that there is no conflict of interest.
References
- DeFalco, T.A.; Bender, K.W.; Snedden, W.A. Breaking the code: Ca2+ sensors in plant signalling. Biochem. J. 2010, 425, 27–40. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ng, C.K.Y.; Mcainsh, M.R.; Gray, J.E.; Hunt, L.; Leckie, C.P.; Mills, L.; Hetherington, A.M. Calcium-based signalling systems in guard cells. New Phytol. 2001, 151, 109–120. [Google Scholar] [CrossRef]
- Reddy, A.S.N.; Ali, G.S.; Celesnik, H.; Day, I.S. Coping with Stresses: Roles of Calcium- and Calcium/Calmodulin-Regulated Gene Expression. Plant Cell. 2011, 23, 2010–2032. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Batistic, O.; Kudla, J. Analysis of calcium signaling pathways in plants. BBA-Gen. Subj. 2012, 1820, 1283–1293. [Google Scholar] [CrossRef]
- Bouche, N.; Yellin, A.; Snedden, W.A.; Fromm, H. Plant-specific calmodulin-binding proteins. Annu. Rev. Plant Biol. 2005, 56, 435–466. [Google Scholar] [CrossRef] [PubMed]
- Luan, S.; Kudla, J.; Rodriguez-Concepcion, M.; Yalovsky, S.; Gruissem, W. Calmodulins and calcineurin B-like proteins: Calcium sensors for specific signal response coupling in plants. Plant Cell. 2002, 14, S389–S400. [Google Scholar] [CrossRef] [Green Version]
- Reddy, A.S.N. Calcium: Silver bullet in signaling. Plant Sci. 2001, 160, 381–404. [Google Scholar] [CrossRef]
- Yang, T.B.; Poovaiah, B.W. Calcium/calmodulin-mediated signal network in plants. Trends Plant Sci. 2003, 8, 505–512. [Google Scholar] [CrossRef]
- Bhattacharya, S.; Bunick, C.G.; Chazin, W.J. Target selectivity in EF-hand calcium binding proteins. BBA-Mol. Cell Res. 2004, 1742, 69–79. [Google Scholar] [CrossRef] [Green Version]
- Abel, S.; Savchenko, T.; Levy, M. Genome-wide comparative analysis of the IQD gene families in Arabidopsis thaliana and Oryza sativa. BMC Evol. Biol. 2005, 5, 72. [Google Scholar] [CrossRef] [Green Version]
- Zhou, Y.P.; Chen, Y.Z.; Yamamoto, K.T.; Duan, J.; Tian, C.E. Sequence and expression analysis of the Arabidopsis IQM family. Acta Physiol. Plant 2010, 32, 191–198. [Google Scholar] [CrossRef]
- Sävenstrand, H.; Strid, A. Six genes strongly regulated by mercury in Pisum sativum roots. Plant Physiol. Biochem. 2004, 42, 135–142. [Google Scholar] [CrossRef] [PubMed]
- Zhou, Y.P.; Duan, J.; Fujibe, T.; Yamamoto, K.T.; Tian, C.E. AtIQM1, a novel calmodulin-binding protein, is involved in stomatal movement in Arabidopsis. Plant Mol. Biol. 2012, 79, 333–346. [Google Scholar] [CrossRef]
- Lv, T.; Li, X.; Fan, T.; Luo, H.; Xie, C.; Zhou, Y.; Tian, C.E. The Calmodulin-Binding Protein IQM1 Interacts with CATALASE2 to Affect Pathogen Defense. Plant Physiol. 2019, 181, 1314–1327. [Google Scholar] [CrossRef] [PubMed]
- Zhou, Y.P.; Wu, J.H.; Xiao, W.H.; Chen, W.; Chen, Q.H.; Fan, T.; Xie, C.P.; Tian, C.E. Arabidopsis IQM4, a Novel Calmodulin-Binding Protein, Is Involved With Seed Dormancy and Germination in Arabidopsis. Front. Plant Sci. 2018, 9, 721. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gong, L.P.; Cheng, J.Z.; Zhou, Y.P.; Huang, X.L.; Tian, C.E. Disruption of IQM5 delays flowering possibly through modulating the juvenile-to-adult transition. Acta Physiol. Plant. 2016, 39, 21. [Google Scholar] [CrossRef]
- Mello, B. Estimating TimeTrees with MEGA and the TimeTree Resource. Mol. Biol. Evol. 2018, 35, 2334–2342. [Google Scholar] [CrossRef] [Green Version]
- Cannon, S.B.; Mitra, A.; Baumgarten, A.; Young, N.D.; May, G. The roles of segmental and tandem gene duplication in the evolution of large gene families in Arabidopsis thaliana. BMC Plant Biol. 2004, 4, 10. [Google Scholar] [CrossRef] [Green Version]
- Hurst, L.D. The Ka/Ks ratio: Diagnosing the form of sequence evolution. Trends Genet. TIG 2002, 18, 486. [Google Scholar] [CrossRef]
- Shiu, S.H.; Karlowski, W.M.; Pan, R.S.; Tzeng, Y.H.; Mayer, K.F.X.; Li, W.H. Comparative analysis of the receptor-like kinase family in Arabidopsis and rice. Plant Cell 2004, 16, 1220–1234. [Google Scholar] [CrossRef] [Green Version]
- Guo, Y.L.; Ge, S. Molecular phylogeny of Oryzeae (Poaceae) based on DNA sequences from chloroplast, mitochondrial, and nuclear genomes. Am. J. Bot. 2005, 92, 1548–1558. [Google Scholar] [CrossRef] [Green Version]
- Sato, Y.; Namiki, N.; Takehisa, H.; Kamatsuki, K.; Minami, H.; Ikawa, H.; Ohyanagi, H.; Sugimoto, K.; Itoh, J.I.; Antonio, B.A.; et al. RiceFREND: A platform for retrieving coexpressed gene networks in rice. Nucleic Acids Res. 2013, 41, D1214–D1221. [Google Scholar] [CrossRef] [Green Version]
- Tatusov, R.L.; Koonin, E.V.; Lipman, D.J. A genomic perspective on protein families. Science 1997, 278, 631–637. [Google Scholar] [CrossRef] [Green Version]
- Sonnhammer, E.L.; Koonin, E.V. Orthology, paralogy and proposed classification for paralog subtypes. Trends Genet. TIG 2002, 18, 619–620. [Google Scholar] [CrossRef]
- Hughes, A.L. The evolution of functionally novel proteins after gene duplication. Proc. Biol. Sci. 1994, 256, 119–124. [Google Scholar] [CrossRef] [PubMed]
- Adams, K.L.; Wendel, J.F. Polyploidy and genome evolution in plants. Curr. Opin. Plant Biol. 2005, 8, 135–141. [Google Scholar] [CrossRef] [PubMed]
- Cai, R.H.; Zhang, C.S.; Zhao, Y.; Zhu, K.J.; Wang, Y.F.; Jiang, H.Y.; Xiang, Y.; Cheng, B.J. Genome-wide analysis of the IQD gene family in maize. Mol. Genet. Genom. 2016, 291, 543–558. [Google Scholar] [CrossRef] [PubMed]
- Yuan, J.P.; Liu, T.K.; Yu, Z.H.; Li, Y.; Ren, H.B.; Hou, X.L.; Li, Y. Genome-wide analysis of the Chinese cabbage IQD gene family and the response of BrIQD5 in drought resistance. Plant Mol. Biol. 2019, 99, 603–620. [Google Scholar] [CrossRef] [PubMed]
- Wu, M.; Li, Y.; Chen, D.M.; Liu, H.L.; Zhu, D.Y.; Xiang, Y. Genome-wide identification and expression analysis of the IQD gene family in moso bamboo (Phyllostachys edulis). Sci. Rep. 2016, 6, 24520. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Levy, M.; Wang, Q.M.; Kaspi, R.; Parrella, M.P.; Abel, S. Arabidopsis IQD1, a novel calmodulin-binding nuclear protein, stimulates glucosinolate accumulation and plant defense. Plant J. 2005, 43, 79–96. [Google Scholar] [CrossRef]
- Lynch, M.; Conery, J.S. The evolutionary fate and consequences of duplicate genes. Science 2000, 290, 1151–1155. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Spoel, S.H.; Dong, X. Making sense of hormone crosstalk during plant immune responses. Cell Host Microbe 2008, 3, 348–351. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Su, T.; Wang, P.P.; Li, H.J.; Zhao, Y.W.; Lu, Y.; Dai, P.; Ren, T.Q.; Wang, X.F.; Li, X.Z.; Shao, Q.; et al. The Arabidopsis catalase triple mutant reveals important roles of catalases and peroxisome-derived signaling in plant development. J. Integr. Plant Biol. 2018, 60, 591–607. [Google Scholar] [CrossRef] [PubMed]
- Zafar, S.A.; Hameed, A.; Ashraf, M.; Khan, A.S.; Zia-ul-Qamar; Li, X.Y.; Siddique, K.H.M. Agronomic, physiological and molecular characterisation of rice mutants revealed the key role of reactive oxygen species and catalase in high-temperature stress tolerance. Funct. Plant Biol. 2020, 47, 440–453. [Google Scholar] [CrossRef] [PubMed]
- Kim, C.Y.; Vo, K.T.X.; Nguyen, C.D.; Jeong, D.H.; Lee, S.K.; Kumar, M.; Kim, S.R.; Park, S.H.; Kim, J.K.; Jeon, J.S. Functional analysis of a cold-responsive rice WRKY gene, OsWRKY71. Plant Biotechnol. Rep. 2016, 10, 13–23. [Google Scholar] [CrossRef]
- Raineri, J.; Wang, S.H.; Peleg, Z.; Blumwald, E.; Chan, R.L. The rice transcription factor OsWRKY47 is a positive regulator of the response to water deficit stress. Plant Mol. Biol. 2015, 88, 401–413. [Google Scholar] [CrossRef]
- Choi, C.; Hwang, S.H.; Fang, I.R.; Il Kwon, S.; Park, S.R.; Ahn, I.; Kim, J.B.; Hwang, D.J. Molecular characterization of Oryza sativa WRKY6, which binds to W-box-like element 1 of the Oryza sativa pathogenesis-related (PR) 10a promoter and confers reduced susceptibility to pathogens. New Phytol. 2015, 208, 846–859. [Google Scholar] [CrossRef]
- Han, M.; Ryu, H.S.; Kim, C.Y.; Park, D.S.; Ahn, Y.K.; Jeon, J.S. OsWRKY30 is a transcription activator that enhances rice resistance to the Xanthomonas oryzae pathovar oryzae. J. Plant Biol. 2013, 56, 258–265. [Google Scholar] [CrossRef]
- Hwang, S.H.; Kwon, S.I.; Jang, J.Y.; Fang, I.L.; Lee, H.; Choi, C.; Park, S.; Ahn, I.; Bae, S.C.; Hwang, D.J. OsWRKY51, a rice transcription factor, functions as a positive regulator in defense response against Xanthomonas oryzae pv. oryzae. Plant Cell Rep. 2016, 35, 1975–1985. [Google Scholar] [CrossRef] [PubMed]
- Lan, A.; Huang, J.; Zhao, W.; Peng, Y.; Chen, Z.; Kang, D. A salicylic acid-induced rice (Oryza sativa L.) transcription factor OsWRKY77 is involved in disease resistance of Arabidopsis thaliana. Plant Biol. 2013, 15, 452–461. [Google Scholar] [CrossRef]
- Yokotani, N.; Shikata, M.; Ichikawa, H.; Mitsuda, N.; Ohme-Takagi, M.; Minami, E.; Nishizawa, Y. OsWRKY24, a blast-disease responsive transcription factor, positively regulates rice disease resistance. J. Gen. Plant Pathol. 2018, 84, 85–91. [Google Scholar] [CrossRef]
- Lee, H.; Cha, J.; Choi, C.; Choi, N.; Ji, H.S.; Park, S.R.; Lee, S.; Hwang, D.J. Rice WRKY11 Plays a Role in Pathogen Defense and Drought Tolerance. Rice 2018, 11, 5. [Google Scholar] [CrossRef]
- Peng, X.X.; Hu, Y.J.; Tang, X.K.; Zhou, P.L.; Deng, X.B.; Wang, H.H.; Guo, Z.J. Constitutive expression of rice WRKY30 gene increases the endogenous jasmonic acid accumulation, PR gene expression and resistance to fungal pathogens in rice. Planta 2012, 236, 1485–1498. [Google Scholar] [CrossRef]
- Zhao, J.F.; Zhao, L.L.; Zhang, M.; Zafar, S.A.; Fang, J.J.; Li, M.; Zhang, W.H.; Li, X.Y. Arabidopsis E3 Ubiquitin Ligases PUB22 and PUB23 Negatively Regulate Drought Tolerance by Targeting ABA Receptor PYL9 for Degradation. Int. J. Mol. Sci. 2017, 18, 1841. [Google Scholar] [CrossRef]
- Letunic, I.; Copley, R.R.; Schmidt, S.; Ciccarelli, F.D.; Doerks, T.; Schultz, J.; Ponting, C.P.; Bork, P. SMART 4.0: Towards genomic data integration. Nucleic Acids Res. 2004, 32, D142–D144. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yang, W.; Fan, T.; Hu, X.Y.; Cheng, T.H.; Zhang, M.Y. Overexpressing osa-miR171c decreases salt stress tolerance in rice. J. Plant Biol. 2017, 60, 485–492. [Google Scholar] [CrossRef]
- Fan, T.; Li, X.M.; Yang, W.; Xia, K.F.; Jie, O.Y.; Zhang, M.Y. Rice osa-miR171c Mediates Phase Change from Vegetative to Reproductive Development and Shoot Apical Meristem Maintenance by Repressing Four OsHAM Transcription Factors. PLoS ONE 2015, 10, e0125833. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yoo, S.D.; Cho, Y.H.; Sheen, J. Arabidopsis mesophyll protoplasts: A versatile cell system for transient gene expression analysis. Nat. Protoc. 2007, 2, 1565–1572. [Google Scholar] [CrossRef] [PubMed] [Green Version]
Figure 1.
Phylogenetic tree and distribution of IQM protein from 7 plant species. (A) The full-length amino acid sequences of the IQM proteins from Brachypodium distachyon, Oryza sativa, Arabidopsis thaliana, Populus trichocarpa, Zea mays, Glycine max and Solanum lycopersicum were aligned using ClustalX 1.83, and the phylogenetic tree was constructed by the NJ method with MEGA X. The number of bootstrap values was 1000 replicates. Red—subfamily I; blue—subfamily II; green—subfamily III. IQM proteins from the same species were marked with geometrical patterning: hollow circle—Arabidopsis thaliana; filled circle—Oryza sativa; hollow triangle—Brachypodium distachyon; filled inverse-triangle—Glycine max; filled triangle—Solanum lycopersicum; filled quadrate—Populus trichocarpa; hollow quadrate—Zea mays. (B) A percentage representation of IQM proteins across the plant species in each subfamily.
Figure 1.
Phylogenetic tree and distribution of IQM protein from 7 plant species. (A) The full-length amino acid sequences of the IQM proteins from Brachypodium distachyon, Oryza sativa, Arabidopsis thaliana, Populus trichocarpa, Zea mays, Glycine max and Solanum lycopersicum were aligned using ClustalX 1.83, and the phylogenetic tree was constructed by the NJ method with MEGA X. The number of bootstrap values was 1000 replicates. Red—subfamily I; blue—subfamily II; green—subfamily III. IQM proteins from the same species were marked with geometrical patterning: hollow circle—Arabidopsis thaliana; filled circle—Oryza sativa; hollow triangle—Brachypodium distachyon; filled inverse-triangle—Glycine max; filled triangle—Solanum lycopersicum; filled quadrate—Populus trichocarpa; hollow quadrate—Zea mays. (B) A percentage representation of IQM proteins across the plant species in each subfamily.

Figure 2.
Phylogenetic relationship and exon–intron organization of rice IQM genes. The unrooted phylogenetic tree of eight rice IQM proteins was constructed by the NJ method with 1000 bootstrap replicates. Exons and introns are represented by yellow boxes and black lines, respectively. Untranslated regions (UTRs) are indicated by blue boxes. The sizes of each IQM gene can be estimated using the scale at the bottom.
Figure 2.
Phylogenetic relationship and exon–intron organization of rice IQM genes. The unrooted phylogenetic tree of eight rice IQM proteins was constructed by the NJ method with 1000 bootstrap replicates. Exons and introns are represented by yellow boxes and black lines, respectively. Untranslated regions (UTRs) are indicated by blue boxes. The sizes of each IQM gene can be estimated using the scale at the bottom.

Figure 3.
Motif distribution in IQM proteins of rice. Motifs of the OsIQM proteins were identified by the online MEME program. Each motif was represented by a different colored box with its serial number in central of the box. The colored boxes were ordered manually according to the results of the MEME server. The length of each colored box does not represent the actual motif size in the corresponding proteins.
Figure 3.
Motif distribution in IQM proteins of rice. Motifs of the OsIQM proteins were identified by the online MEME program. Each motif was represented by a different colored box with its serial number in central of the box. The colored boxes were ordered manually according to the results of the MEME server. The length of each colored box does not represent the actual motif size in the corresponding proteins.

Figure 4.
Chromosomal distribution and segmental duplication events of IQM genes in rice. (A) Eight OsIQM genes are mapped to 7 of the 12 rice chromosomes. The nearest SSR markers of eight OsIQM genes were mapped to 7 rice chromosomes. The duplicated paralogous pairs of OsIQM genes were connected with red lines. Chromosome numbers were located at the bottom of each vertical bar. Different colored circles on the left of the gene name represented corresponding subfamily which this gene belongs to. Red—subfamily I; blue—subfamily II; green—subfamily III. (B,C) Distribution of Ka/Ks values (B) and duplication date (MY) (C) were obtained from paralogous gene-pairs in the rice genome.
Figure 4.
Chromosomal distribution and segmental duplication events of IQM genes in rice. (A) Eight OsIQM genes are mapped to 7 of the 12 rice chromosomes. The nearest SSR markers of eight OsIQM genes were mapped to 7 rice chromosomes. The duplicated paralogous pairs of OsIQM genes were connected with red lines. Chromosome numbers were located at the bottom of each vertical bar. Different colored circles on the left of the gene name represented corresponding subfamily which this gene belongs to. Red—subfamily I; blue—subfamily II; green—subfamily III. (B,C) Distribution of Ka/Ks values (B) and duplication date (MY) (C) were obtained from paralogous gene-pairs in the rice genome.

Figure 5.
Subcellular localization of OsIQMs in Arabidopsis protoplasts. Bars = 5 μm. Bright, bright-field image; GFP—GFP fluorescence image; Chlorophyll—chlorophyll autofluorescence; merged—merged bright-field, GFP fluorescence and chlorophyll autofluorescence images.
Figure 5.
Subcellular localization of OsIQMs in Arabidopsis protoplasts. Bars = 5 μm. Bright, bright-field image; GFP—GFP fluorescence image; Chlorophyll—chlorophyll autofluorescence; merged—merged bright-field, GFP fluorescence and chlorophyll autofluorescence images.

Figure 6.
OsIQMs interaction with OsCaMs and exploration of the calmodulin-binding site of OsIQMs. (A) The bait construct of pGBKT7-OsIQMs and prey construct were co-transformed into the yeast strain AH109 and then were examined on SD/-Trp/-Leu and SD/-Trp/-Leu/-His/-Ade plates. (B) The interaction between OsCaM1 and OsIQMsDelLQ. Positive control: pGBKT7-53 and pGADT7-T; Negative control: pGBKT7-Lam and pGADT7-T.
Figure 6.
OsIQMs interaction with OsCaMs and exploration of the calmodulin-binding site of OsIQMs. (A) The bait construct of pGBKT7-OsIQMs and prey construct were co-transformed into the yeast strain AH109 and then were examined on SD/-Trp/-Leu and SD/-Trp/-Leu/-His/-Ade plates. (B) The interaction between OsCaM1 and OsIQMsDelLQ. Positive control: pGBKT7-53 and pGADT7-T; Negative control: pGBKT7-Lam and pGADT7-T.

Figure 7.
Heat map of the real-time quantitative PCR (qRT-PCR) analysis results of OsIQMs genes across different tissues analyzed, with three biological and technical replicates. The scale representing the relative signal intensity values is shown above. 2L-R—roots of 2-leaf stage seedlings; 2L-S—2-leaf stage seedlings; 10L-SA—shoot apex of 10-leaf stage seedling; ≤1P—developing panicles with a length of ≤1 cm; BP—booting panicle.
Figure 7.
Heat map of the real-time quantitative PCR (qRT-PCR) analysis results of OsIQMs genes across different tissues analyzed, with three biological and technical replicates. The scale representing the relative signal intensity values is shown above. 2L-R—roots of 2-leaf stage seedlings; 2L-S—2-leaf stage seedlings; 10L-SA—shoot apex of 10-leaf stage seedling; ≤1P—developing panicles with a length of ≤1 cm; BP—booting panicle.

Figure 8.
Heat map of the real-time quantitative PCR (qRT-PCR) analysis results of OsIQMs genes in shoots and roots under PEG and NaCl treatment, with three biological and technical replicates. The scale representing the relative signal intensity values is shown above.
Figure 8.
Heat map of the real-time quantitative PCR (qRT-PCR) analysis results of OsIQMs genes in shoots and roots under PEG and NaCl treatment, with three biological and technical replicates. The scale representing the relative signal intensity values is shown above.

Figure 9.
Heat map of the real-time quantitative PCR (qRT-PCR) analysis results of OsIQMs genes in leaves under ABA and MeJA treatment, with three biological and technical replicates. The scale representing the relative signal intensity values is shown above.
Figure 9.
Heat map of the real-time quantitative PCR (qRT-PCR) analysis results of OsIQMs genes in leaves under ABA and MeJA treatment, with three biological and technical replicates. The scale representing the relative signal intensity values is shown above.

Figure 10.
Co-expression network of the eight OsIQMs. The numbers represent the corresponding OsIQMs genes. The KEGG pathways are highlighted as dots: red—nucleotide excision repair (osa03420); blue—plant–pathogen interaction (osa04626).
Figure 10.
Co-expression network of the eight OsIQMs. The numbers represent the corresponding OsIQMs genes. The KEGG pathways are highlighted as dots: red—nucleotide excision repair (osa03420); blue—plant–pathogen interaction (osa04626).


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Table 1.
Eight identified members of the rice IQM gene family.
| Locus ID | Gene Name | Chr a | Pos b | ORF Length (bp) | Number of Amino Acid | Molecular Weight (Mw/Da) | Theoretical pI | FL-cDNAc | WoLF PSORT c |
|---|---|---|---|---|---|---|---|---|---|
| Os01g0570800 | OsIQM1 | 1 | 21900067 | 1710 | 570 | 63,264.22 | 9.15 | AB190923 | CH |
| Os03g0167200 | OsIQM2 | 3 | 3631183 | 1833 | 611 | 68,581.08 | 9.28 | AK066554 | N |
| Os03g0374500 | OsIQM3 | 3 | 14739099 | 1734 | 578 | 63,230.78 | 6.43 | AK071310 | N |
| Os05g0197300 | OsIQM4 | 5 | 5996776 | 1671 | 557 | 61,573.53 | 8.89 | AK106389 | N |
| Os07g0633400 | OsIOM5 | 7 | 26287974 | 1758 | 586 | 65,197.82 | 6.19 | AK071894 | N |
| Os10g0411500 | OsIQM6 | 10 | 14319443 | 1950 | 650 | 73,057.63 | 8.25 | AK072572 | N |
| Os11g0151002 | OsIQM7 | 11 | 2367903 | 1425 | 475 | 52,247.14 | 9.15 | AK102404 | N |
| Os12g0149900 | OsIQM8 | 12 | 2451806 | 1434 | 477 | 52,997.92 | 8.37 | AB164644 | N |
a—the chromosome in which the gene is located. b—the start position of the gene on the chromosome. c—CH chloroplasts, N nucleus.
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Fan, T.; Lv, T.; Xie, C.; Zhou, Y.; Tian, C. Genome-Wide Analysis of the IQM Gene Family in Rice (Oryza sativa L.). Plants 2021, 10, 1949. https://0-doi-org.brum.beds.ac.uk/10.3390/plants10091949
AMA Style
Fan T, Lv T, Xie C, Zhou Y, Tian C. Genome-Wide Analysis of the IQM Gene Family in Rice (Oryza sativa L.). Plants. 2021; 10(9):1949. https://0-doi-org.brum.beds.ac.uk/10.3390/plants10091949
Chicago/Turabian StyleFan, Tian, Tianxiao Lv, Chuping Xie, Yuping Zhou, and Changen Tian. 2021. "Genome-Wide Analysis of the IQM Gene Family in Rice (Oryza sativa L.)" Plants 10, no. 9: 1949. https://0-doi-org.brum.beds.ac.uk/10.3390/plants10091949
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.