
Relating Trait Variation to Species and Community Productivity in Contrasting Oro-Mediterranean Pastures: A 7-Years Study in the Pollino National Park (S-Italy)

 , ,
, ,
Abstract
:1. Introduction
2. Results
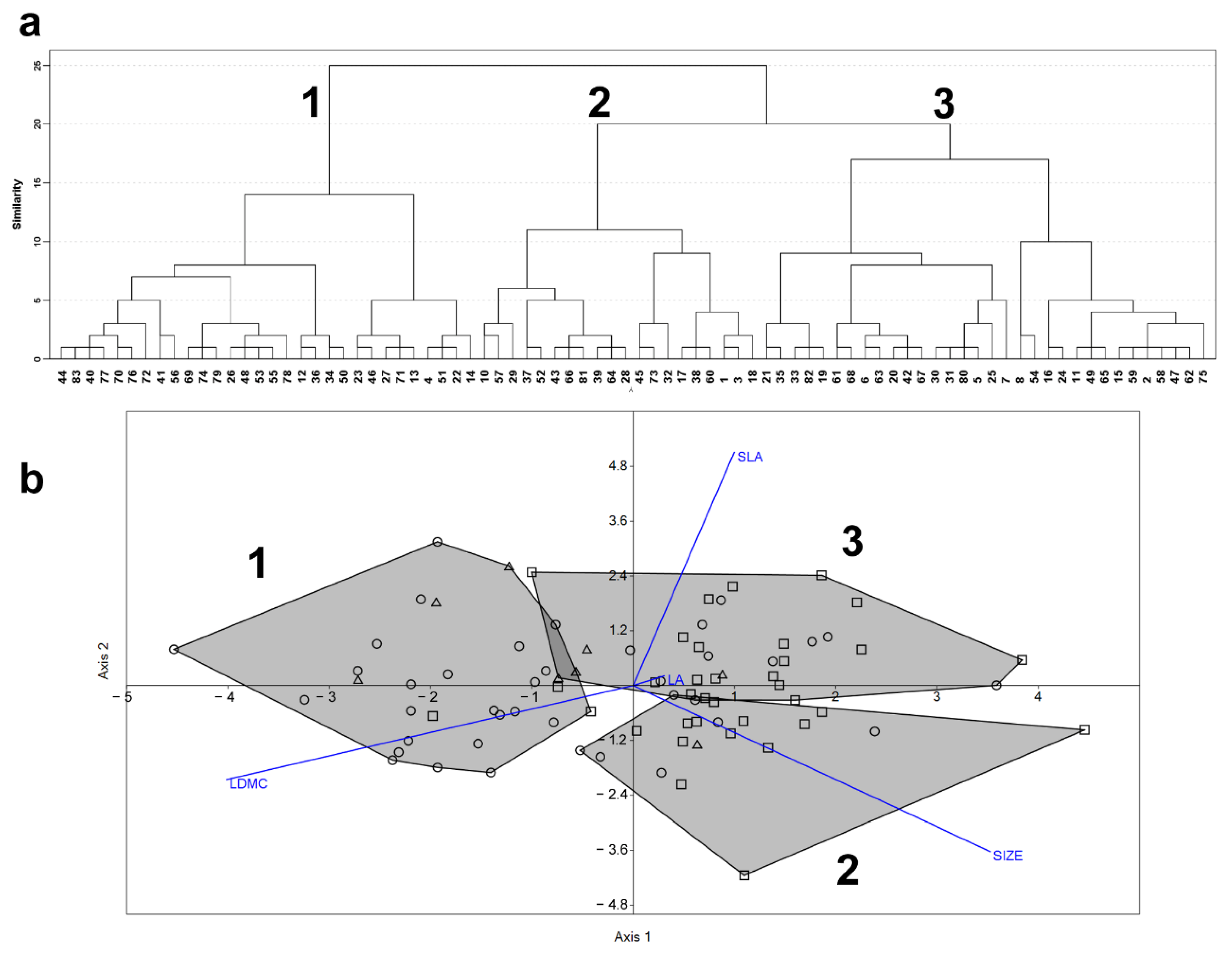
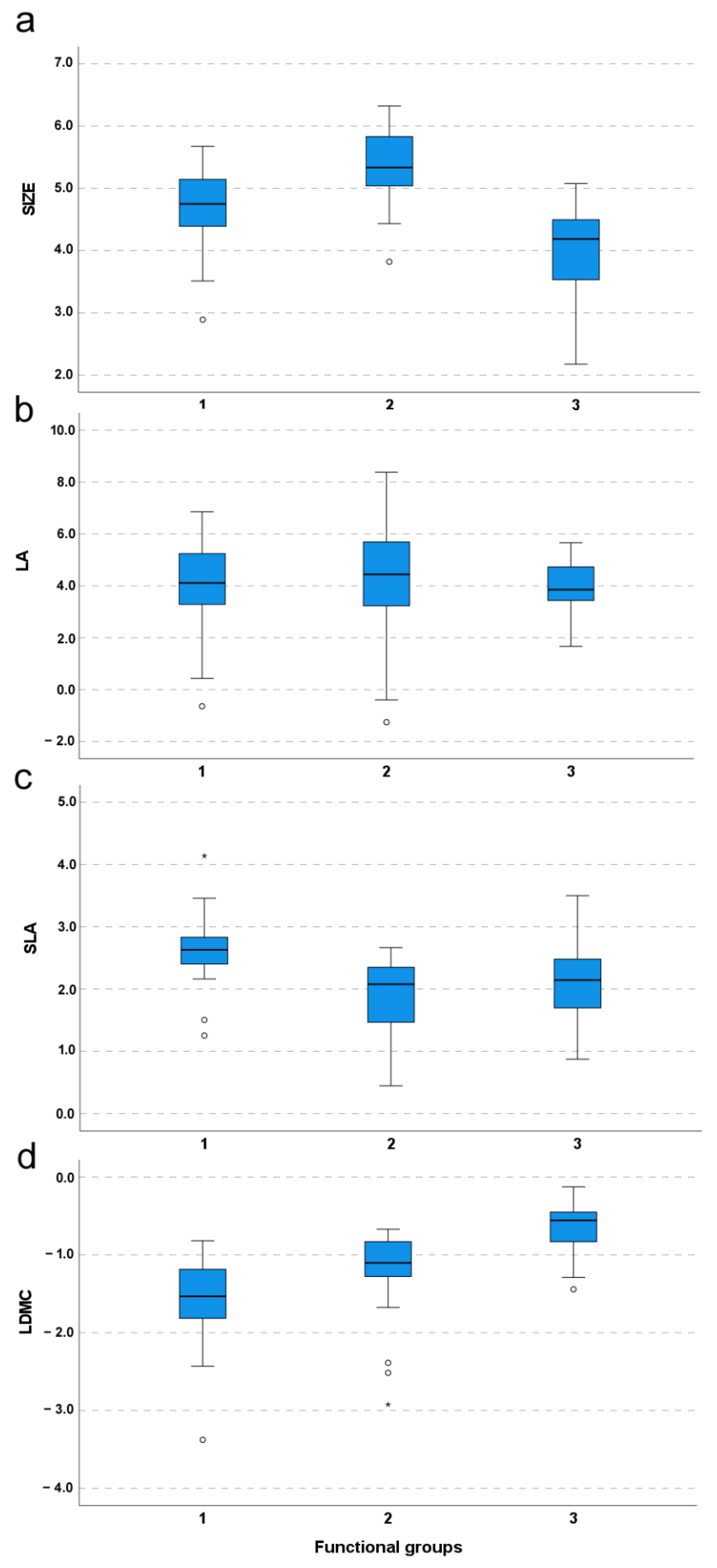
2.1. Species Functional Classification and Trait Contribution to Species Functional Differentiation (Q1)
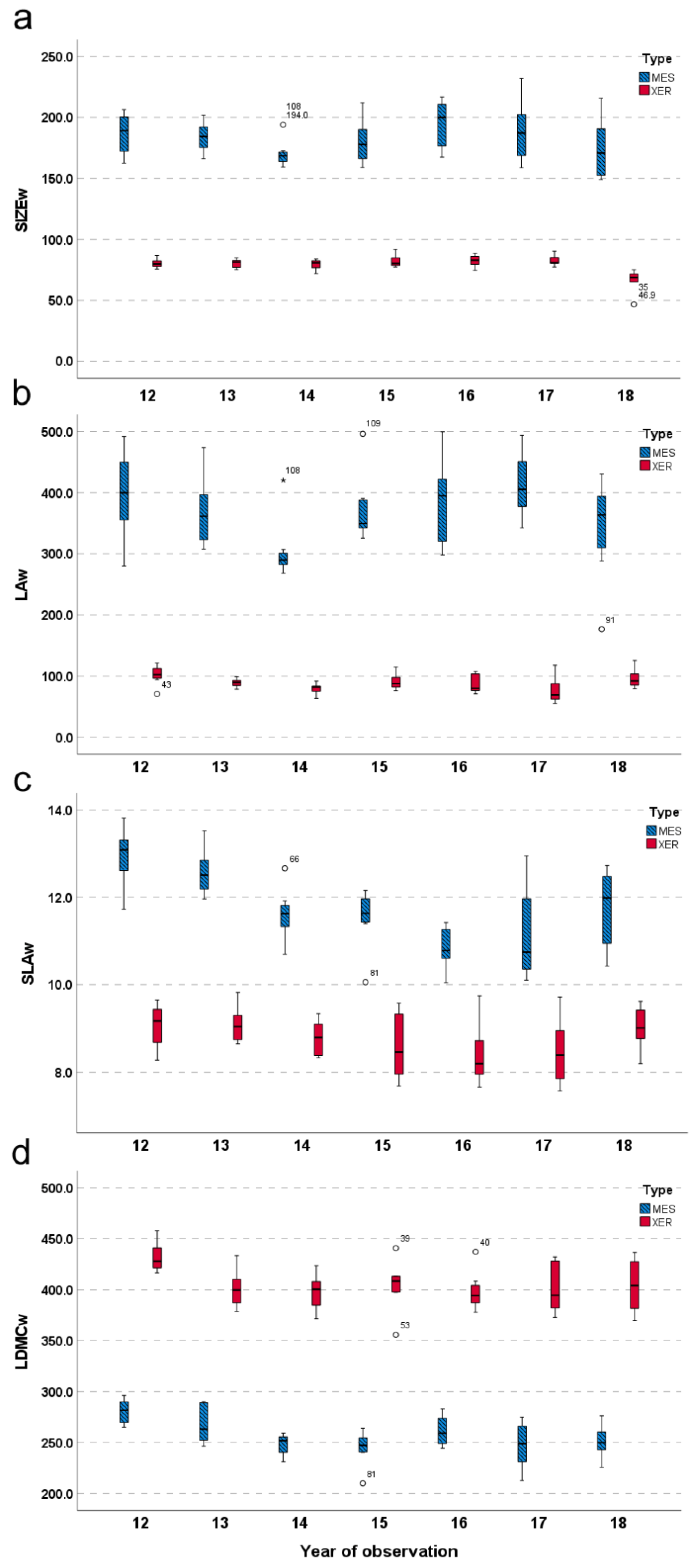
2.2. Community-Level Effect of Species Functional Differentiation (Q2)
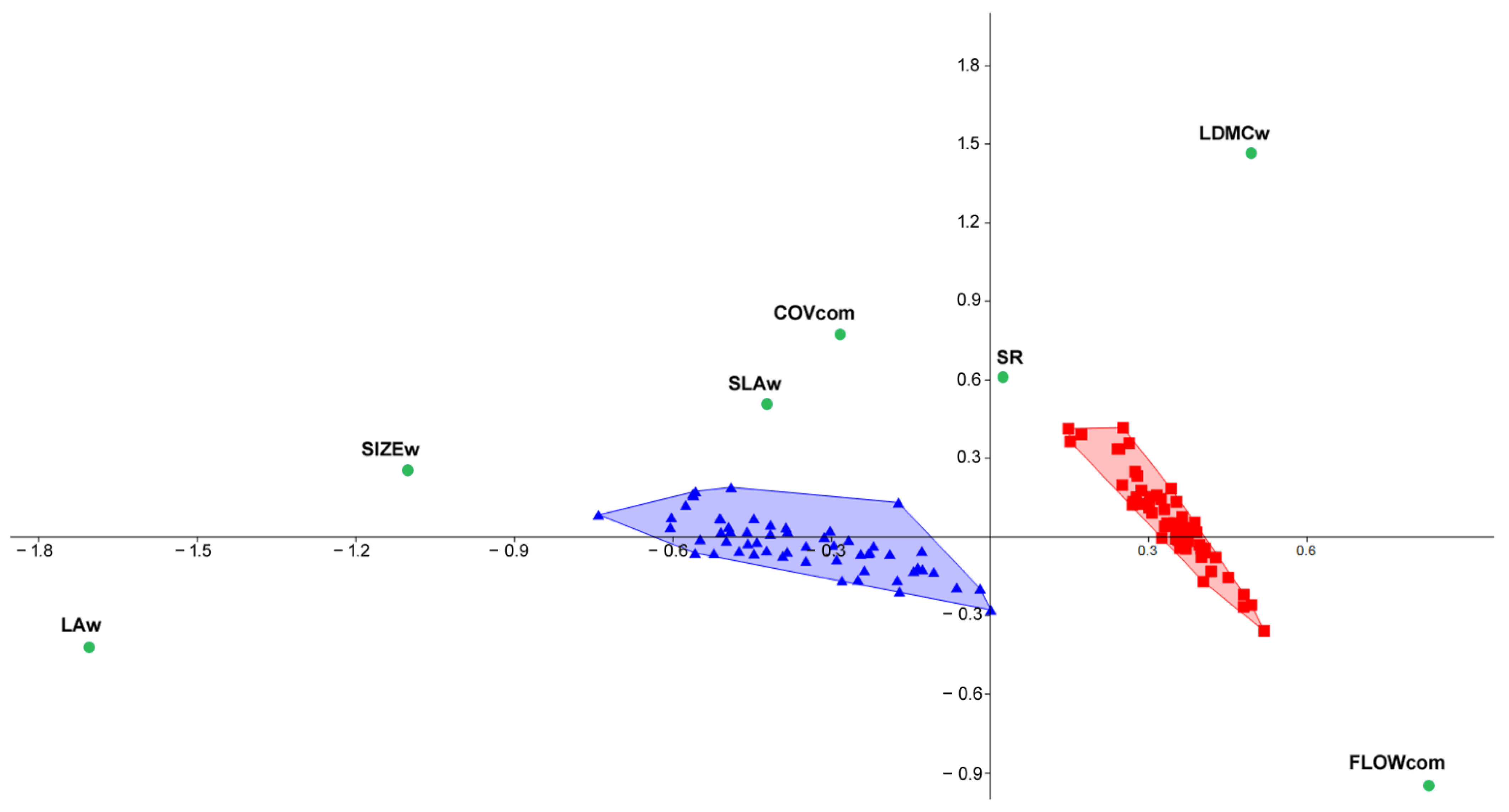
2.3. Investigating Relationships between Functional and Productivity Patterns of Species and Related Communities (Q3)
3. Discussion
3.1. Functional Characterization of Species and Communities (Q1–2)
3.2. Linking Functional Features to Inter-Annual Productivity Patterns (Q3)
4. Materials and Methods
4.1. Study Sites
4.2. Trait and Plant Sampling for Functional Measures
4.3. Floristic Composition, Plant Cover and Flowering Rates
4.4. Data Analysis
4.4.1. Species Functional Classification and Trait Contribution to Species Functional Differentiation (Q1)
4.4.2. Community-Level Effect of Species Functional Differentiation (Q2)
4.4.3. Linking Functional Features to Productivity Patterns of Species and Communities (Q3)
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Myers, N.; Mittermeier, R.A.; Mittermeier, C.G.; Da Fonseca, G.A.; Kent, J. Biodiversity hotspots for conservation priorities. Nature 2000, 403, 853–858. [Google Scholar] [CrossRef]
- Thompson, J.D. Plant Evolution in the Mediterranean; Oxford University Press: Oxford, UK, 2005. [Google Scholar]
- Cañadas, E.M.; Fenu, G.; Peñas, J.; Lorite, J.; Mattana, E.; Bacchetta, G. Hotspots within hotspots: Endemic plant richness, environmental drivers, and implications for conservation. Biol. Conserv. 2014, 170, 282–291. [Google Scholar] [CrossRef]
- Wilson, J.B.; Peet, R.K.; Dengler, J.; Pärtel, M. Plant species richness: The word records. J. Veg. Sci. 2012, 23, 796–802. [Google Scholar] [CrossRef]
- Schröter, D.; Cramer, W.; Leemans, R.; Prentice, I.C.; Araújo, M.B.; Arnell, N.W.; Bondeau, A.; Bugmann, H.; Carter, T.R.; Gracia, C.A.; et al. Ecosystem service supply and vulnerability to global change in Europe. Science 2005, 310, 1333–1337. [Google Scholar] [CrossRef]
- Bozinovic, F.; Bastias, D.A.; Boher, F.; Clavijo-Baquet, S.; Estay, S.A.; Angilletta, M.J. The Mean and Variance of Environmental Temperature Interact to Determine Physiological Tolerance and Fitness. Physiol. Biochem. Zool. 2011, 84, 543–552. [Google Scholar] [CrossRef]
- Gottfried, M.; Pauli, H.; Futschik, A.; Akhalkatsi, M.; Barančok, P.; Benito Alonso, J.L.; Coldea, G.; Dick, J.; Erschbamer, B.; Fernández Calzado, M.A.R.; et al. Continent-wide response of mountain vegetation to climate change. Nat. Clim. Chang. 2012, 2, 111–115. [Google Scholar] [CrossRef]
- Pauli, H.; Gottfried, M.; Dullinger, S.; Abdaladze, O.; Akhalkatsi, M.; Alonso, J.L.B.; Coldea, G.; Dick, J.; Erschbamer, B.; Calzado, M.R.F.; et al. Recent plant diversity change on Europe’s mountain summits. Science 2012, 336, 353–355. [Google Scholar] [CrossRef]
- Gargano, D.; Mingozzi, A.; Massolo, A.; Rinaldo, S.; Bernardo, L. Patterns of vegetation cover/dynamics in a protected Mediterranean mountain area: Influence of the ecological context and protection policy. Plant Biosyst. 2011, 146, 9–18. [Google Scholar] [CrossRef]
- Petraglia, A.; Cacciatori, C.; Chelli, S.; Fenu, G.; Calderisi, G.; Gargano, D.; Abeli, T.; Orsenigo, S.; Carbognani, M. Litter decomposition: Effects of temperature driven by soil moisture and vegetation type. Plant Soil 2019, 435, 187–200. [Google Scholar] [CrossRef]
- Peco, B.; Navarro, E.; Carmona, C.P.; Medina, N.G.; Marques, M.J. Effects of grazing abandonment on soil multifunctionality: The role of plant functional traits. Agric. Ecosyst. Environ. 2017, 249, 215–225. [Google Scholar] [CrossRef]
- Isbell, F.I.; Tilman, D.; Polasky, S.; Loreau, M. The biodiversity-dependent ecosystem service debt. Ecol. Lett. 2015, 18, 119–134. [Google Scholar] [CrossRef] [PubMed]
- Schirpke, U.; Kohler, M.; Leitinger, G.; Fontana, V.; Tasser, E.; Tappeiner, U. Future impacts of changing land-use and climate on ecosystem services of mountain grassland and their resilience. Ecosyst. Serv. 2017, 26, 79–94. [Google Scholar] [CrossRef] [PubMed]
- Attorre, F.; Abeli, T.; Bacchetta, G.; Farcomeni, A.; Fenu, G.; De Sanctis, M.; Gargano, D.; Peruzzi, L.; Montagnani, C.; Rossi, G. How to include the impact of climate change in the extinction risk assessment of policy plant species? J. Nat. Conserv. 2018, 44, 43–49. [Google Scholar] [CrossRef]
- Fridley, J.D.; Grime, J.P.; Askew, A.P.; Moser, B.; Stevens, C.J. Soil heterogeneity buffers community response to climate change in species-rich grassland. Glob. Change Biol. 2011, 17, 2002–2011. [Google Scholar] [CrossRef]
- Godfree, R.; Lepschi, B.; Reside, A.; Bolger, T.; Robertson, B.; Marshall, D.; Carnegie, M. Multiscale topoedaphic heterogeneity increases resilience and resistance of a dominant grassland species to extreme drought and climate change. Glob. Chang. Biol. 2011, 17, 943–958. [Google Scholar] [CrossRef]
- García-Palacios, P.; Maestre, F.T.; Bardgett, R.D.; De Kroon, H. Plant responses to soil heterogeneity and global environmental change. J. Ecol. 2012, 100, 1303–1314. [Google Scholar] [CrossRef]
- Bjorkman, A.D.; Myers-Smith, I.H.; Elmendorf, S.C.; Normand, S.; Rüger, N.; Beck, P.S.A.; Blach-Overgaard, A.; Blok, D.; Cornelissen, J.H.C.; Forbes, B.C.; et al. Plant functional trait change across a warming tundra biome. Nature 2018, 562, 57–62. [Google Scholar] [CrossRef]
- Sebastià, M.T. Role of topography and soils in a grassland structuring at the landscape and community scales. Basic Appl. Ecol. 2004, 5, 331–346. [Google Scholar] [CrossRef]
- Gargano, D.; Vecchio, D.; Bernardo, L. Plant-soil relationships in fragments of Mediterranean snow beds: Ecological and conservation implications. Plant Ecol. 2010, 207, 175–189. [Google Scholar] [CrossRef]
- Grime, J.P. Dominant and subordinate components of plant communities: Implications for succession, stability and diversity. In Colonization, Succession and Stability; Gray, A.J., Crawley, M.J., Eds.; Blackwell Scientific: Oxford, UK, 1984; pp. 413–428. [Google Scholar]
- May, R.M. Stability and Complexity in Model Ecosystems; Princeton Landmarks in Biology; Princeton University Press: Princeton, NJ, USA, 1973. [Google Scholar]
- Doak, D.F.; Bigger, D.; Harding, E.; Marvier, M.; O’Malley, R.; Thomson, D. The statistical inevitability of stability-diversity relationships in community ecology. Am. Nat. 1998, 151, 264–276. [Google Scholar] [CrossRef]
- Figueroa, M.E.; Davy, A.J. Response of Mediterranean Grassland Species to Changing Rainfall. J. Ecol. 1991, 79, 925–941. [Google Scholar] [CrossRef]
- Grime, J.P.; Brown, V.K.; Thompson, K.; Masters, G.J.; Hillier, S.H.; Clarke, I.P.; Askew, A.P.; Corker, D.; Kielty, J.P. The response of two contrasting limestone grasslands to simulated climate change. Science 2000, 289, 762–765. [Google Scholar] [CrossRef] [PubMed]
- Vogel, A.; Scherer-Lorenzen, M.; Weigelt, A. Grassland resistance and resilience after drought depends on management intensity and species richness. PLoS ONE 2012, 7, e36992. [Google Scholar] [CrossRef]
- Tilman, D.; Downing, J.A. Biodiversity and stability in grasslands. Nature 1994, 367, 363–365. [Google Scholar] [CrossRef]
- Tilman, D.; Reich, P.B.; Knops, J.M.H. Biodiversity and ecosystem stability in a decade-long grassland experiment. Nature 2006, 441, 629–632. [Google Scholar] [CrossRef]
- Liu, J.; Liu, D.; Xu, K.; Gao, L.M.; Ge, X.-J.; Burgess, K.S.; Cadotte, M.W. Biodiversity explains maximum variation in productivity under experimental warming, nitrogen addition, and grazing in mountain grasslands. Ecol. Evol. 2018, 8, 10094–10112. [Google Scholar] [CrossRef]
- Reich, P.B.; Knops, J.; Tilman, D.; Craine, J.; Ellsworth, D.; Tjoelker, M.; Lee, T.; Wedin, D.; Naeem, S.; Bahauddin, D.; et al. Plant diversity enhances ecosystem responses to elevated CO2 and nitrogen deposition. Nature 2001, 410, 809–810. [Google Scholar] [CrossRef]
- Pfisterer, A.B.; Schmid, B. Diversity-dependent production can decrease the stability of ecosystem functioning. Nature 2002, 416, 84–86. [Google Scholar] [CrossRef]
- Van Ruijven, J.; Berendse, F. Diversity enhances community recovery, but not resistance, after drought. J. Ecol. 2010, 88, 634–644. [Google Scholar] [CrossRef]
- Olsen, S.L.; Töpper, J.P.; Skarpaas, O.; Vandvik, V. From facilitation to competition: Temperature-driven shift in dominant plant interactions affects population dynamics in seminatural grasslands. Glob. Chang. Biol. 2016, 22, 1915–1926. [Google Scholar] [CrossRef]
- Isbell, F.I.; Wilsey, B.J. Increasing native, but not exotic, biodiversity increases aboveground productivity in ungrazed and intensely grazed grasslands. Oecologia 2011, 165, 771–781. [Google Scholar] [CrossRef]
- MacLean, S.A.; Beissinger, S.R. Species’ traits as predictors of range shifts under contemporary climate change: A review and meta-analysis. Glob. Chang. Biol. 2017, 23, 4094–4105. [Google Scholar] [CrossRef] [PubMed]
- Blonder, B.; Kapas, R.E.; Dalton, R.M.; Graae, B.J.; Heiling, J.M.; Opedal, Ø.H. Microenvironment and functional-trait context dependence predict alpine plant community dynamics. J. Ecol. 2018, 106, 1323–1337. [Google Scholar] [CrossRef]
- Joswig, J.S.; Wirth, C.; Schuman, M.C.; Kattge, J.; Reu, B.; Wrigth, I.J.; Sippel, S.D.; Rüger, N.; Richter, R.; Schaepman, M.E.; et al. Climatic and soil factors explain the two-dimensional spectrum of global plant trait variation. Nature Ecol. Evol. 2022, 6, 36–50. [Google Scholar] [CrossRef] [PubMed]
- Díaz, S.; Kattge, J.; Cornelissen, J.H.C.; Wright, I.J.; Lavorel, S.; Dray, S.; Reu, B.; Kleyer, M.; Wirth, C.; Prentice, I.C.; et al. The global spectrum of plant form and function. Nature 2015, 529, 167–171. [Google Scholar] [CrossRef]
- Grime, J.P.; Pierce, S. The Evolutionary Strategies That Shape Ecosystems; John Wiley & Sons: Hoboken, NJ, USA, 2012. [Google Scholar]
- Pierce, S.; Negreiros, D.; Cerabolini, B.E.L.; Kattge, J.; Diaz, S.; Kleyer, M.; Shipley, B.; Wright, S.J.; Soudzilvskaia, N.A.; Onipchenko, V.G.; et al. A global method for calculating plant CSR ecological strategies applied across biomes word-wide. Funct. Ecol. 2017, 31, 444–457. [Google Scholar] [CrossRef]
- Bruelheide, H.; Dengler, J.; Purschke, O.; Lenoir, J.; Jiménez-Alfaro, B.; Hennekens, S.M.; Botta-Dukát, Z.; Chytrý, M.; Field, R.; Jansen, F.; et al. Global trait-environment relationships of plant communities. Nat. Ecol. Evol. 2018, 2, 1906–1917. [Google Scholar] [CrossRef]
- Gargano, D.; Aiello, S.; Abeli, T.; Schettino, A.; Bernardo, L. Monitoring biodiversity patterns in three Mediterranean mountain pastures in the Pollino National Park (S-Italy). Plant Sociol. 2017, 54, 51–59. [Google Scholar] [CrossRef]
- Fry, E.; Power, S.A.; Manning, P. Trait-based classification and manipulation of plant functional groups for biodiversity-ecosystem function experiments. J. Veg. Sci. 2014, 25, 248–261. [Google Scholar] [CrossRef]
- Garnier, E.; Shipley, B.; Roumet, C.; Laurent, G. A standardized protocol for the determination of specific leaf area and leaf dry matter content. Funct. Ecol. 2001, 15, 688–695. [Google Scholar] [CrossRef]
- Cornelissen, J.H.C.; Lavorel, S.; Garnier, E.; Díaz, S.; Buchmann, N.; Gurvich, D.E.; Reich, P.B.; Steege, H.T.; Morgan, H.D.; van der Heijden, M.G.A.; et al. A handbook of protocols for standardised and easy measurement of plant functional traits worldwide. Aust. J. Bot. 2003, 51, 335–380. [Google Scholar] [CrossRef]
- Laughlin, D.C.; Fulé, P.Z.; Huffman, D.W.; Crouse, J.; Laliberté, E. Climatic constraints on trait-based forest assembly. J. Ecol. 2011, 99, 1489–1499. [Google Scholar] [CrossRef]
- Grime, J.P. Evidence for the existence of three primary strategies in plants and its relevance to ecological and evolutionary theory. Amer. Natur. 1977, 111, 1169–1194. [Google Scholar] [CrossRef]
- Hodgson, D.A.; Rastetter, E.B.; Shaver, G.R.; Agren, G.I. Effects of plant growth characteristics on biogeochemistry and community composition in a changing climate. Ecosystem 1999, 2, 367–382. [Google Scholar] [CrossRef]
- Garnier, E.; Fayolle, A.; Navas, M.-L.; Damgaard, C.; Cruz, P.; Hubert, D.; Richarte, J.; Autran, P.; Leurent, C.; Violle, C. Plant demographic and functional responses to management intensification: A long-term study in a Mediterranean rangeland. J. Ecol. 2018, 106, 1363–1376. [Google Scholar] [CrossRef]
- Federico, S.; Avolio, E.; Pasqualoni, L.; De Leo, L.; Sempreviva, A.M.; Bellecci, C. Preliminary results of a 30-year daily rainfall data base in southern Italy. Atmos. Res. 2009, 94, 641–651. [Google Scholar] [CrossRef]
- Federico, S.; Pasqualoni, L.; Avolio, E.; Bellecci, C. Brief communication “Calabria daily rainfall from 1970 to 2006”. Nat. Hazards Earth Syst. Sci. 2010, 10, 717–722. [Google Scholar] [CrossRef]
- Biondi, E.; Ballelli, S.; Allegrezza, M.; Zuccarello, V. La vegetazione dell’ordine Brometalia erecti Br.-Bl. 1936 nell’Appennino (Italia). Fitosociologia 1995, 30, 3–45. [Google Scholar]
- Bonin, G. Première contribution a l’étude des pelouses mésophile et des groupements hygrophiles du Monte Pollino. Phyton 1972, 14, 271–280. [Google Scholar]
- Schneider, C.A.; Rasband, W.S.; Eliceiri, K.W. NIH Image to ImageJ: 25 years of image analysis. Nat. Methods 2012, 9, 671–675. [Google Scholar] [CrossRef]
- van der Maarel, E. Trasformation of cover-abundance values in phytosociology and its effects on community similarity. Vegetatio 1979, 39, 97–144. [Google Scholar]
- Mendes, R.S.; Evangelista, L.R.; Thomaz, S.M.; Agostinho, A.A.; Gomez, L.C. A unified index to measure ecological diversity and species rarity. Ecography 2008, 31, 450–456. [Google Scholar] [CrossRef]
- Vymazalová, M.; Axmanová, I.; Tichý, L. Effect of intra-seasonal variability on vegetation data. J. Veg. Sci. 2021, 23, 978–984. [Google Scholar] [CrossRef]
- Legendre, P.; Legendre, L. Numerical Ecology; Elsevier: Amsterdam, The Netherlands, 1998. [Google Scholar]
- Hammer, Ø.; Harper, D.A.T.; Ryan, P.D. PAST: Paleontological Statistics Software Package for Education and Data Analysis. Palaent. Electron. 2001, 4, 9. [Google Scholar]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| FG | 1 | 2 | 3 | Total | CC(%) |
|---|---|---|---|---|---|
| 1 | 27 | 3 | 2 | 32 | 84.4 |
| 2 | 1 | 19 | 0 | 20 | 95.0 |
| 3 | 0 | 0 | 30 | 30 | 100.0 |
| Total | 28 | 22 | 32 | 82 |
| All Plant Species | ||||||||||||
| Measures of Species Productivity | ||||||||||||
| COVave | COVvar | FLOWave | FLOWvar | |||||||||
| Functional trait | N | t | p | N | t | p | N | t | p | N | t | p |
| SIZE | 79 | 0.985 | 0.328 | 79 | 1.031 | 0.306 | 79 | −2.052 | 0.04 * | 80 | −1.080 | 0.284 |
| LA | 79 | 1.057 | 0.294 | 79 | 1.146 | 0.256 | 79 | −1.847 | 0.07 | 80 | −1.392 | 0.168 |
| SLA | 79 | −0.098 | 0.922 | 79 | 1.386 | 0.170 | 79 | 1.806 | 0.08 | 80 | 0.752 | 0.454 |
| LDMC | 79 | 1.301 | 0.197 | 79 | 3.105 | 0.003 ** | 79 | 2.267 | 0.03 * | 80 | 2.113 | 0.04 * |
| Mesophile plant species | ||||||||||||
| Measures of species productivity | ||||||||||||
| COVave | COVvar | FLOWave | FLOWvar | |||||||||
| Functional trait | N | t | p | N | t | p | N | t | p | N | t | p |
| SIZE | 36 | 0.267 | 0.791 | 36 | 1.758 | 0.089 | 33 | −0.732 | 0.470 | 34 | −0.215 | 0.831 |
| LA | 36 | 2.949 | 0.006 ** | 36 | −0.647 | 0.523 | 33 | −0.12 | 0.991 | 34 | −0.165 | 0.870 |
| SLA | 36 | 0.058 | 0.954 | 36 | 0.346 | 0.732 | 33 | 0.448 | 0.657 | 34 | −0.340 | 0.737 |
| LDMC | 36 | 2.298 | 0.028 * | 36 | 0.711 | 0.482 | 33 | 1.176 | 0.250 | 34 | 0.761 | 0.453 |
| Xerophile plant species | ||||||||||||
| Measures of species productivity | ||||||||||||
| COVave | COVvar | FLOWave | FLOWvar | |||||||||
| Functional trait | N | t | p | N | t | p | N | t | p | N | t | p |
| SIZE | 43 | 0.042 | 0.967 | 43 | 0.000 | 1.000 | 45 | −0.979 | 0.333 | 46 | −0.515 | 0.610 |
| LA | 43 | −0.657 | 0.515 | 43 | 1.625 | 0.112 | 45 | −1.695 | 0.098 | 46 | −1.290 | 0.204 |
| SLA | 43 | −0.285 | 0.777 | 43 | 1.528 | 0.135 | 45 | 1.581 | 0.122 | 46 | 1.058 | 0.296 |
| LDMC | 43 | 0.549 | 0.586 | 43 | 2.518 | 0.016* | 45 | 1.719 | 0.93 | 46 | 1.577 | 0.123 |
| Overall Vegetation Data | |||||||||
| Measures of Community Productivity | |||||||||
| SR | COVcom | FLOWcom | |||||||
| Functional trait | N | t | p | t | p | t | p | ||
| SIZEw | 112 | −0.109 | 0.254 | 0.642 ** | <0.001 | −0.598 ** | <0.001 | ||
| LAw | 112 | −0.109 | 0.252 | 0.634 ** | <0.001 | −0.551 ** | <0.001 | ||
| SLAw | 112 | 0.053 | 0.576 | 0.510 ** | <0.001 | −0.405 ** | <0.001 | ||
| LDMCw | 112 | 0.010 | 0.913 | −0.560 ** | <0.001 | 0.516 ** | <0.001 | ||
| Samples from the mesophile community | |||||||||
| Measures of community productivity | |||||||||
| SR | COVcom | FLOWcom | |||||||
| Functional trait | N | t | p | N | t | p | N | t | p |
| SIZEw | 56 | −0.549 ** | <0.001 | 56 | 0.403 ** | 0.002 | 56 | −0.412 ** | 0.002 |
| LAw | 56 | −0.403 ** | 0.002 | 56 | 0.255 | 0.058 | 56 | −0.283 * | 0.034 |
| SLAw | 56 | 0.255 | 0.058 | 56 | −0.248 | 0.066 | 56 | 0.286 * | 0.032 |
| LDMCw | 56 | −0.003 | 0.982 | 56 | 0.049 | 0.719 | 56 | 0.000 | 1.000 |
| Samples from the xerophile community | |||||||||
| Measures of community productivity | |||||||||
| SR | COVcom | FLOWcom | |||||||
| Functional trait | N | t | p | N | t | p | N | t | p |
| SIZEw | 56 | 0.041 | 0.397 | 56 | −0.299 * | 0.025 * | 56 | −0.366 * | 0.011 |
| LAw | 56 | −0.061 | 0.653 | 56 | 0.070 | 0.607 | 56 | 0.162 | 0.232 |
| SLAw | 56 | 0.109 | 0.424 | 56 | 0.272 * | 0.042 | 56 | 0.158 | 0.245 |
| LDMCw | 56 | −0.122 | 0.370 | 56 | 0.266 * | 0.048 | 56 | −0.090 | 0.509 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Gargano, D.; Fenu, G.; Passalacqua, N.G.; Schettino, A.; Bernardo, L. Relating Trait Variation to Species and Community Productivity in Contrasting Oro-Mediterranean Pastures: A 7-Years Study in the Pollino National Park (S-Italy). Plants 2022, 11, 2471. https://0-doi-org.brum.beds.ac.uk/10.3390/plants11192471
Gargano D, Fenu G, Passalacqua NG, Schettino A, Bernardo L. Relating Trait Variation to Species and Community Productivity in Contrasting Oro-Mediterranean Pastures: A 7-Years Study in the Pollino National Park (S-Italy). Plants. 2022; 11(19):2471. https://0-doi-org.brum.beds.ac.uk/10.3390/plants11192471
Chicago/Turabian StyleGargano, Domenico, Giuseppe Fenu, Nicodemo G. Passalacqua, Aldo Schettino, and Liliana Bernardo. 2022. "Relating Trait Variation to Species and Community Productivity in Contrasting Oro-Mediterranean Pastures: A 7-Years Study in the Pollino National Park (S-Italy)" Plants 11, no. 19: 2471. https://0-doi-org.brum.beds.ac.uk/10.3390/plants11192471