
Photoprotective Agents Obtained from Aromatic Plants Grown in Colombia: Total Phenolic Content, Antioxidant Activity, and Assessment of Cytotoxic Potential in Cancer Cell Lines of Cymbopogon flexuosus L. and Tagetes lucida Cav. Essential Oils



Abstract
:1. Introduction
2. Results
2.1. Quantification of Total Phenolic Content in EOs
2.2. Assessment of UV Protective Properties
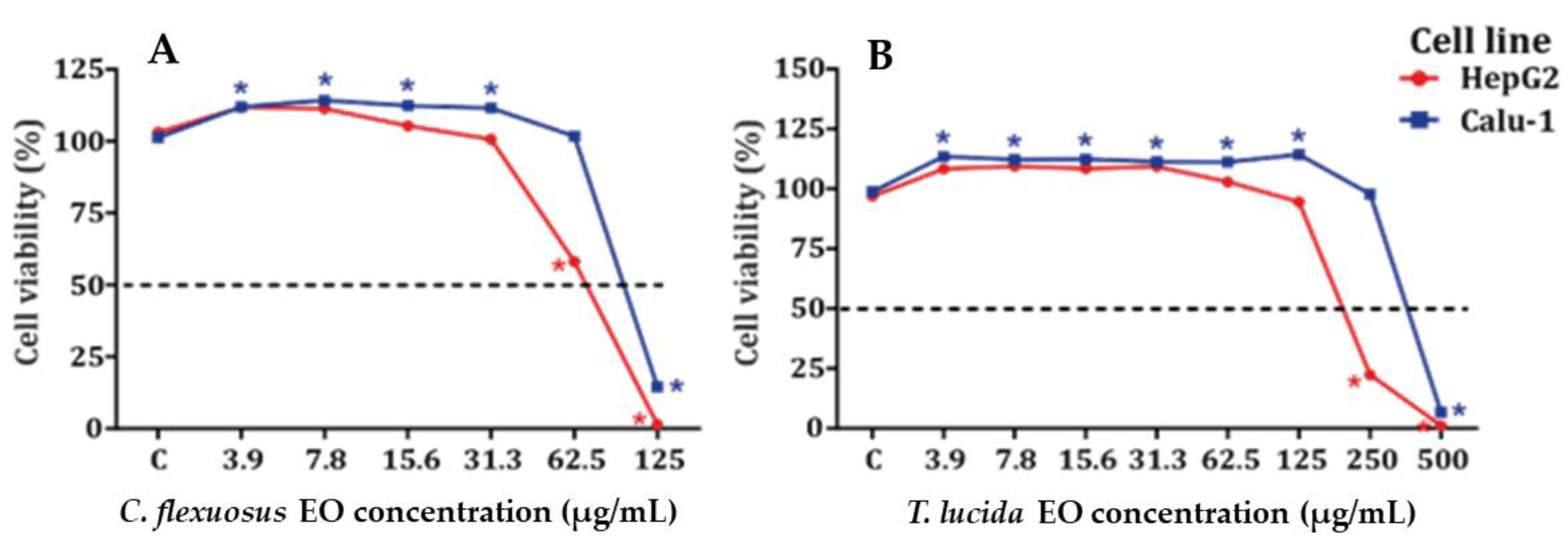
2.3. Determination of the Cytotoxic Potential of HepG2 and Calu-1 Cells in the MTT Viability Assay
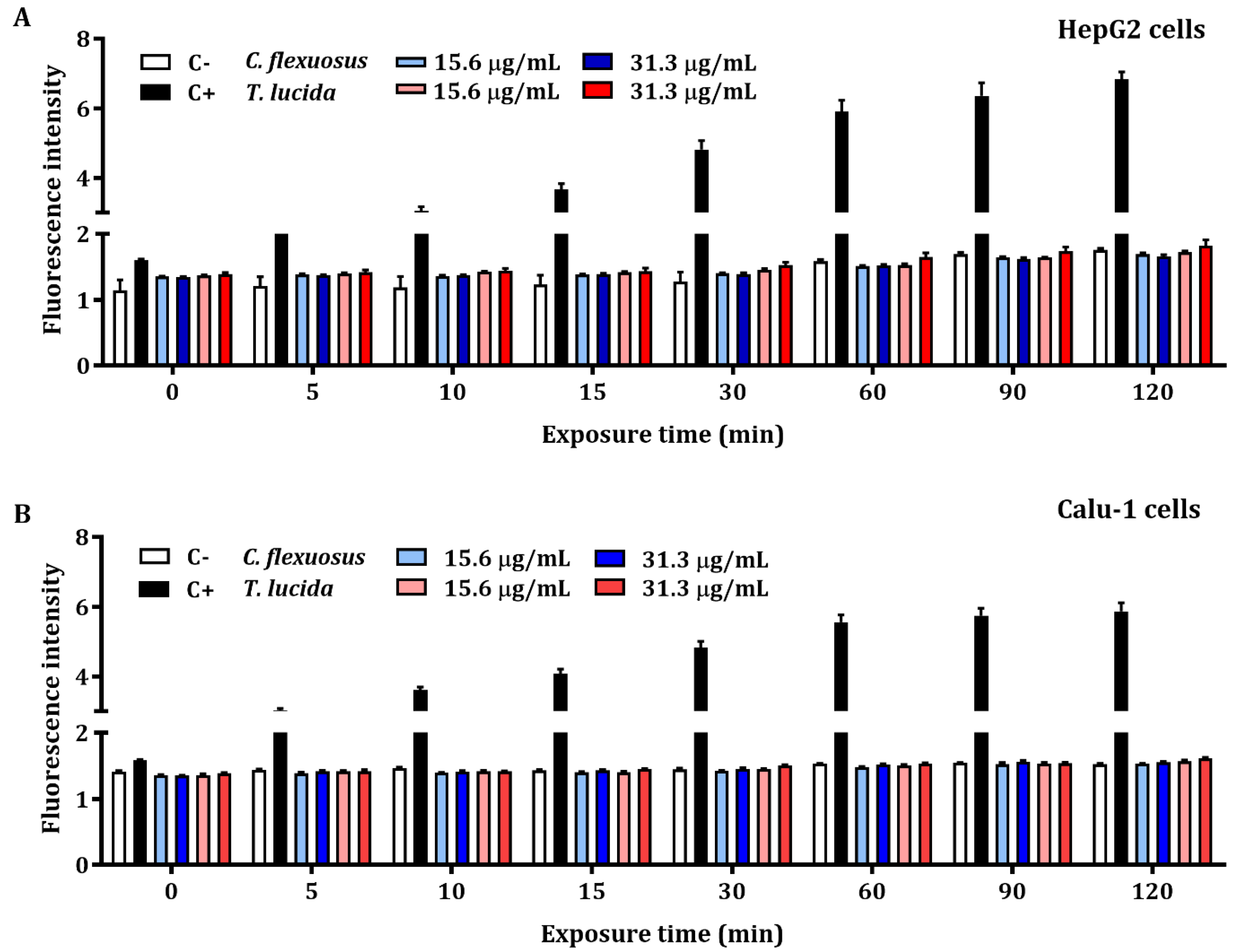
2.4. ROS Detection Assay of EOs in HepG2 and Calu-1 Cells
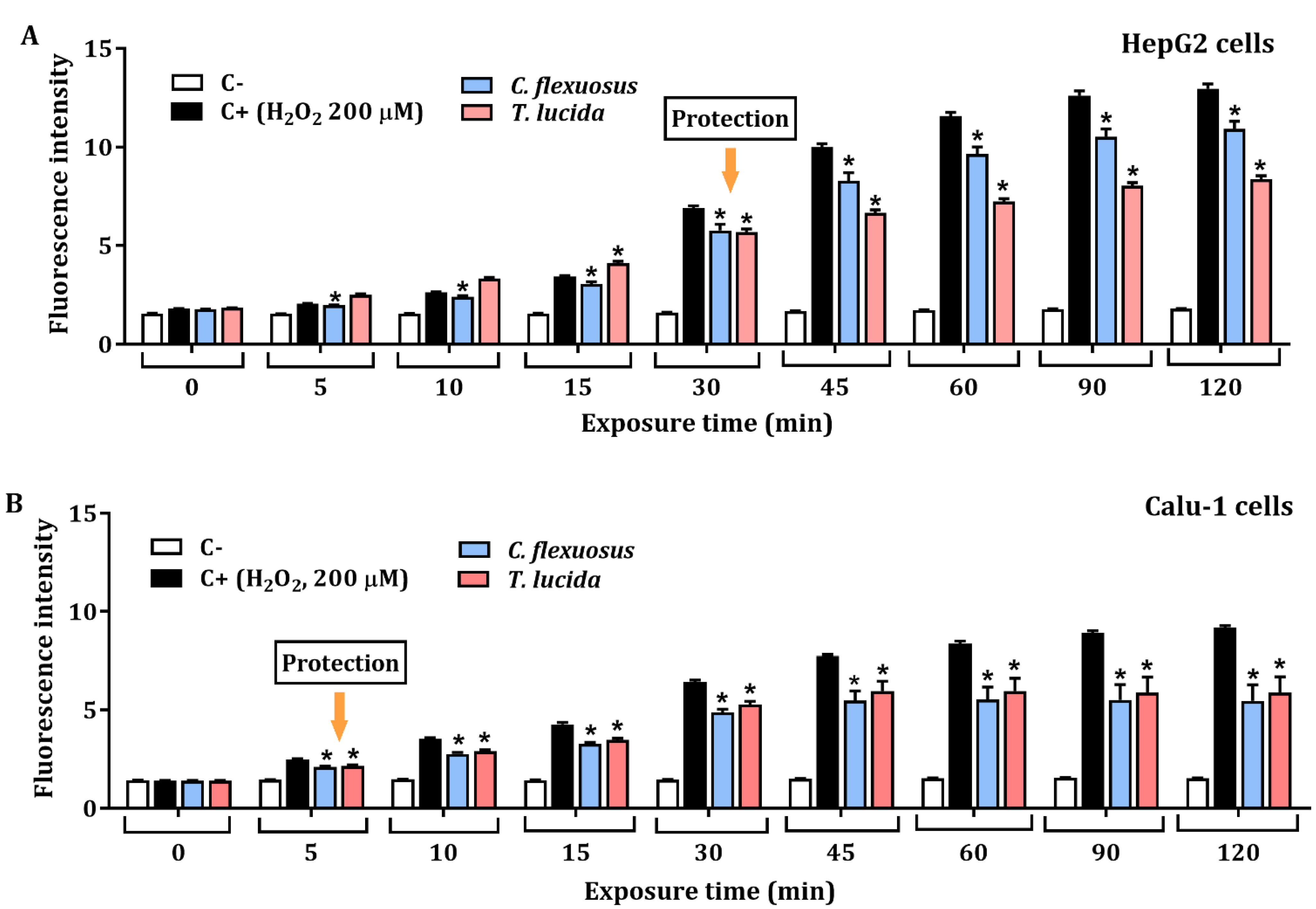
2.5. Assessment of Protective Effect against H2O2-Induced ROS
3. Discussion
4. Materials and Methods
4.1. Plant Material Collection, Extraction, and Characterization of EOs
4.2. Determination of Total Phenolic Content (TPC)
4.3. In Vitro Assessment of the UV Protective Properties of EOs
4.3.1. Sun Protection Factor (SPF)
4.3.2. Critical Wavelength (λc)
4.3.3. UVA/UVB Ratio
4.3.4. Transmission of Erythema (%) and Transmission of Pigmentation (%)
4.4. Maintenance of Cell Lines
4.5. Determination of the Cytotoxic Potential of HepG2, and Calu-1 Cells in the MTT Viability Assay
4.6. ROS Detection Assay of EOs in HepG2 and Calu-1 Cells
4.7. Statistical Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Olivero-Verbel, J.; González-Cervera, T.; Güette-Fernandez, J.; Jaramillo-Colorado, B.; Stashenko, E.E. Chemical composition and antioxidant activity of essential oils isolated from Colombian plants. Rev. Bras. Farmacogn. 2010, 20, 568–574. [Google Scholar] [CrossRef] [Green Version]
- Stashenko, E.E.; Martínez, R.J. Study of essential oils obtained from tropical plants grown in Colombia. In Essential Oils-Oils of Nature; El-Shemy, H.A., Ed.; IntechOpen: London, UK, 2019; pp. 403–451. [Google Scholar]
- Kumar, S.; Pandey, A.K. Chemistry and biological activities of flavonoids: An overview. Sci. World J. 2013, 2013, 162750. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ngamsuk, S.; Huang, T.C.; Hsu, J.L. Determination of phenolic compounds, procyanidins, and antioxidant activity in processed Coffea arabica L. leaves. Foods 2019, 8, 389. [Google Scholar] [CrossRef] [Green Version]
- Fuentes, J.L.; Villamizar Mantilla, D.A.; Flores González, S.J.; Núñez, L.A.; Stashenko, E.E. Plants growing in Colombia as sources of active ingredients for sunscreens. Int. J. Radiat. Biol. 2021, 97, 1705–1715. [Google Scholar] [CrossRef]
- Dorni, A.C.; Amalraj, A.; Gopi, S.; Varma, K.; Anjana, S.N. Novel cosmeceuticals from plants—An industry guided review. J. Appl. Res. Med. Aromat. Plants 2017, 7, 1–26. [Google Scholar]
- Boveiri-Dehsheikh, A.; Mahmoodi Sourestani, M.; Boveiri Dehsheikh, P.; Vitalini, S.; Iriti, M.; Mottaghipisheh, J. A comparative study of essential oil constituents and phenolic compounds of Arabian Lilac (Vitex trifolia var. Purpurea): An evidence of season effects. Foods 2019, 8, 52. [Google Scholar] [CrossRef] [Green Version]
- Kostka, T.; Ostberg-Potthoff, J.J.; Briviba, K.; Matsugo, S.; Winterhalter, P.; Esatbeyoglu, T. Pomegranate (Punica granatum L.) extract and its anthocyanin and copigment fractions—Free radical scavenging activity and influence on cellular oxidative stress. Foods 2020, 9, 1617. [Google Scholar] [CrossRef]
- Angelini, P.; Tirillini, B.; Akhtar, M.S.; Dimitriu, L.; Bricchi, E.; Bertuzzi, G.; Venanzoni, R. Essential oil with anticancer activity: An overview. In Anticancer Plants: Natural Products and Biotechnological Implements; Akhtar, M., Swamy, M., Eds.; Springer: Singapore, 2018; pp. 207–231. [Google Scholar]
- Bayala, B.; Coulibaly, A.Y.; Djigma, F.W.; Nagalo, B.M.; Baron, S.; Figueredo, G.; Lobaccaro, J.; Simpore, J. Chemical composition, antioxidant, anti-inflammatory and antiproliferative activities of the essential oil of Cymbopogon nardus, a plant used in traditional medicine. Biomol. Concepts 2020, 11, 86–96. [Google Scholar] [CrossRef]
- Saikia, D.; Khanuja, S.P.; Kahol, A.P.; Gupta, S.C.; Kumar, S. Comparative antifungal activity of essential oils and constituents from three distinct genotypes of Cymbopogon spp. Curr. Sci. 2001, 80, 1264–1266. [Google Scholar]
- Sepulveda-Arias, J.C.; Veloza, L.A.; Escobar, L.M.; Orozco, L.M.; Lopera, I.A. Anti-inflammatory effects of the main constituents and epoxides derived from the essential oils obtained from Tagetes lucida, Cymbopogon citratus, Lippia alba and Eucalyptus citriodora. J. Essent. Oil Res. 2013, 25, 186–193. [Google Scholar] [CrossRef]
- Hernández-León, A.; González-Trujano, M.E.; Narváez-González, F.; Pérez-Ortega, G.; Rivero-Cruz, F.; Aguilar, M.I. Role of β-caryophyllene in the antinociceptive and anti-inflammatory effects of Tagetes lucida Cav. Essential oil. Molecules 2020, 25, 675. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gasparro, F.P.; Mitchnick, M.; Nash, J.F. A review of sunscreen safety and efficacy. Photochem. Photobiol. 1998, 68, 243–256. [Google Scholar] [CrossRef] [PubMed]
- Tampucci, S.; Burgalassi, S.; Chetoni, P.; Monti, D. Cutaneous permeation and penetration of sunscreens: Formulation strategies and in vitro methods. Cosmetics 2018, 5, 1. [Google Scholar] [CrossRef] [Green Version]
- Cheimonidi, C.; Samara, P.; Polychronopoulos, P.; Tsakiri, E.N.; Nikou, T.; Myrianthopoulos, V.; Sakallaropoulos, T.; Zoumpourlis, V.; Mikros, E.; Pappassideri, I.; et al. Selective cytotoxicity of the herbal substance acteoside against tumor cells and its mechanistic insights. Redox Biol. 2018, 16, 169–178. [Google Scholar] [CrossRef]
- Salucci, S.; Burattini, S.; Curzi, D.; Buontempo, F.; Martelli, A.M.; Zappia, G.; Falcieri, E.; Battistelli, M. Antioxidants in the prevention of UVB-induced keratynocyte apoptosis. J. Photochem. Photobiol. B Biol. 2014, 141, 1–9. [Google Scholar] [CrossRef]
- Kansanen, E.; Kuosmanen, S.M.; Leinonen, H.; Levonen, A.L. The Keap1-Nrf2 pathway: Mechanisms of Activation and Dysregulation in Cancer. Redox Biol. 2013, 1, 45–49. [Google Scholar] [CrossRef] [Green Version]
- Albuquerque, B.R.; Heleno, S.A.; Oliveira, M.B.P.; Barros, L.; Ferreira, I.C. Phenolic compounds: Current industrial applications, limitations, and future challenges. Food Funct. 2021, 12, 14–29. [Google Scholar] [CrossRef]
- Panyakaew, J.; Chalom, S.; Sookkhee, S.; Saiai, A.; Chandet, N.; Meepowpan, P.; Thavornyutikarn, P.; Mungkornasawakul, P. Kaempferia sp. extracts as UV protecting and antioxidant agents in sunscreen. J. Herbs Spices Med. Plants 2021, 27, 37–56. [Google Scholar] [CrossRef]
- Rosenthal, A.; Stoddard, M.; Chipps, L.; Herrmann, J. Skin cancer prevention: A review of current topical options complementary to sunscreens. J. Eur. Acad. Dermatol. Venereol. 2019, 33, 1261–1267. [Google Scholar] [CrossRef]
- Shanbhag, S.; Nayak, A.; Narayan, R.; Nayak, U.Y. Anti-aging and sunscreens: Paradigm shift in cosmetics. Adv. Pharm. Bull. 2019, 9, 348. [Google Scholar] [CrossRef] [Green Version]
- Marabini, L.; Melzi, G.; Lolli, F.; Dell’Agli, M.; Piazza, S.; Sangiovanni, E.; Marinovich, M. Effects of Vitis vinifera L. leaves extract on UV radiation damage in human keratinocytes (HaCaT). J. Photochem. Photobiol. B Biol. 2020, 204, 111810. [Google Scholar] [CrossRef] [PubMed]
- Luze, H.; Nischwitz, S.P.; Zalaudek, I.; Müllegger, R.; Kamolz, L.P. DNA repair enzymes in sunscreens and their impact on photoageing—A systematic review. Photodermatol. Photoimmunol. Photomed. 2020, 36, 424–432. [Google Scholar] [CrossRef] [PubMed]
- Adewinogo, S.O.; Sharma, R.; Africa, C.W.; Marnewick, J.L.; Hussein, A.A. Chemical composition and cosmeceutical potential of the essential oil of Oncosiphon suffruticosum (L.) Källersjö. Plants 2021, 10, 1315. [Google Scholar] [CrossRef] [PubMed]
- Panich, U.; Sittithumcharee, G.; Rathviboon, N.; Jirawatnotai, S. Ultraviolet radiation-induced skin aging: The role of DNA damage and oxidative stress in epidermal stem cell damage mediated skin aging. Stem Cells Int. 2016, 2016, 7370642. [Google Scholar] [CrossRef] [Green Version]
- Fuentes, J.L. Las plantas como fuente de compuestos fotoprotectores frente al daño en el ADN producido por la radiación ultravioleta. Rev. Acad. Colomb. Ci. Exact. 2019, 43, 550–562. [Google Scholar] [CrossRef]
- Ikehata, H.; Higashi, S.; Nakamura, S.; Daigaku, Y.; Furusawa, Y.; Kamei, Y.; Watanabe, M.; Yamamoto, K.; Hieda, K.; Munakata, N.; et al. Action spectrum analysis of UVR genotoxicity for skin: The border wavelengths between UVA and UVB can bring serious mutation loads to skin. J. Investig. Dermatol. 2013, 133, 1850–1856. [Google Scholar] [CrossRef] [Green Version]
- Lan, C.C.E.; Hung, Y.T.; Fang, A.H.; Ching-Shuang, W. Effects of irradiance on UVA-induced skin aging. J. Dermatol. Sci. 2019, 94, 220–228. [Google Scholar] [CrossRef] [Green Version]
- Lohani, A.; Mishra, A.K.; Verma, A. Cosmeceutical potential of geranium and calendula essential oil: Determination of antioxidant activity and in vitro sun protection factor. J. Cosmet. Dermatol. 2019, 18, 550–557. [Google Scholar] [CrossRef]
- Lohani, A.; Verma, A.; Hema, G.; Pathak, K. Topical delivery of geranium/calendula essential oil-entrapped ethanolic lipid vesicular cream to combat skin aging. Biomed Res. Int. 2021, 2021, 4593759. [Google Scholar] [CrossRef]
- Zhou, Y.; He, L.; Zhang, N.; Ma, L.; Yao, L. Photoprotective effect of Artemisia sieversiana Ehrhart essential oil against UVB-induced photoaging in Mice. Photochem. Photobiol. 2021. Advance online publication. [Google Scholar] [CrossRef]
- da Fonseca, E.Q.; da Silva, G.F.; Maquiné, L.V.; Matsdorff, Y.; Monteiro, R.S.G.; Aranha, E.S.P.; de Vasconcellos, M.; Albuquerque, P.M. Photoprotective, antioxidant, antimicrobial and cytotoxic activities of guarana (Paullinia cupana) seed extracts. Int. J. Phytocos. Nat. Ingred. 2020, 7, 10. [Google Scholar] [CrossRef]
- Caballero-Gallardo, K.; Rodriguez-Niño, D.; Fuentes-Lopez, K.; Stashenko, E.; Olivero-Verbel, J. Chemical composition and bioactivity of essential oils from Cymbopogon nardus L. and Rosmarinus officinalis L. against Ulomoides dermestoides (Fairmaire, 1893) (Coleoptera: Tenebrionidae). J. Essent. Oil-Bear. Plants 2021, 24, 547–560. [Google Scholar] [CrossRef]
- Baptista-Silva, S.; Borges, S.; Ramos, O.L.; Pintado, M.; Sarmento, B. The progress of essential oils as potential therapeutic agents: A review. J. Essent. Oil Res. 2020, 32, 279–295. [Google Scholar] [CrossRef]
- Alcala-Orozco, M.; Caballero-Gallardo, K.; Stashenko, E.E.; Olivero-Verbel, J. Repellent and fumigant actions of the essential oils from Elettaria cardamomum (L.) Maton, Salvia officinalis (L.) Linnaeus, and Lippia origanoides (V.) Kunth against Tribolium castaneum and Ulomoides dermestoides. J. Essent. Oil-Bear. Plants 2019, 22, 18–30. [Google Scholar] [CrossRef]
- Cervantes-Ceballos, L.; Caballero-Gallardo, K.; Olivero-Verbel, J. Repellent and anti-quorum sensing activity of six aromatic plants occurring in Colombia. Nat. Prod. Commun. 2015, 10, 1934578X1501001030. [Google Scholar] [CrossRef] [Green Version]
- Happy, A.A.; Jahan, F.; Momen, M.A. Essential oils: Magical ingredients for skin care. J. Plant Sci. 2021, 9, 54–64. [Google Scholar]
- Kaur, C.D.; Saraf, S. In vitro sun protection factor determination of herbal oils used in cosmetics. Pharmacogn. Res. 2010, 2, 22–25. [Google Scholar]
- Omoruyi, B.E.; Muchenje, V. Phytomedical assessment of two Cymbopogon species found in Nkonkobe Municipality: Toxicological effect on human Chang liver cell line. BMC Complement. Altern. Med. 2017, 17, 287. [Google Scholar] [CrossRef] [Green Version]
- Choudhary, S.; Zehra, A.; Mukarram, M.; Wani, K.I.; Naeem, M.; Khan, M.M.A.; Aftab, T. Salicylic acid-mediated alleviation of soil boron toxicity in Mentha arvensis and Cymbopogon flexuosus: Growth, antioxidant responses, essential oil contents and components. Chemosphere 2021, 276, 130153. [Google Scholar] [CrossRef]
- Devi, M.A.; Sahoo, D.; Singh, T.B.; Rajashekar, Y. Antifungal activity and volatile organic compounds analysis of essential oils from Cymbopogon species using solid-phase microextraction-gas chromatography-mass spectrometry. J. Agric. Food Res. 2021, 3, 100110. [Google Scholar] [CrossRef]
- Castillo, R.M.; Serrano, S.O.; Villamizar, A.L.R.; Sanchez, S.C.M.; Luna, J.E.D. Impact of Cymbopogon flexuosus L. (Poaceae) essential oil and primary components on the eclosion and larval development of Aedes aegypti (Diptera: Culicidae). BioRxiv. 2021, 11, 24291. [Google Scholar]
- Bhatnagar, A. Chemical composition and antioxidant activity of essential oil of Cymbopogon flexuosus. J. Appl. Nat. Sci. 2020, 12, 25–29. [Google Scholar] [CrossRef]
- Spencer, P.V.; Libardi, S.H.; Dias, F.F.; Oliveira, W.D.S.; Thomasini, R.L.; Godoy, H.T.; Cardoso, D.; Bogusz, S. Chemical composition, antioxidant and antibacterial activities of essential oil from Cymbopogon densiflorus (Steud.) Stapf flowers. J. Essent. Oil-Bear. Plants 2021, 24, 40–52. [Google Scholar] [CrossRef]
- Lal, M.; Baruah, J.; Begum, T.; Pandey, S.K. Identification of a novel myrcene and methyl iso-eugenol rich essential oil variant (Jor Lab L-11) of lemon grass (Cymbopogon flexuosus L.). J. Essent. Oil-Bear. Plants 2020, 23, 660–668. [Google Scholar] [CrossRef]
- Choudhary, S.; Zehra, A.; Naeem, M.; Khan, M.M.A.; Aftab, T. Effects of boron toxicity on growth, oxidative damage, antioxidant enzymes and essential oil fingerprinting in Mentha arvensis and Cymbopogon flexuosus. Chem. Biol. Technol. 2020, 7, 8. [Google Scholar] [CrossRef]
- Sharma, P.R.; Mondhe, D.M.; Muthiah, S.; Pal, H.C.; Shahi, A.K.; Saxena, A.K.; Qazi, G.N. Anticancer activity of an essential oil from Cymbopogon flexuosus. Chem. Biol. Interact. 2009, 179, 160–168. [Google Scholar] [CrossRef]
- Han, X.; Parker, T.L. Lemon grass (Cymbopogon flexuosus) essential oil demonstrated anti-inflammatory effect in pre-inflamed human dermal fibroblasts. Biochim. Open 2017, 4, 107–111. [Google Scholar] [CrossRef]
- Melo, A.M.D.; Turola Barbi, R.C.; Souza, W.F.C.D.; Luna, L.C.; de Souza, H.J.B.; Lucena, G.L.; Rocha, M.; de Sousa, S. Microencapsulated lemon grass (Cymbopogon flexuosus) essential oil: A new source of natural additive applied to Coalho cheese. J. Food Process. Preserv. 2020, 44, e14783. [Google Scholar] [CrossRef]
- López, M.A.; Stashenko, E.E.; Fuentes, J.L. Chemical composition and antigenotoxic properties of Lippia alba essential oils. Genet. Mol. Biol. 2011, 34, 479–488. [Google Scholar] [CrossRef] [Green Version]
- Monterrosas-Brisson, N.; Herrera-Ruiz, M.; Jiménez-Ferrer, E.J.F.; Bahena-Pérez, R.; Aviles-Flores, M.; Manasés, G.C. Anti-inflammatory activity of coumarins isolated from Tagetes lucida Cav. Nat. Prod. Res. 2020, 34, 3244–3248. [Google Scholar]
- El-Newary, S.A.; Ismail, R.F.; Shaffie, N.M.; Hendawy, S.F.; Omer, E.; Ahmed, M.M.; ELsayed, W.M. Hepatoprotective effects of Tagetes lucida root extract in carbon tetrachloride-induced hepatotoxicity in Wistar albino rats through amelioration of oxidative stress. Pharm. Biol. 2021, 59, 986–997. [Google Scholar] [CrossRef] [PubMed]
- Aquino, R.; Cáceres, A.; Morelli, S.; Rastrelli, L. An extract of Tagetes lucida and its phenolic constituents as antioxidants. J. Nat. Prod. 2002, 65, 1773–1776. [Google Scholar] [CrossRef] [PubMed]
- Monzote, L.; Gutiérrez, Y.; Machin, L.; Staniek, K.; Scull, R.; Satyal, P.; Gille, L.; Setzer, W.N. Antileishmanial activity and influence on mitochondria of the essential oil from Tagetes lucida Cav. and its main component. Sci. Pharm. 2020, 88, 31. [Google Scholar] [CrossRef]
- Ventura-Martínez, R.; Angeles-Lopez, G.E.; González-Trujano, M.E.; Carrasco, O.F.; Deciga-Campos, M. Study of antispasmodic and antidiarrheal activities of Tagetes lucida (Mexican Tarragon) in experimental models and its mechanism of action. Evid. Based Complement. Alternat. Med. 2020, 2020, 7140642. [Google Scholar] [CrossRef] [PubMed]
- Bergau, N.; Herfurth, U.M.; Sachse, B.; Abraham, K.; Monien, B.H. Bioactivation of estragole and anethole leads to common adducts in DNA and hemoglobin. Food Chem. Toxicol. 2021, 153, 112253. [Google Scholar] [CrossRef] [PubMed]
- Castillo, R.M.; Stashenko, E.; Duque, J.E. Insecticidal and repellent activity of several plant-derived essential oils against Aedes aegypti. J. Am. Mosq. Control Assoc. 2017, 33, 25–35. [Google Scholar] [CrossRef]
- Salehi, B.; Valussi, M.; Morais-Braga, M.F.B.; Carneiro, J.N.P.; Leal, A.L.A.B.; Coutinho, H.D.M.; Vitalini, S.; Kregiel, D.; Antolak, H.; Sharifi-Rad, M.; et al. Tagetes spp. essential oils and other extracts: Chemical characterization and biological activity. Molecules 2018, 23, 2847. [Google Scholar] [CrossRef] [Green Version]
- Radice, M.; Manfredini, S.; Ziosi, P.; Dissette, V.; Buso, P.; Fallacara, A.; Vertuani, S. Herbal extracts, lichens and biomolecules as natural photo-protection alternatives to synthetic UV filters. A systematic review. Fitoterapia 2016, 114, 144–162. [Google Scholar] [CrossRef]
- Rahardhian, M.R.R.; Suharsanti, R.; Sugihartini, N.; Lukitaningsih, E. In vitro assessment of total phenolic, total flavonoid and sunscreen activities of crude ethanolic extract of Belimbing wuluh (Averrhoa bilimbi) fruits and leaves. J. Glob. Pharma Technol. 2019, 11, 308–313. [Google Scholar]
- Karunarathne, W.A.H.M.; Molagoda, I.M.N.; Lee, K.T.; Choi, Y.H.; Yu, S.M.; Kang, C.H.; Kim, G.Y. Protective effect of anthocyanin-enriched polyphenols from Hibiscus syriacus L. (Malvaceae) against ultraviolet b-induced damage. Antioxidants 2021, 10, 584. [Google Scholar] [CrossRef]
- Lyons, A.B.; Trullas, C.; Kohli, I.; Hamzavi, I.H.; Lim, H.W. Photoprotection beyond ultraviolet radiation: A review of tinted sunscreens. J. Am. Acad. Dermatol. 2021, 84, 1393–1397. [Google Scholar] [CrossRef] [PubMed]
- Parrado, C.; Mercado-Saenz, S.; Perez-Davo, A.; Gilaberte, Y.; Gonzalez, S.; Juarranz, A. Environmental stressors on skin aging. mechanistic insights. Front. Pharmacol. 2019, 10, 759. [Google Scholar] [CrossRef] [PubMed]
- Liu, X.; Gao, Y.; Long, X.; Hayashi, T.; Mizuno, K.; Hattori, S.; Fujisaki, H.; Ogura, T.; Wang, D.O.; Ikejima, T. Type I collagen promotes the migration and myogenic differentiation of C2C12 myoblasts via the release of interleukin-6 mediated by FAK/NF-κB p65 activation. Food Funct. 2020, 11, 328–338. [Google Scholar] [CrossRef] [PubMed]
- Lionetti, N.; Rigano, L. The new sunscreens among formulation strategy, stability issues, changing norms, safety and efficacy evaluations. Cosmetics 2017, 4, 15. [Google Scholar] [CrossRef] [Green Version]
- Guan, L.L.; Lim, H.W.; Mohammad, T.F. Sunscreens and photoaging: A review of current literature. Am. J. Clin. Dermatol. 2021, 22, 819–828. [Google Scholar] [CrossRef] [PubMed]
- Mishra, A.K.; Mishra, A.; Chattopadhyay, P. Assessment of in vitro sun protection factor of Calendula officinalis L. (Asteraceae) essential oil formulation. J. Young Pharm. 2012, 4, 17–21. [Google Scholar] [CrossRef] [Green Version]
- Gaonkar, R.; Shiralgi, Y.; Lakkappa, D.B.; Hegde, G. Essential oil from Cymbopogon flexuosus as the potential inhibitor for HSP90. Toxicol. Rep. 2018, 5, 489–496. [Google Scholar] [CrossRef]
- Jakubczyk, K.; Dec, K.; Kałduńska, J.; Kawczuga, D.; Kochman, J.; Janda, K. Reactive oxygen species-sources, functions, oxidative damage. Pol. Merkur. Lekarski. 2020, 48, 124–127. [Google Scholar]
- Renaudin, X. Reactive oxygen species and DNA damage response in cancer. Int. Rev. Cell Mol. Biol. 2021, 364, 139–161. [Google Scholar]
- Wu, Z.; Wang, H.; Fang, S.; Xu, C. Roles of endoplasmic reticulum stress and autophagy on H2O2-induced oxidative stress injury in HepG2 cells. Mol. Med. Rep. 2018, 18, 4163–4174. [Google Scholar] [CrossRef] [Green Version]
- Xu, J.; Zhang, Y.; Ren, G.; Yang, R.; Chen, J.; Xiang, X.; Qin, J.; Chen, J. Inhibitory effect of delphinidin on oxidative stress induced by H2O2 in HepG2 cells. Oxid. Med. Cell. Longev. 2020, 2020, 1–12. [Google Scholar] [CrossRef] [PubMed]
- Nassar, M.; Zadri, F.; Slimani, S. Assessment of the protective effect of the methanolic extract from Salvia verbenaca roots against oxidative damage induced by hydrogen peroxide (H2O2). Turkish J. Pharm. Sci. 2020, 18, 360–366. [Google Scholar] [CrossRef] [PubMed]
- Lister, I.N.E.; Ginting, C.N.; Girsang, E.; Nataya, E.D.; Azizah, A.M.; Widowati, W. Hepatoprotective properties of red betel (Piper crocatum Ruiz and Pav) leaves extract towards H2O2-induced HepG2 cells via anti-inflammatory, antinecrotic, antioxidant potency. Saudi Pharm. J. 2020, 28, 1182–1189. [Google Scholar] [CrossRef]
- Elmann, A.; Mordechay, S.; Rindner, M.; Larkov, O.; Elkabetz, M.; Ravid, U. Protective effects of the essential oil of Salvia fruticosa and its constituents on astrocytic susceptibility to hydrogen peroxide-induced cell death. J. Agric. Food Chem. 2009, 57, 6636–6641. [Google Scholar] [CrossRef] [PubMed]
- Porres-Martínez, M.; González-Burgos, E.; Carretero, M.E.; Gómez-Serranillos, M.P. Protective properties of Salvia lavandulifolia Vahl. essential oil against oxidative stress-induced neuronal injury. Food Chem. Toxicol. 2015, 80, 154–162. [Google Scholar] [CrossRef]
- Kang, J.S.; Han, M.H.; Kim, G.Y.; Kim, C.M.; Chung, H.Y.; Hwang, H.J.; Kim, B.W.; Choi, Y.H. Schisandrae semen essential oil attenuates oxidative stress-induced cell damage in C2C12 murine skeletal muscle cells through Nrf2-mediated upregulation of HO-1. Int. J. Mol. Med. 2015, 35, 453–459. [Google Scholar] [CrossRef] [Green Version]
- Quintanilla, R.R.; Carlos, R.U.; Moyano, G.A.; Salazar, H.C.; Martínez, J.; Stashenko, E. Estudio comparativo de la composición de los aceites esenciales de cuatro especies del género Cymbopogon (Poaceae) cultivadas en Colombia. Bol. Latinoam. Caribe Plantas Med. Aromát. 2012, 11, 77–85. [Google Scholar]
- Ríos, N.; Stashenko, E.E.; Duque, J.E. Evaluation of the insecticidal activity of essential oils and their mixtures against Aedes aegypti (Diptera: Culicidae). Rev. Bras. Entomol. 2017, 61, 307–311. [Google Scholar] [CrossRef]
- Sánchez-Gutiérrez, J.A.; Moreno-Lorenzana, D.; Álvarez-Bernal, D.; Rodríguez-Campos, J.; Medina-Medrano, J.R. Phenolic profile, antioxidant, and anti-proliferative activities of methanolic extracts from Asclepias linaria Cav. leaves. Molecules 2020, 25, 54. [Google Scholar] [CrossRef] [Green Version]
- De Souza Mansur, J.; Breder, M.N.R.; dAscensão Mansur, M.C.; Azulay, R.D. Determination of sun protecting factor in human beings and by spectrophotometry: Comparison between of the two methods. An. Bras. Dermatol. 1986, 61, 167–172. [Google Scholar]
- Sayre, R.M.; Agin, P.P.; LeVee, G.J.; Marlowe, E. A comparison of in vivo and in vitro testing of sun screening formulas. Photochem. Photobiol. 1979, 29, 559–566. [Google Scholar] [CrossRef] [PubMed]
- Springsteen, A.; Yurek, R.; Frazier, M.; Carr, K.F. In vitro measurement of sun protection factor of sunscreens by diffuse transmittance. Anal. Chim. Acta 1999, 380, 155–164. [Google Scholar] [CrossRef]
- Pelizzo, M.; Zattra, E.; Nicolosi, P.; Peserico, A.; Garoli, D.; Alaibac, M. In vitro evaluation of sunscreens: An update for the clinicians. Int. Sch. Res. Not. 2012, 2012, 352135. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Boots the Chemist Ltd. The Revised Guidelines to the Practical Measurement of UVA/UVB Ratios According to the Boots Star Rating System; The Boots Company PLC: Nottingham, UK, 2004. [Google Scholar]
- Donglikar, M.M.; Deore, S.L. Sunscreens: A review. Pharmacogn. Mag. 2016, 8, 171–179. [Google Scholar] [CrossRef] [Green Version]
- Sami, F.J.; Soekamto, N.H.; Latip, J. Bioactivity profile of three of seaweed as an antioxidant, UV-protection as sunscreen and their correlation activity. Food Res. 2021, 5, 441–447. [Google Scholar] [CrossRef]
- Caballero-Gallardo, K.; Guerrero-Castilla, A.; Johnson-Restrepo, B.; de la Rosa, J.; Olivero-Verbel, J. Chemical and toxicological characterization of sediments along a Colombian shoreline impacted by coal export terminals. Chemosphere 2015, 138, 837–846. [Google Scholar] [CrossRef]
- Alvarez-Ortega, N.; Caballero-Gallardo, K.; Taboada-Alquerque, M.; Franco, J.; Stashenko, E.E.; Juan, C.; Juan-García, A.; Olivero-Verbel, J. Protective effects of the hydroethanolic extract of Fridericia chica on undifferentiated human neuroblastoma cells exposed to α-Zearalenol (α-ZEL) and β-Zearalenol (β-ZEL). Toxins 2021, 13, 748. [Google Scholar] [CrossRef]
- Caballero-Gallardo, K.; Olivero-Verbel, J.; Corada-Fernández, C.; Lara-Martín, P.A.; Juan-García, A. Emerging contaminants and priority substances in marine sediments from Cartagena Bay and the Grand Marsh of Santa Marta (Ramsar site), Colombia. Environ. Monit. Assess. 2021, 193, 596. [Google Scholar] [CrossRef]
- Barzegar, A.; Moosavi-Movahedi, A.A. Intracellular ROS protection efficiency and free radical-scavenging activity of curcumin. PLoS ONE 2011, 6, e26012. [Google Scholar] [CrossRef] [Green Version]
- Agahi, F.; Álvarez-Ortega, N.; Font, G.; Juan-García, A.; Juan, C. Oxidative stress, glutathione, and gene expression as key indicators in SH-SY5Y cells exposed to zearalenone metabolites and beauvericin. Toxicol. Lett. 2020, 334, 44–52. [Google Scholar] [CrossRef]
- Barnes, P.W.; Williamson, C.E.; Lucas, R.M.; Robinson, S.A.; Madronich, S.; Paul, N.D.; Bornman, J.F.; Bais, A.F.; Sulzberger, B.; Wilson, S.R.; et al. Ozone depletion, ultraviolet radiation, climate change and prospects for a sustainable future. Nat. Sustain. 2019, 2, 569–579. [Google Scholar] [CrossRef] [Green Version]




{kind=link}
{kind=link}
{kind=link}
| Essential Oil | Mean ± SEM |
|---|---|
| Phenolic Content (mg GAE/g of ByProduct) | |
| Cananga odorata | 24.4 ± 0.4 |
| Citrus sinensis | 13.8 ± 0.9 |
| Cymbopogon citratus | 26.6 ± 0.3 |
| C. flexuosus | 10.6 ± 0.1 |
| C. martinii | 13.6 ± 0.1 |
| C. nardus | 18.6 ± 0.2 |
| Elettaria cardamomum | 12.7 ± 0.1 |
| Lippia alba | 16.4 ± 0.1 |
| L. origanoides (Phellandrene) | 21.9 ± 0.1 |
| L. origanoides (Carvacrol/thymol) | 26.9 ± 0.1 |
| Tagetes lucida | 12.7 ± 0.1 |
| Thymus vulgaris | 19.0 ± 0.1 |
| Essential Oils | In vitro Measurements of Sunscreen Protection (Mean ± SEM), n = 3 | ||||
|---|---|---|---|---|---|
| SPF a | λc | UVA/UVB Ratio | Transmission of Erythema (%) | Transmission of Pigmentation (%) | |
| C. odorata | 8.7 ± 0.0 | ND | 0.4 ± 0.0 | 4.4 ± 0.0 | 43.3 ± 0.2 |
| C. sinensis | 3.2 ± 0.0 | ND | 1.1 ± 0.1 | 17.0 ± 0.1 | 71.9 ± 0.4 |
| C. citratus | 10.0 ± 0.2 | 360.9 ± 1.8 | 1.2 ± 0.0 | 3.5 ± 0.2 | 23.5 ± 0.7 |
| C. flexuosus | 13.4 ± 0.3 | 391.1 ± 1.9 | 0.7 ± 0.0 | 1.4 ± 0.1 | 20.3 ± 0.6 |
| C. martini | 5.2 ± 0.0 | ND | 0.6 ± 0.0 | 10.0 ± 0.1 | 55.9 ± 0.2 |
| C. nardus | 4.5 ± 0.0 | ND | 0.7 ± 0.0 | 12.0 ± 0.2 | 59.8 ± 0.3 |
| E. cardamomum | 4.7 ± 0.0 | ND | 0.7 ± 0.0 | 11.3 ± 0.2 | 57.5 ± 0.5 |
| L. alba | 9.6 ± 0.1 | 346.7 ± 1.5 | 1.0 ± 0.0 | 3.4 ± 0.0 | 29.8 ± 0.0 |
| L. origanoides (Phellandrene) | 3.6 ± 0.0 | ND | 0.3 ± 0.0 | 15.5 ± 0.4 | 70.4 ± 0.9 |
| L. origanoides (Carvacrol/thymol) | 11.7 ± 0.2 | 351.4 ± 0.0 | 0.1 ± 0.0 | 1.9 ± 0.0 | 63.9 ± 0.9 |
| T. lucida | 14.7 ± 0.0 | 393.5 ± 0.2 | 0.7 ± 0.0 | 1.2 ± 0.0 | 12.5 ± 0.1 |
| T. vulgaris | 4.8 ± 0.0 | ND | 0.5 ± 0.0 | 11.1 ± 0.0 | 58.1 ± 0.1 |
| Trolox b | 3.6 ± 0.0 | 371.6 ± 0.5 | 0.1 ± 0.0 | 38.3 ± 0.1 | 71.3 ± 0.2 |
| Vanillin b | 56.4 ± 0.9 | 375.1 ± 0.0 | 0.6 ± 0.0 | 0.0 ± 0.0 | 5.5 ± 0.1 |
| Gallic acid b | 34.7 ± 0.1 | 348.2 ± 0.4 | 0.0 ± 0.0 | 0.0 ± 0.0 | 45.3 ± 0.5 |
| Body lotion c | 36.3 ± 0.1 | 392.6 ± 0.3 | 0.4 ± 0.1 | 0.02 ± 0.2 | 11.7 ± 0.1 |
| Sun protector c | 54.3 ± 0.2 | 393.1 ± 0.1 | 2.7 ± 0.2 | 0.0 ± 0.0 | 0.0 ± 0.0 |
| Essential Oil | Cytotoxic Potential Derived from MTT Viability Assay (24 h) | |||||
|---|---|---|---|---|---|---|
| HepG2 | Calu-1 | |||||
| IC50 | R2 | p-Value | IC50 | R2 | p-Value | |
| Cymbopogon flexuosus | 75 | 0.986 | <0.001 | 100 | 0.865 | 0.007 |
| Tagetes lucida | 270 | 0.912 | <0.001 | 381 | 0.856 | 0.001 |
| Plant | Family | Major Constituents (%) | Reference |
|---|---|---|---|
| C. odorata | Annonaceae | Ethyl benzoate (18.2), linalool (14.0), benzyl benzoate (12.3), methyl benzoate (10.0) | [43] |
| C. sinensis | Rutaceae | Limonene (71.3) | [43] |
| C. citratus | Poaceae | Geranial (34.4), neral (28.4), geraniol (11.5) | [43] |
| C. flexuosus | Poaceae | Neral (28.2), geranial (28.2), geranyl acetate (10.0) | [43] |
| C. martinii | Poaceae | Geraniol (83.9) | [80] |
| C. nardus | Poaceae | Citronellal (25.3), citronellol (17.9), geraniol (11.6) | [34] |
| E. cardamomum | Zingiberaceae | 1,8-Cineole (30.9), terpinyl acetate (26.4) | [36] |
| L. alba | Verbenaceae | Carvone (38.3), limonene (31.8), bicyclo- sesquiphellandrene (11.4) | [43] |
| L. origanoides (Phellandrene chemotype) | Verbenaceae | Limonene (15.0), p-cymene (14.6), α-phellandrene (10.3) | [80] |
| L. origanoides (Carvacrol/thymol chemotype) | Verbenaceae | Carvacrol (50.6), thymol (11.5) | [43] |
| T. lucida | Asteraceae | Estragole (95.7) | [43] |
| T. vulgaris | Lamiaceae | Thymol (42.0), p-cymene (26.4) | [80] |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Caballero-Gallardo, K.; Quintero-Rincón, P.; Stashenko, E.E.; Olivero-Verbel, J. Photoprotective Agents Obtained from Aromatic Plants Grown in Colombia: Total Phenolic Content, Antioxidant Activity, and Assessment of Cytotoxic Potential in Cancer Cell Lines of Cymbopogon flexuosus L. and Tagetes lucida Cav. Essential Oils. Plants 2022, 11, 1693. https://0-doi-org.brum.beds.ac.uk/10.3390/plants11131693
Caballero-Gallardo K, Quintero-Rincón P, Stashenko EE, Olivero-Verbel J. Photoprotective Agents Obtained from Aromatic Plants Grown in Colombia: Total Phenolic Content, Antioxidant Activity, and Assessment of Cytotoxic Potential in Cancer Cell Lines of Cymbopogon flexuosus L. and Tagetes lucida Cav. Essential Oils. Plants. 2022; 11(13):1693. https://0-doi-org.brum.beds.ac.uk/10.3390/plants11131693
Chicago/Turabian StyleCaballero-Gallardo, Karina, Patricia Quintero-Rincón, Elena E. Stashenko, and Jesus Olivero-Verbel. 2022. "Photoprotective Agents Obtained from Aromatic Plants Grown in Colombia: Total Phenolic Content, Antioxidant Activity, and Assessment of Cytotoxic Potential in Cancer Cell Lines of Cymbopogon flexuosus L. and Tagetes lucida Cav. Essential Oils" Plants 11, no. 13: 1693. https://0-doi-org.brum.beds.ac.uk/10.3390/plants11131693