
Effects of Different Drought Degrees on Physiological Characteristics and Endogenous Hormones of Soybean
 and
and {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
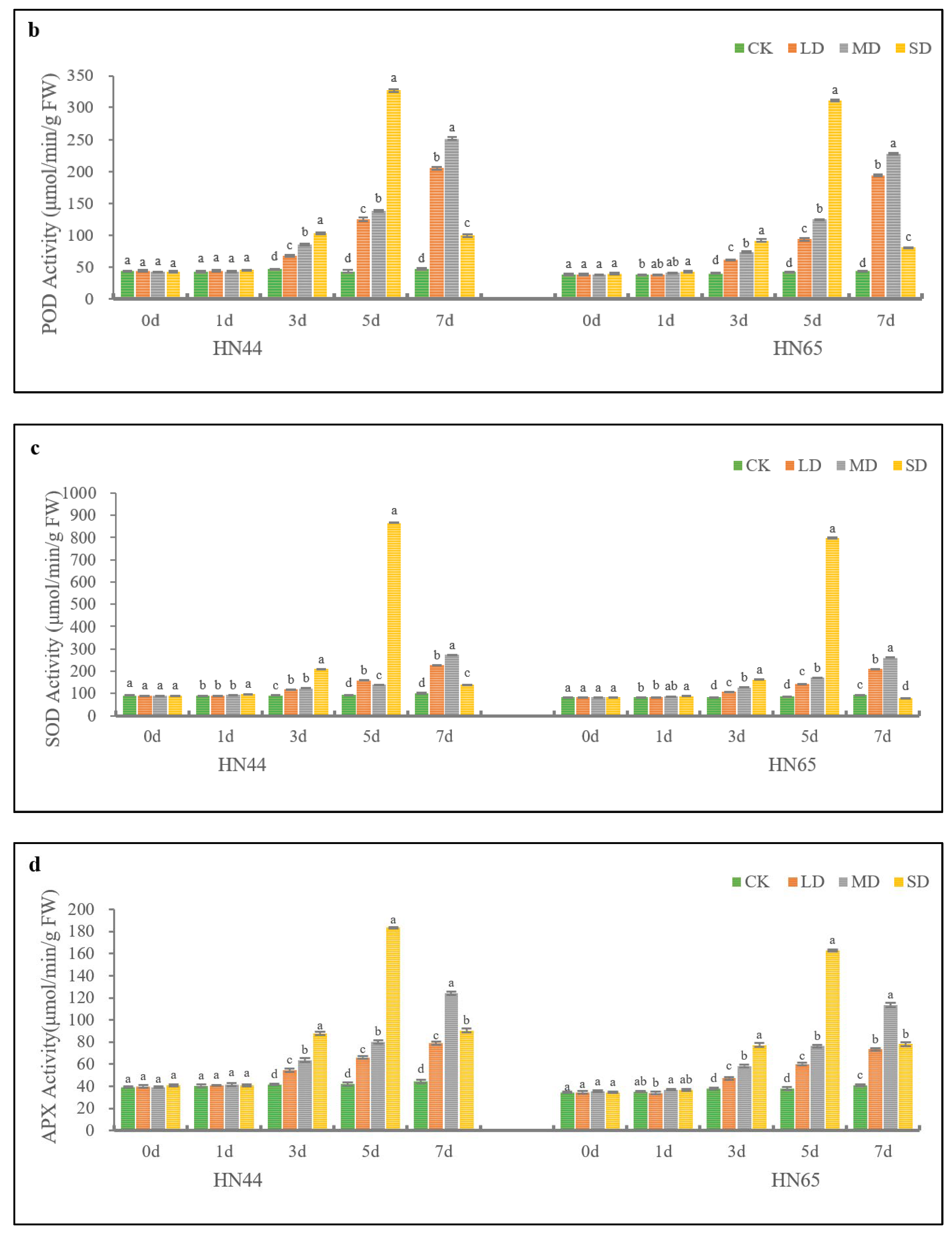
2.1. Effects of Drought Stress on Antioxidant Enzymes of Soybean
2.2. Effects of Drought Stress on Antioxidant Enzymes of Soybean
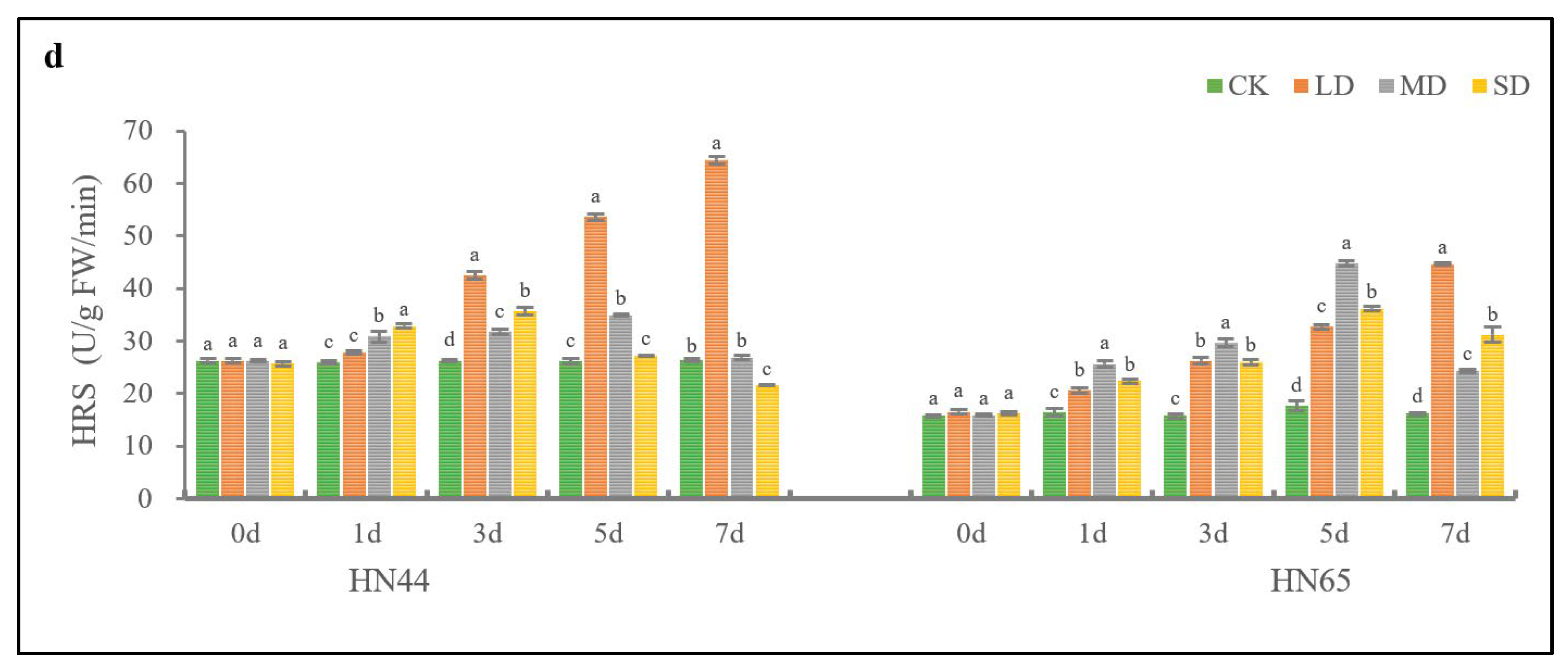
2.3. Effects of Drought Stress on Osmotic Regulators of Soybean
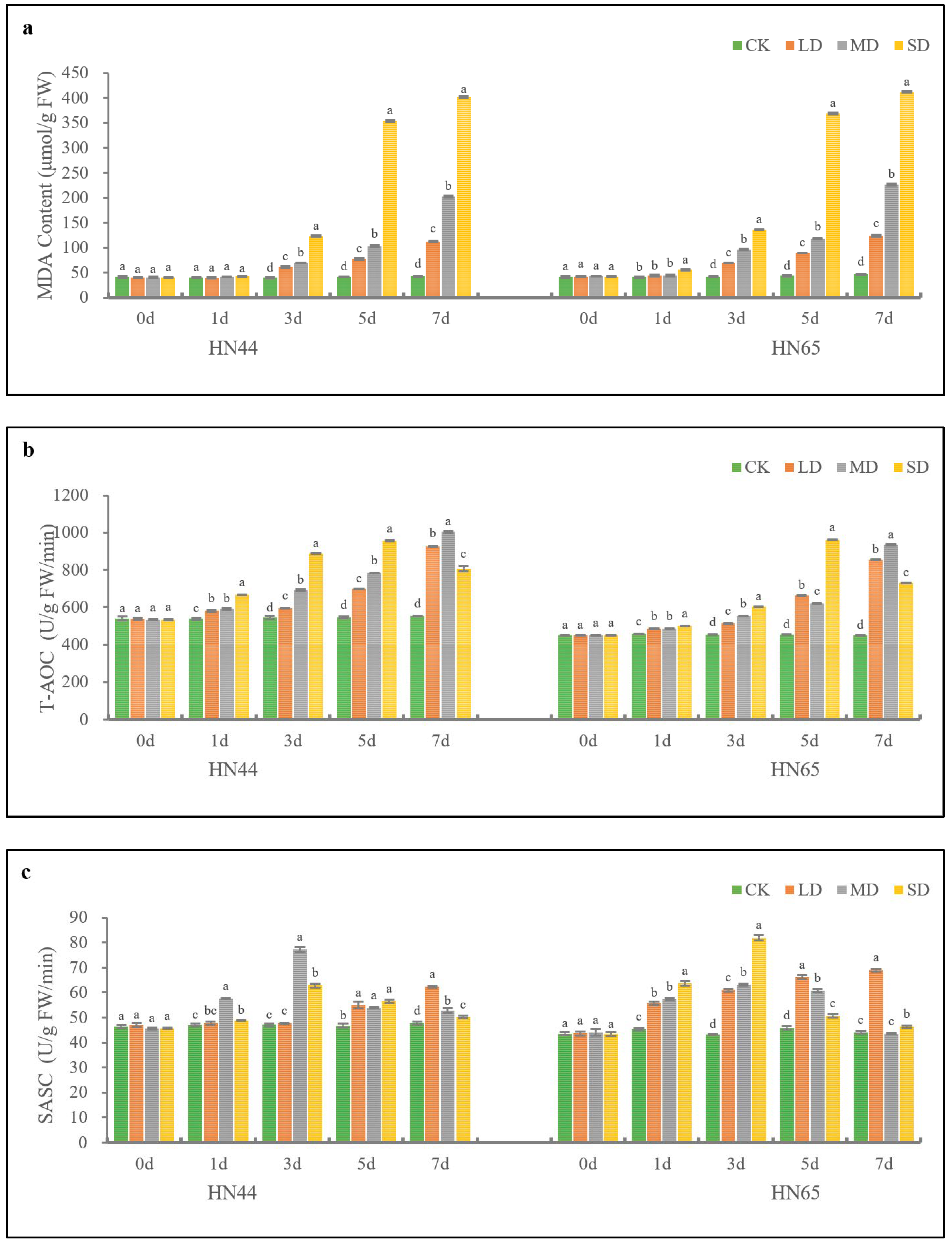
2.4. Effects of Drought Stress on MDA and Antioxidant Capacity of Soybean
2.5. Effect of Drought Stress on Endogenous Hormone Content of Soybean
2.6. Principal Component Analysis
2.7. Correlation Plot
3. Discussion
4. Materials and Methods
4.1. Testing Material
4.2. Experimental Design
4.3. Determination of Physiological and Biochemical Indexes
4.3.1. Determination of Relative Moisture Content of Leaves
4.3.2. Determination of Antioxidant Enzyme Activity
4.3.3. Determination of Osmotic Adjustment Substance Content
4.3.4. Determination of MDA Content and Antioxidant Capacity
4.3.5. Determination of Endogenous Hormone Content
4.3.6. Data Analysis
5. Conclusions
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
References
- Eric, S.J.; Wu, F.; Yoshie, H. Soybean domestication: The origin, genetic architecture and molecular bases. New Phytol. 2017, 214, 539–553. [Google Scholar]
- Costa, J.H.; Roque, A.L.M.; Aziz, S.; Dos, S.C.P.; Germano, T.A.; Batista, M.C.; Thiers, K.L.L. Genome-wide identification of ascorbate-glutathione cycle gene families in soybean (Glycine max) reveals gene duplication events and specificity of gene members linked to development and stress conditions. Int. J. Biol. Macromol. 2021, 187, 528–543. [Google Scholar] [CrossRef] [PubMed]
- Food and Agriculture Organization. Soybeans. 2021. Available online: https://www.fao.org/home/en/ (accessed on 1 August 2022).
- Bhusal, N.; Lee, M.; Han, A.R.; Han, A.; Kim, H.S. Responses to drought stress in Prunus sargentii and Larix kaempferi seedlings using morphological and physiological parameters. For. Ecol. Manag. 2020, 465, 118099. [Google Scholar] [CrossRef]
- Das, K.; Roychoudhury, A. Reactive oxygen species (ROS) and response of antioxidants as ROS-scavengers during environmental stress in plants. Front. Environ. Sci. 2014, 2, 53. [Google Scholar]
- Li, B.Z.; Fan, R.N.; Sun, G.L.; Sun, T.; Fan, Y.T.; Bai, S.G.; Guo, S.Y.; Huang, S.Q.; Liu, J.; Zhang, H.; et al. Flavonoids improve drought tolerance of maize seedlings by regulating the homeostasis of reactive oxygen species. Plant Soil 2021, 461, 389–405. [Google Scholar] [CrossRef]
- Gill, S.S.; Tuteja, N. Reactive oxygen species and antioxidant machinery in abiotic stress tolerance in crop plants. Plant Physiol. Biochem. 2010, 48, 909–930. [Google Scholar] [CrossRef]
- Liu, B.H.; Jing, D.W.; Liu, F.C.; Ma, H.L.; Liu, X.H.; Peng, L. Serendipita indica alleviates drought stress responses in walnut (Juglans regia L.) seedlings by stimulating osmotic adjustment and antioxidant defense system. Appl. Microbiol. Biotechnol. 2021, 105, 8951–8968. [Google Scholar] [CrossRef]
- Cao, L.; Jin, X.J.; Zhang, Y.X. Melatonin confers drought stress tolerance in soybean (Glycine max L.) by modulating photosynthesis, osmolytes, and reactive oxygen metabolism. Photosynthetica 2019, 3, 812–819. [Google Scholar] [CrossRef]
- Kaur, G.; Asthir, B. Proline: A key player in plant abiotic stress tolerance. Biol. Plant 2015, 59, 609–619. [Google Scholar] [CrossRef]
- Saddique, M.A.B.; Zulfiqar, A.; Sher, M.A.; Farid, B.; Ikram, R.M.; Ahmad, M.S. Proline, Total Antioxidant Capacity, and Gene Activity in Radical and Plumule of Rice are Efficient Drought Tolerance Indicator Traits. Int. J. Agron. 2020, 2020, 8862792. [Google Scholar] [CrossRef]
- Wang, X.Y.; Wu, Z.H.; Zhou, Q.; Wang, X.; Song, S.; Dong, S.K. Physiological Response of Soybean Plants to Water Deficit. Front. Recent Dev. Plant Sci. 2021, 12, 809692. [Google Scholar] [CrossRef]
- Pirasteh, A.H.; Emam, Y.; Pessarakli, M. Changes in Endogenous Hormonal Status in Corn (Zea mays) Hybrids Under Drought Stress. J. Plant Nutr. 2013, 36, 1695–1707. [Google Scholar] [CrossRef]
- Muhammad, A.; Shao, Y.H.; Liu, S.X.; Wang, F.; Gao, J.W.; Jiang, D.; Tian, Z.W.; Dai, T.B. Pre-drought priming sustains grain development under post-anthesis drought stress by regulating the growth hormones in winter wheat (Triticum aestivum L.). Planta 2017, 246, 509–524. [Google Scholar]
- Soba, D.; Zhou, B.W.; Arrese-lgor, C.; Munne-Bosch, S.; Aranjuelo, I. Physiological, Hormonal and Metabolic Responses of two Alfalfa Cultivars with Contrasting Responses to Drought. Int. J. Mol. Sci. 2019, 20, 5099. [Google Scholar] [CrossRef]
- Elizabeth, A.M.; Sergi, M.B. Photo- and antioxidant protection and salicylic acid accumulation during post-anthesis leaf senescence in Salvia lanigera grown under Mediterranean climate. Physiol. Plant. 2007, 131, 590–598. [Google Scholar]
- Nir, I.; Moshelion, M.; Weiss, D. The Arabidopsis gibberellin methyl transferase 1 suppresses gibberellin activity, reduces whole-plant transpiration and promotes drought tolerance in transgenic tomato. Plant Cell Environ. 2014, 37, 113–123. [Google Scholar] [CrossRef]
- Hellal, F.A.; Shabrawi, H.M.; Abd El-Hady, M.; Khatab, I.A.; El-Sayed, S.A.A.; Chedly, A. Influence of PEG induced drought stress on molecular and biochemical constituents and seedling growth of Egyptian barley cultivars. J. Genet. Eng. Biotechnol. 2018, 16, 203–212. [Google Scholar] [CrossRef]
- Ahmend, M.; Kheir, A.M.S.; Mehmood, M.Z.; Ahmad, S.; Hasanuzzaman, M. Changes in Germination and Seedling Traits of Sesame under Simulated Drought. Phyton 2022, 91, 713–726. [Google Scholar]
- Badr, A.; El-Shazly, H.; Tarawneh, R.A.; Borner, A. Screening for Drought Tolerance in Maize (Zea mays L.) Germplasm Using Germination and Seedling Traits under Simulated Drought Conditions. Plants. 2022, 9, 565. [Google Scholar] [CrossRef]
- Anjum, S.A.; Ashraf, U.; Tanveer, M.; Khan, I.; Hussain, S.; Shahzad, B.; Zohaib, A.; Abbas, F.; Saleem, M.F.; Ail, I.; et al. Drought Induced Changes in Growth, Osmolyte Accumulation and Antioxidant Metabolism of Three Maize Hybrids. Front. Plant Sci. 2017, 8, 69. [Google Scholar] [CrossRef]
- Chen, W.Q.; Miao, Y.L.; Ayyaz, A.; Hannan, F.; Huang, Q.; Ulhassan, Z.; Zhou, Y.Y.; Islam, F.; Hong, Z.Y.; Farooq, M.A.; et al. Purple stem Brassica napus exhibits higher photosynthetic efficiency, antioxidant potential and anthocyanin biosynthesis related genes expression against drought stress. Front. Plant Sci. 2022, 13, 936696. [Google Scholar] [CrossRef] [PubMed]
- Alireza, K.; Rohangiz, N.; Cecilia, B.; Elena, M.B.; Alireza, S.S.; Mesbah, B. Morphological, physiochemical and antioxidant responses of Maclura pomifera to drought stress. Sci. Rep. 2019, 9, 19250. [Google Scholar]
- Rima, I.A.; Mannan, M.A.; Mamun, M.A.A.; Kamal, Z.U. Morpho-Physiological Traits of Soybean as Affected by Drought. Bangladesh Agron. J. 2020, 22, 41–54. [Google Scholar] [CrossRef]
- Bhusal, N.; Lee, M.; Lee, H.; Adhikari, A.; Han, R.; Han, A.; Kim, H.S. Evaluation of morphological, physiological, and biochemical traits for assessing drought resistance in eleven tree species. Sci. Total Environ. 2021, 779, 146466. [Google Scholar] [CrossRef]
- Mona, F.A.D.; Abbu, Z.; Arafat, A.H.A.L. Salicylic Acid Spraying-Induced Resilience Strategies Against the Damaging Impacts of Drought and/or Salinity Stress in Two Varieties of Vicia faba L. Seedlings. J. Plant Growth Regul. 2022, 41, 1919–1942. [Google Scholar]
- Balfagon, D.; Teran, F.; Oliveira, T.R.; Santa-Catarina, C.; Gomez-Cadenas, A. Citrus rootstocks modify scion antioxidant system under drought and heat stress combination. Plant Cell Rep. 2021, 41, 593–602. [Google Scholar] [CrossRef]
- Munir, O.; Bengu, T.U.; Pedro, G.C.; Anum, K.; Alvina, G.; Mirza, H. Osmoregulation and its actions during the drought stress in plants. Physiol. Plant. 2020, 172, 1321–1335. [Google Scholar]
- Li, B.Y.; Feng, Y.N.; Zong, Y.Z.; Zhang, D.S.; Hao, X.Y.; Li, P. Elevated CO 2 -induced changes in photosynthesis, antioxidant enzymes and signal transduction enzyme of soybean under drought stress. Plant Physiol. Biochem. 2020, 154, 105–114. [Google Scholar] [CrossRef]
- Guellim, A.; Hirel, B.; Chabrerie, O.; Catterou, M.; Tetu, T.; Dubois, F.; Ahmed, H.B.; Kichey, T. Screening for durum wheat (Triticum durum Desf.) cultivar resistance to drought stress using an integrated physiological approach. J. Crop Sci. Biotechnol. 2020, 23, 355–365. [Google Scholar] [CrossRef]
- Rui, H.Y.; Wang, L.N.; Jin, L.; Li, L.; Pan, H.; Li, J.Y.; Li, Z.Y.; Gu, W.; Jin, X. Effects of Drought Stress at Seedling Stage on Protective Enzyme Activity and Malondialdehyde Content in Leaves of Different Soybean Varieties. Soybean Sci. 2013, 32, 647–649. [Google Scholar]
- Ren, J.Y.; Wang, J.; Ai, X.; Zhao, S.L.; Li, R.Y.; Jiang, C.J.; Zhao, X.H.; Yin, D.M.; Yu, H.Q. Analysis of Drought Tolerance Physiological Response Characteristics of Peanut Seedling under Drought Stress. Chin. J. Oil Crop Sci. 2022, 44, 138–146. [Google Scholar]
- Tan, X.R.; Wu, X.Q.; Du, S.W.; Kong, J.; Dai, Y. Effect of Drought Stress on Active Oxygen Scavenging Ability of Wheat Seedlings. J. Anhui Agric. Sci. 2006, 15, 3565–3567. [Google Scholar]
- Sah, S.K.; Reddy, K.R.; Li, J. Abscisic Acid and Abiotic Stress Tolerance in Crop Plants. Front. Plant Sci. 2016, 7, 571. [Google Scholar] [CrossRef]
- Bhusal, N.; Han, S.G.; Yoon, T.M. Impact of drought stress on photosynthetic response, leaf water potential, and stem sap flow in two cultivars of bi-leader apple trees (Malus × domestica Borkh). Sci. Hortic. 2019, 246, 535–543. [Google Scholar] [CrossRef]
- Zhou, Y.F.; Wang, D.Q.; Lu, Z.S.; Wang, N.; Wang, Y.T.; Li, F.X.; Xu, J.; Huang, R.D. Effects of drought stress on photosynthetic characteristics and endogenous hormones ABA and CTK content of stay-green sorghum. Chin. Agric. Sci. 2014, 47, 655–663. [Google Scholar]
- Shi, H.T.; Chen, L.; Ye, T.T.; Liu, X.D.; Ding, K.J.; Chan, Z.L. Modulation of auxin content in Arabidopsis confers improved drought stress resistance. Plant Physiol. Biochem. 2014, 82, 209–217. [Google Scholar] [CrossRef]
- Wang, L.B.; Liu, J.L.; Pei, Y.F.; Dong, S.K.; Sun, C.S.; Zu, W.; Ruan, Y.H. Drought Resistance Identification of Soybean Germplasm Resources at Germination Stage. J. Northeast Agric. Univ. 2012, 43, 36–43. [Google Scholar]
- Wang, X.; Li, X.M.; Dong, S.K. Screening and identification of drought tolerance of spring soybean at seedling stage under climate change. Front. Sustain. Food Syst. 2022, 6, 988319. [Google Scholar] [CrossRef]
- Waheed, H.; Javaid, M.M.; Shahid, A.; Ali, H.H.; Nargis, J.; Mehmood, A. Impact of foliar-applied Hoagland’s nutrient solution on growth and yield of mash bean (Vigna mungo L.) under different growth stages. J. Plant Nutr. 2019, 42, 1133–1141. [Google Scholar] [CrossRef]
- Badr, A.; Bruggemann, W. Special issue in honour of Prof. Reto J. Strasser—Comparative analysis of drought stress response of maize genotypes using chlorophyll fluorescence measurements and leaf relative water content. Photosynthetica 2020, 58, 638–645. [Google Scholar] [CrossRef]
- Wang, X.K.; Huang, J.L. Principles and Techniques of Plant Physiological Biochemical Experiment; Higher Education Press: Beijing, China, 2015; pp. 280–288. [Google Scholar]
- Zhang, Z.A.; Zhang, M.S.; Wei, H.R. Experimental Guidance of Plant Physiology; China Agricultural Science and Technology Press: Beijing, China, 2004; pp. 138–140. [Google Scholar]
- Alica, B.; Alzbeta, D.; Lubomir, B.; Lubos, H.; Marek, B.; Lukas, J.; Katatina, P.; Monika, B.; Matus, B.; Julius, Á. Detection of Changes in Total Antioxidant Capacity, the Content of Polyphenols, Caffeine, and Heavy Metals of Teas in Relation to Their Origin and Fermentation. Foods 2021, 10, 1821. [Google Scholar]
- Wu, C.Q.; Chen, G.H. Detection of synergistic effect of superoxide dismutase and jujubosides on scavenging superoxide anion radical by capillary electrophoresis. Biomed. Chromatogr. 2019, 33, e4630. [Google Scholar] [CrossRef] [PubMed]
- Zhuang, Y.; Yan, P.S. Study on Hydroxyl Radical Scavenging Ability of Fermented Kelp Waste. Chem. Eng. Trans. 2016, 51, 289–294. [Google Scholar]










Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zhou, Q.; Li, Y.; Wang, X.; Yan, C.; Ma, C.; Liu, J.; Dong, S. Effects of Different Drought Degrees on Physiological Characteristics and Endogenous Hormones of Soybean. Plants 2022, 11, 2282. https://0-doi-org.brum.beds.ac.uk/10.3390/plants11172282
Zhou Q, Li Y, Wang X, Yan C, Ma C, Liu J, Dong S. Effects of Different Drought Degrees on Physiological Characteristics and Endogenous Hormones of Soybean. Plants. 2022; 11(17):2282. https://0-doi-org.brum.beds.ac.uk/10.3390/plants11172282
Chicago/Turabian StyleZhou, Qi, Yongping Li, Xiaojing Wang, Chao Yan, Chunmei Ma, Jun Liu, and Shoukun Dong. 2022. "Effects of Different Drought Degrees on Physiological Characteristics and Endogenous Hormones of Soybean" Plants 11, no. 17: 2282. https://0-doi-org.brum.beds.ac.uk/10.3390/plants11172282