1. Introduction
Onion is second most important vegetable crop after tomato with global production of 104.6 million tonnes from 5.48 million hectare area [
1]. Asia accounts for the maximum share (66.8%) of total onion production in the world, followed by Africa (13.5%), Europe (9.9%), the Americas (9.6), and Oceania (0.3%) [
1]. Maximum bulb onion production (26.74 million tonnes) in India consists of a 1.43 million hectare area with a productivity of 18.64 t/ha, which is less than other countries, viz. Republic of Korea (79.6 t/ha), USA, Australia, Spain, and Japan. In India, onions are grown in a wide range of climatic conditions [
2]. Onion is consumed both fresh as well as in the form of processed products, owing to its nutritional and medicinal properties [
3]. Onion contains bioactive compounds (quercetin, rutin etc.) and its regular consumption reduces the risk of cancer, heart diseases, diabetes, and production of reactive oxygen species [
4]. Most onions in India are produced during the post-monsoon season; owing to shallow roots, onions are highly vulnerable to climatic conditions such as salinity and drought because of inadequate rainfall and low moisture content [
5]. Onion is a salt-sensitive crop and because of its threshold level (1.4 dS m
−1), the per unit increase in EC reduces 18.52% of the tuber yield [
6]. The world’s 950 mha area of land (10%), 300 mha cultivated land (20%), and 230 m ha (50%) irrigated land are distressed by salt stress. Annually, about a USD 12 billion loss is estimated because of salt stress, which adversely affects agriculture production. Salinity changes the morpho-physiological, biochemical, and metabolic processes and affects onion growth and development [
7]. Soil salinity increases the Na
+ and Cl
− concentration in different plant parts, which ultimately influences the ionic activity in plant cells [
8]. Salt stress-induced oxidative stress leads to leakage of ions, plasmolysis, membrane damage, disturbed nutrient flux, and ROS detoxification systems. These alterations adversely affect respiration, photosynthesis, hormonal balance, antioxidants activity, water use efficiency, transpiration, and plasma membrane functioning [
9]. The adverse effect of salt stress on onion morphology includes reduced bulb weight, bulb diameter, plant height, leaf number per plant, root growth, and crop cycle [
6,
10]. High salinity decreases the production of large bulbs and maximum decreases were observed at 2.8 dS m
−1 [
11]. Plants develop different strategies such as avoidance and tissue tolerance to reduce the negative effects of salt stress. Plants also produce various compatible solutes such as glycine betaine, proline, and sugars to regulate osmotic pressure [
12]. Likewise, plants produce proteins and antioxidants to counter the negative effect of reactive oxygen species. Tissue tolerance protects the plant cells by executing ion compartmentalization in plant tissues [
13]. A low Na
+/K
+ ratio also plays a significant role in balancing the osmotic pressure and membrane stability [
7].
The present study was initiated because there is not much literature on the effect of salinity stress on Indian short-day onions. To develop a breeding strategy for the development of salinity stress-tolerant onions, the basic step is to evaluate the onion accessions and to identify the promising accessions for formulating future strategies.
In view of the scant information on the effect of salinity stress on Indian short-day onions, present study was initiated with the objective to determine the effect of salinity stress on growth and development of onions. Keeping in view this objective, we formulated and tested three research hypothesis, i.e., (i) whether salinity stress negatively affects the bulb yield and yield associated traits in onion, (ii) whether salt tolerant and salt sensitive accessions differ in physicochemical responses?, and (iii) of the most important parameters (morphological, physiological, and biochemical), which ones need to be considered for the screening and development of salt tolerant onion cultivars? To achieve these objectives, thirty-six onion cultivars were considered in the present investigation.
2. Materials and Methods
2.1. Characterization and Evaluation of Onion Accessions
Thirty-six diverse Indian short-day onion accessions including the released cultivars and some of the promising advance lines were collected from the National Horticulture Research and Development Foundation (NHRDF), Karnal; ICAR-Directorate of Onion and Garlic Research, Pune; and ICAR-Indian Agricultural Research Institute (IARI), New Delhi, India (
Table 1). Seeds were sown in nursery beds and seedlings were transplanted in three replications using randomized complete block design (RCBD) under control and under saline conditions (ECiw 7 dS m
−1) in microplots during the rabi season of 2019–20.
In each replication, 20 plants were maintained following a plant geometry of 15 × 10 cm. Soil samples were collected before sowing and after harvesting of the crop of both the treatments and analysed (
Table 2). For saline treatment, natural saline ground water (ECiw~18 dS m
−1) was used for the preparation of desired saline water salinity (ECiw 7 dS m
−1), whereas the normal water of ECiw~1.04 dS m
−1 was used for control treatment. Seven days after transplanting, the irrigation was given as per treatment and subsequent irrigation was given as per crop need on the basis of 100% evapotranspiration (ET), and a total of 17 irrigations were applied during the whole crop cycle. After soil analysis, the required dose of NPK was applied and the recommended packages of practices were followed for growing a good crop. As per recommendations, 50% nitrogen and 100% phosphorus and potassium were applied before transplanting, and the remaining nitrogen dose was applied in two split doses, i.e., 30 and 45 days after transplanting. Harvesting of onion bulbs was done when the leaves turned yellow and more than 50 percent neckfall was observed. The bulbs were dug out along with their leaves and the tops were removed, leaving a 2 cm neck portion attached to the bulbs. Harvested bulbs were kept under shade for curing and then sorting before grading was done.
2.2. Morphometric and Yield-Related Traits
The data recorded were concerned the plant height (cm), no. of leaves/plant, bulb diameter (cm), bulb yield/plant, and total soluble solids (TSS, °Brix). The replicate-wise data of morphological traits of 10 selected plants were measured after 100 days of transplanting. The bulb yield of 10 plants was harvested separately from control and saline environments. Bulbs were inspected visually for rotting or sprouting damage. Equatorial bulb diameter (cm) of the same 10 plants was measured with the help of the digital Vernier Caliper. The TSS content of the selected bulbs was measured just after bulb harvesting with the help of a portable hand refractometer (Erma Inc., Tokyo, Japan) as °Brix (%) at 20 °C.
2.3. Physio-Biochemical and Ion Estimation
A total of 36 accessions were characterized under a saline environment, and out of these, 14 contrasting accessions were further selected for detailed studies of physiological, biochemical, and antioxidant enzymes, as well as ion contents. Out of the 14 accessions, 7 accessions (POS35, L-28, GJWO-1, POS36, L-744, POS37, POS38) had ≤20% yield reduction under saline conditions and were categorised as salt-tolerant. The other 7 accessions (ALR, GJWO3, KKR, L-652, Agrifound White, L-920, Bhima Dark Red) had ≥40% yield reduction and were categorized as salt-sensitive. These 14 accessions were again transplanted in microplots during the rabi season of 2020–21 for physiological, biochemical, and ionic analysis, and the package of practices and salinity treatments were followed as per the previous year’s (2019–20) experiment.
2.3.1. Physiological Traits
Physiological and biochemical traits were measured at 55 days after transplanting. The method given by Weatheraly [
14] was used to estimate the relative water content (RWC). The leaf membrane thermostability index was estimated by using the electrolyte leakage percentage following the procedure of Dionisio-Sese and Tobita [
15]. Photosynthesis rate (Pn), transpiration rate (E), and stomatal conductance (gS) of the 3rd fully expanded leaves were estimated using a portable photosynthetic system (Li 6800, Li-Cor Biosciences, Lincoln, NE, USA) following the method of Kumar et al. [
16].
2.3.2. Biochemical Traits
The method given by Bates et al. [
17] was used to estimate the proline content. H
2O
2 content was calculated as per the method given by Loreto and Velikova [
18]. MDA content was measured at 532 and 600 nm by using same the supernatant which was used for the estimation of H
2O
2 concentration [
19]. A modified approach was followed for the extraction of antioxidant enzymes, superoxide dismutase (SOD), and ascorbate peroxidase (APX) [
20]. Peroxidase (POX) was estimated as per the method suggested by Beauchamp and Fridovich [
21]. One unit of APX corresponded to a change in O.D. of 1.0 per minute [
22]. The POX activity was determined by using 1.0 mol of H
2O
2 per minute [
23]. Based on the breakdown of H
2O
2 at 240 nm, the catalase (CAT) activity was measured for 1 min [
24].
2.3.3. Ion Concentration
Na+ and K+ contents of leaves, roots, and bulbs were determined at the time of harvesting using the flame photometer (Systronics Flame Photometer 128, Olathe, KS, USA) and estimated as mg g−1 of dry weight.
2.4. Statistical Analysis
In the first experiment, five observations, i.e., plant height (cm), number of leaves/plant, equatorial bulb diameter (cm), bulb yield/plant (g), and TSS (°Brix) were recorded for preliminary screening. Onion accessions were categorized on the basis of bulb yield reduction (%) under salinity stress in comparison to the control conditions. Based on percent bulb yield reduction, seven tolerant (≤20%) and seven susceptible (≥40%) accessions were selected for a second experiment to determine the physio-biochemical basis of salinity tolerance.
In experiment 2, all the recorded parameters were put on a Microsoft excel sheet thematically and a test of normality for each parameter was performed through the Shapiro–Wilk test to comply with the assumptions of ANOVA and the appropriate transformation procedure was applied for violated parameters. Further, the approach of two-way analysis of variance (ANOVA) was applied for estimating the effect of genotypes and salinity, and their interaction and group comparison was made between tolerant and sensitive cultivars through contrast analysis using STAR statistical software [
25]. The relative contribution of different traits in total genetic divergence of the onion genotypes in control and salinity stress conditions were quantified through the method proposed by Singh [
26]. To find out the significant differences in various traits recorded in onion genotypes under salinity stress and control conditions, Duncan’s Multiple Range Test (DMRT) was performed using STAR statistical software [
25]. To predict the bulb yield under salinity stress, a response equation was derived through a stepwise regression approach, and significantly associated (
p > 0.005) morphological, physiological, and biochemical traits with bulb yield were also prioritized for conducting future studies on salinity stress tolerance in onions.
4. Discussion
To study the physiological and biochemical responses of thirty six onion accessions, we first evaluated them based on growth and bulb parameters. The results of the primary trial showed a significant reduction in plant height (1.07–12.91%), bulb diameter (0.37–25.85%), bulb yield per plant (14.88–62.86%), and total soluble solids (1.43–20%) under salinity stress in comparison to control (
Table S1). However, accessions differences were observed for reduction percentage in studied parameters. Based on percent bulb yield reduction, we selected seven salt-tolerant (less than 20% yield reduction) and seven-salt sensitive (more than 40% yield reduction) accessions to study the various physiological and biochemical changes in onion varieties as adaptive mechanisms. Plant height reduction in onions may be due to a decline in cell division and cell expansion under stress conditions, which ultimately reduces the overall plant growth [
27]. Moreover, refs. [
28,
29] reported that increased salt stress reduces leaf growth, number of leaves, number of branches, and stem diameter. Onions are a salt-sensitive crop and irrigation with saline water reduces the production of large sized bulbs, fresh weight, bulb firmness, water use efficiency, and bulb yield [
11]. Furthermore, a differential reduction rate in water dropwort cultivars for plant height, number of branches, number of leaves, and stem length was observed with increasing salt concentrations [
30]. Accumulation of excessive salt in leaves might be the probable cause of reduction in the leaf area of onions [
31,
32,
33].
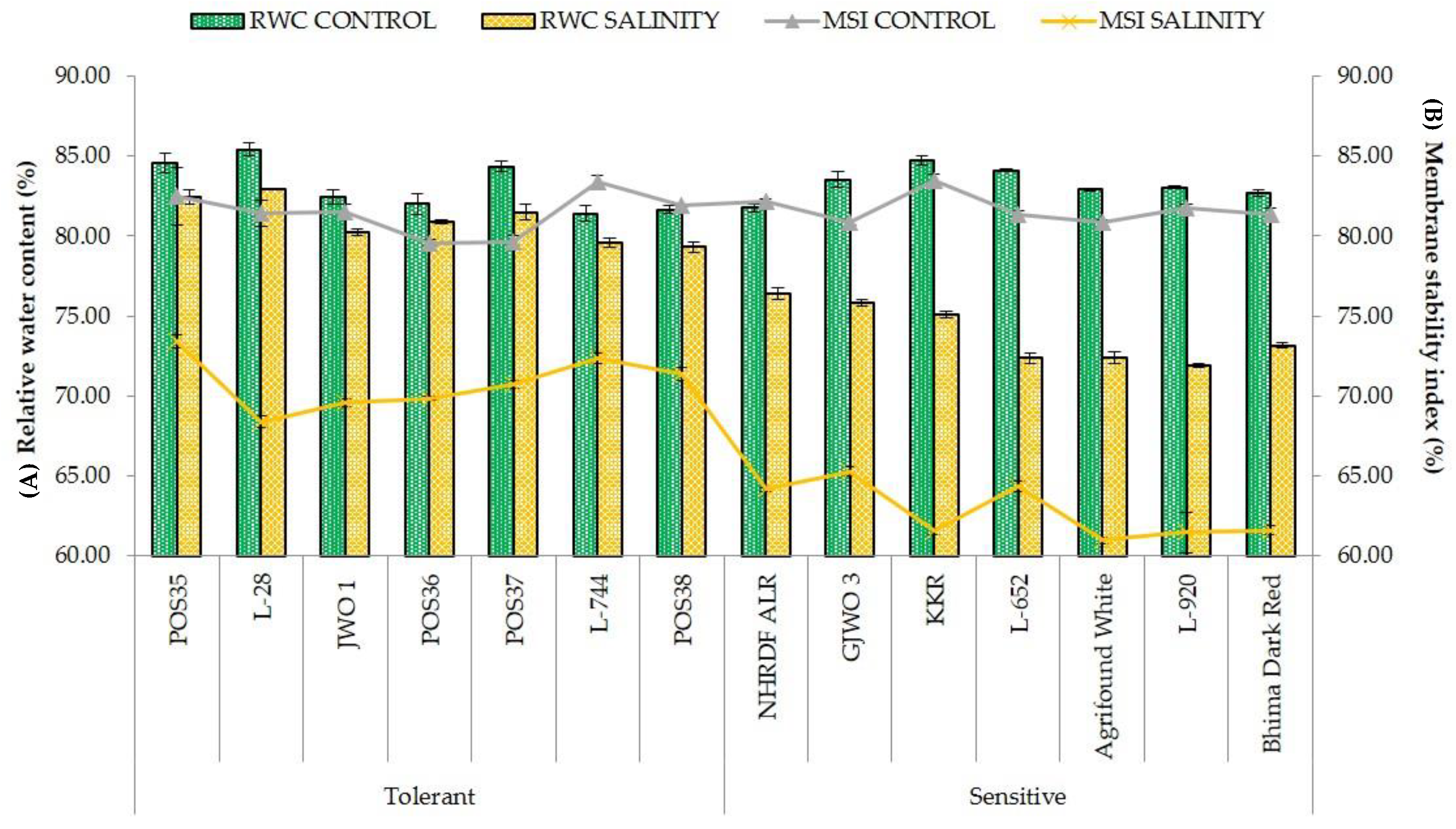
RWC and MSI are the most commonly used adaptive parameters for the selection of tolerant accessions under abiotic stress environments. Maintenance of high RWC and MSI are decisive indicators of high cellular turgidity and less cellular or membrane injuries in plants [
33]. Additionally, most of the physiological and biochemical processes such as cell division, cell enlargement, stomatal opening, and transportation in plants are dependent on the water status of the plant cells. Therefore, any reduction in water status adversely affects the plant’s metabolic functioning. Our results demonstrated that tolerant onion varieties showed high RWC and MSI rather than sensitive genotypes showing an agreement with the results of [
34,
35,
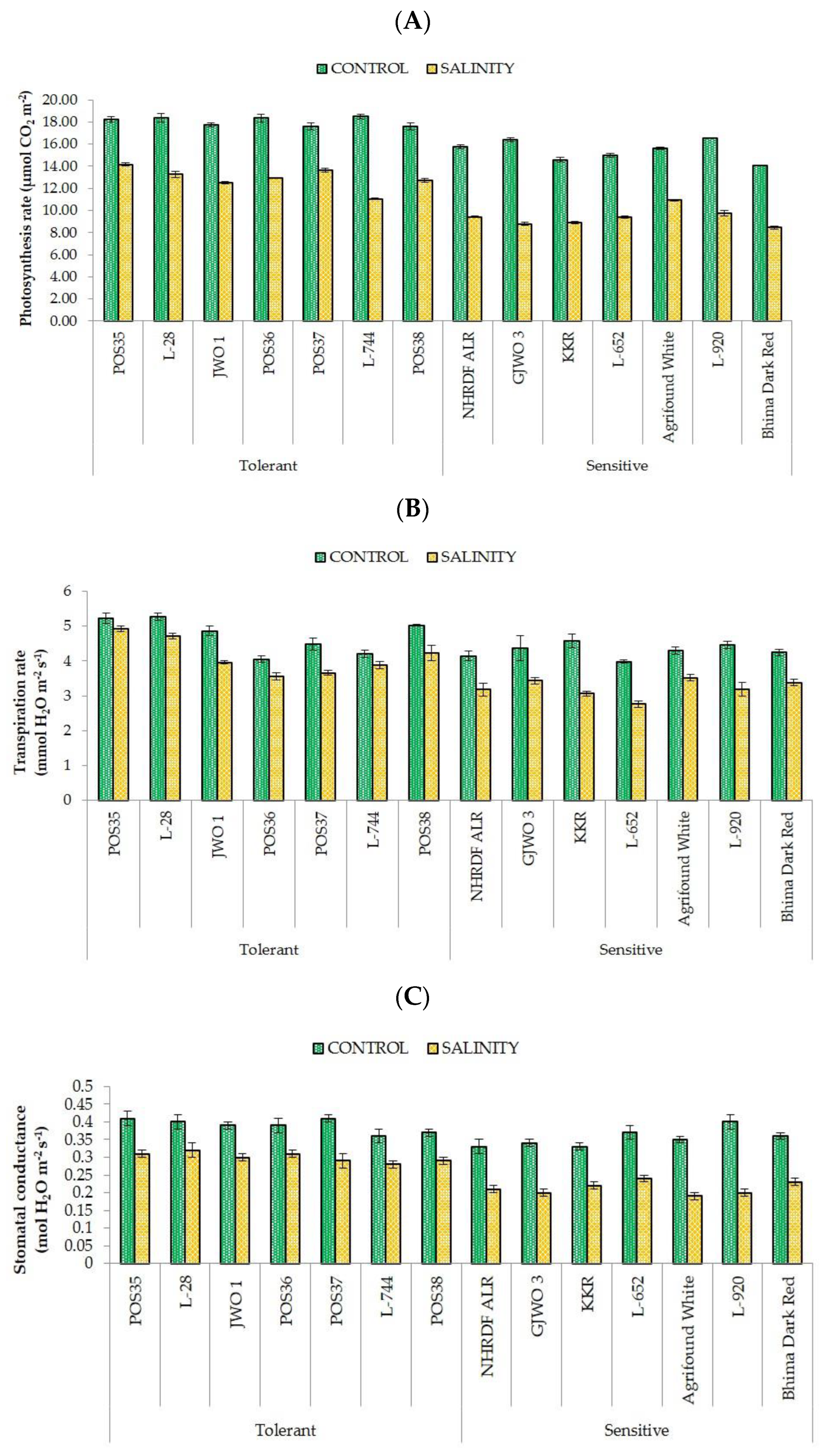
36]. Furthermore, the ability of plants to maintain a normal photosynthetic rate, stomatal conductance, and transpiration rate reflects the ability of salt tolerance. The obtained results in the present study showed a negative effect of salt stress on SPAD, Pn, gS, and E in all fourteen accessions. However, tolerant accessions showed a lesser reduction percentage compared to sensitive ones. These results are in agreement with the findings of earlier studies [
37,
38,
39] that reported a significant decrease in photosynthetic rate, transpiration rate, and chlorophyll content under salt stress. The decline in SPAD index under salinity may be due to photo-oxidation, loss of chloroplast membranes, and membrane injury in chloroplasts by ROS [
40,
41,
42]. However, in some studies, higher chlorophyll content was recorded in the saline environment, which may have been caused by the increase in number of chloroplasts [
30,
43]. Moreover, under excessive salinity, fewer gaseous exchanges occur in leaves because of the high sensitivity of stomata to abiotic stresses, leading to a reduction in transpiration and photosynthetic rates [
44,
45,
46,
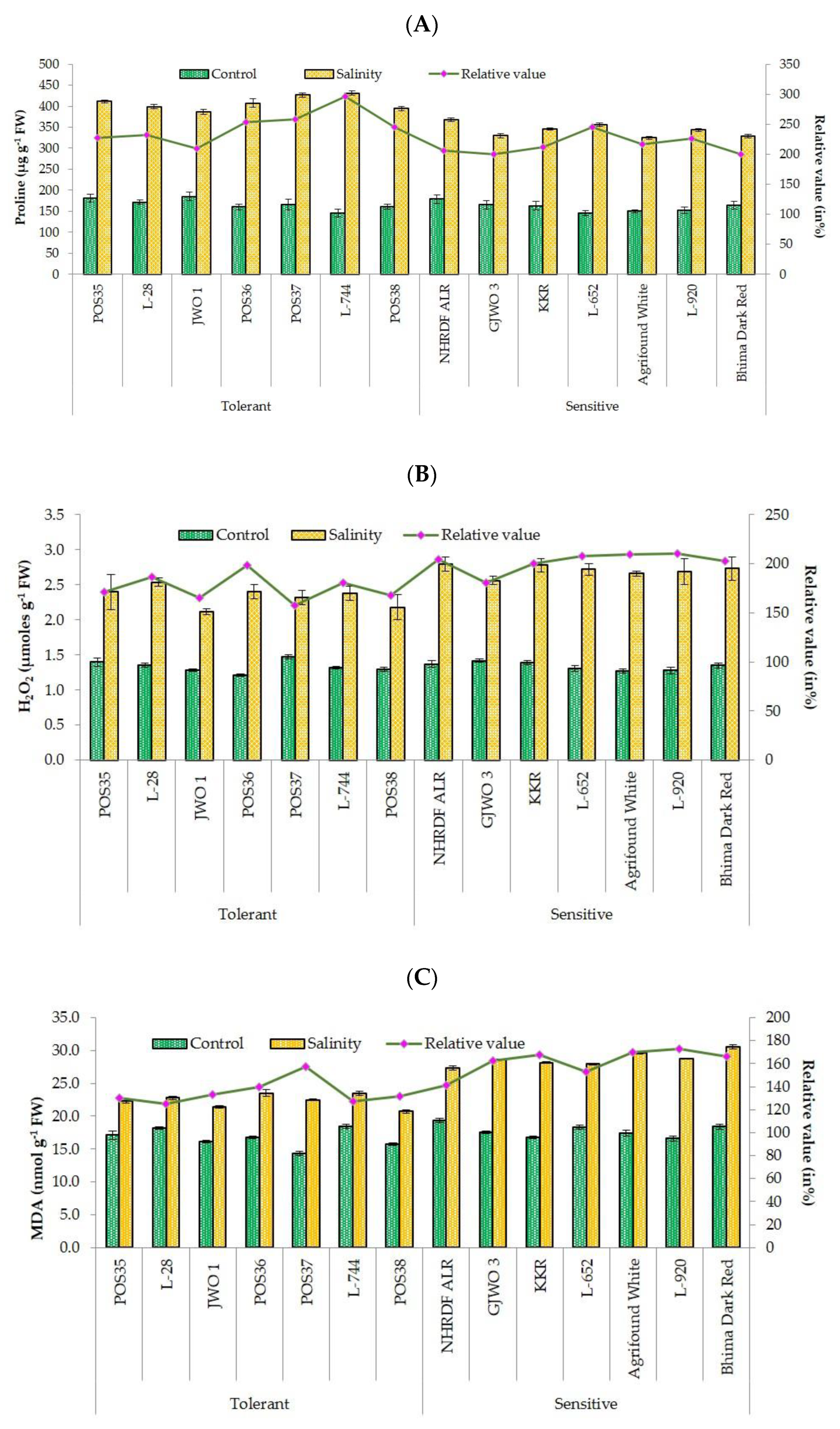
47]. Lipid peroxidation (MDA) and production of reactive oxygen species (H
2O
2) are commonly used biochemical indicators for plants exposed to abiotic stresses [
48]. The high concentration of MDA and H
2O
2 under salt stress leads to cell membrane damage and electrolyte leakage [
49]. In the present research, the accumulation of MDA and H
2O
2 was less in tolerant accessions compared to sensitive ones, indicating less oxidative damage to cell membranes of tolerant accessions. These results are also supported by other studies [
30,
50,
51], in which lower ROS production and lipid peroxidation and high membrane stability in salt-tolerant genotypes of rice, wheat, and water dropwort were reported. The compatible solute proline plays a very important role in maintaining the osmotic potential of cytosol with the external environment [
52]. The proline activity was observed as high under salt-stressed plants of tolerant accessions, whereas in sensitive accessions, the activity was comparatively lower. The enhanced activity of proline might be the important factor in neutralizing the negative effect of salinity stress via osmotic adjustment [
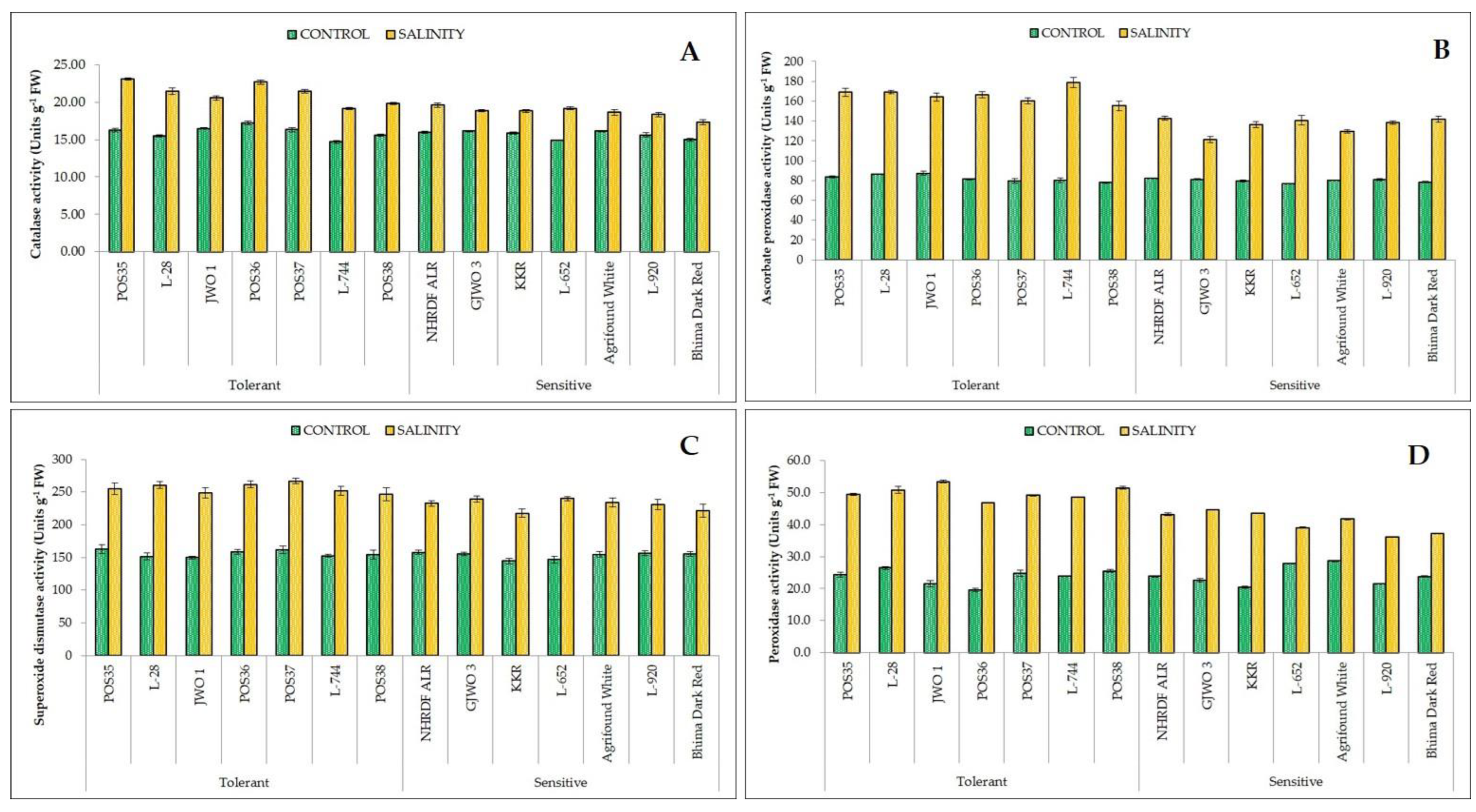
53]. Furthermore, a greater activity of antioxidants (SOD, APX, CAT, and POX) provides salt tolerance by scavenging ROS and reducing membrane damage [
54]. In defensive response, the SOD and CAT convert free radical species into H
2O
2 and oxygen, respectively [
55]. Salt tolerance in tomato, potato, cabbage, and amaranth caused by higher antioxidants activity were also reported [
48,
56,
57,
58,
59]. A positive correlation between enhanced antioxidant activity and reduced oxidative damage has been observed in the present study. The tolerant varieties under salt stress had better activity of these enzymes than those of the sensitive varieties.
Higher salt accumulation in the rhizosphere of plants leads to the accumulation of Na
+ and Cl
− ions, causing an osmotic effect, ionic imbalance, damage to enzymatic activities, and protein metabolism [
60]. Ionic imbalance causes a reduced uptake of essential nutrients such as potassium, manganese, and calcium to plant cells. The increased uptake of external Na
+ will enhance the Na
+ concentration in different plant organs with a concomitant decrease in K
+ [
12,
48,
61]. However, under high salinity stress, tolerant plants have a unique ability to accumulate and compartmentalize Na
+ and Cl
− in older leaves, but sensitive accessions cannot manage such compartmentalization, which leads to ionic and osmotic effects [
61]. This compartmentalization of Na
+ into vacuoles is controlled by Na
+/H
+ antiporters, V-type H
+-ATPase, and H
+-PPase [
62]. In this study, a higher Na
+/K
+ ratio was found in the roots, shoots, and bulbs, but percentage increase in the ratio was higher in sensitive accessions compared to tolerant accessions. Previous studies in carrot, amaranth, and pistachio also showed an increased uptake of Na
+ and decreased uptake of K
+ in salt-sensitive genotypes under salinity stress [
7,
51,
63]. Therefore, many authors suggested that maximal uptake of K
+ and minimal uptake of Na
+ is an indicator of salt tolerance in crop plants [
30,
51].
Stepwise regression analysis is a statistical method which identifies the most important contributing variable, signifying the amount of variability towards dependent variables such as economic yield. Based on this approach, we identified bulb diameter, plant height, ascorbate peroxidase, stomatal conductance, peroxidase, catalase, malondialdehyde, membrane stability index, and bulb Na
+/K
+ ratio as significantly contributing traits towards the total variation present in bulb yield per onion plant under salt stress. The significantly positive and negative coefficient of traits indicates that with increase or decrease of their respective value, there will be an increase or decrease in the final bulb yield per plant, respectively. In the previous research, Bojarian et al. [
64] also reported that single fruit weight, diameter, pericarp thickness, and titratable acidity are the important traits for successful breeding programs in tomatoes. Similarly, Saed-Moucheshi et al. [
65] selected spike weight and chlorophyll content as major traits for wheat breeding programs under different water regime conditions.
5. Conclusions
A wide range of variability was observed in studied onion genotypes for bulb yield and yield associated with morphological, physiological, and biochemical traits. The tolerant accessions selected based on percent yield reduction had strong antioxidant defense systems and lower Na+/K+ ratios in the shoot/leaves and maintained a higher tissue water status and osmoprotectants. Statistical analysis indicated that salinity stress significantly and negatively affects the bulb yield and yield-associated traits in onions, except number of leaves per plant. Interestingly, the salt-tolerant onion-tolerant cultivars showed a higher activity of antioxidant enzymes (APX, SOD, POX, and CAT), higher chlorophyll content (RWC, MSI, gS, and Pn), and lower production of H2O2 and lipid peroxidation compared to sensitive ones. Furthermore, the performance of onion cultivars assessed through traits modeling indicated a total of nine morphological, physiological, and biochemical traits (BD, PH, APX, gS, POX, CAT, MDA, MSI, and bulb Na+/K+ ratio), accounting for maximum variation in bulb yield under salinity stress, and which are the highly weighted variables that can be utilized for onion germplasm screening for salinity stress tolerance. From the experimental findings, we could summarize that the onion cultivars POS35, NHRDF Red (L-28), GWO 1, POS36, NHRDF Red-4 (L-744), POS37, and POS38 may be considered as saline-tolerant. These selected cultivars could be directly recommended for enhancing agricultural resilience in saline agro-ecosystems and can be utilized as potential genetic resources (salt-tolerant donor parents) in onion improvement programs.


 ,
, 





{kind=link}
{kind=link}
{kind=link}
{kind=link}