
Biomass Allocation and Leaf Morphology of Saplings Grown under Various Conditions of Light Availability and Competition Types


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
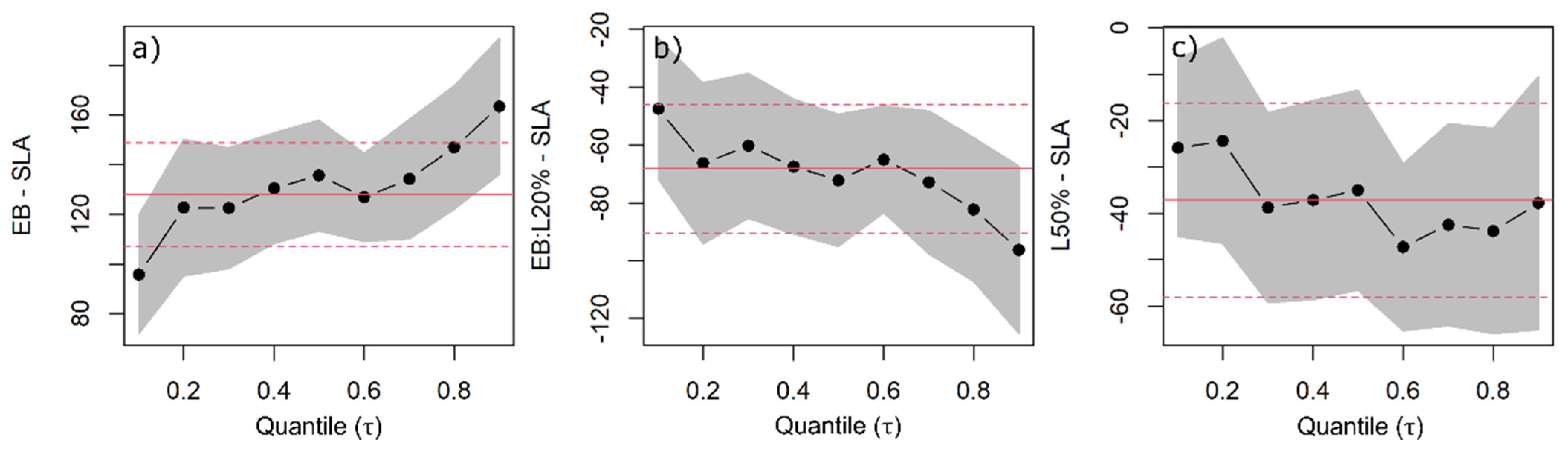
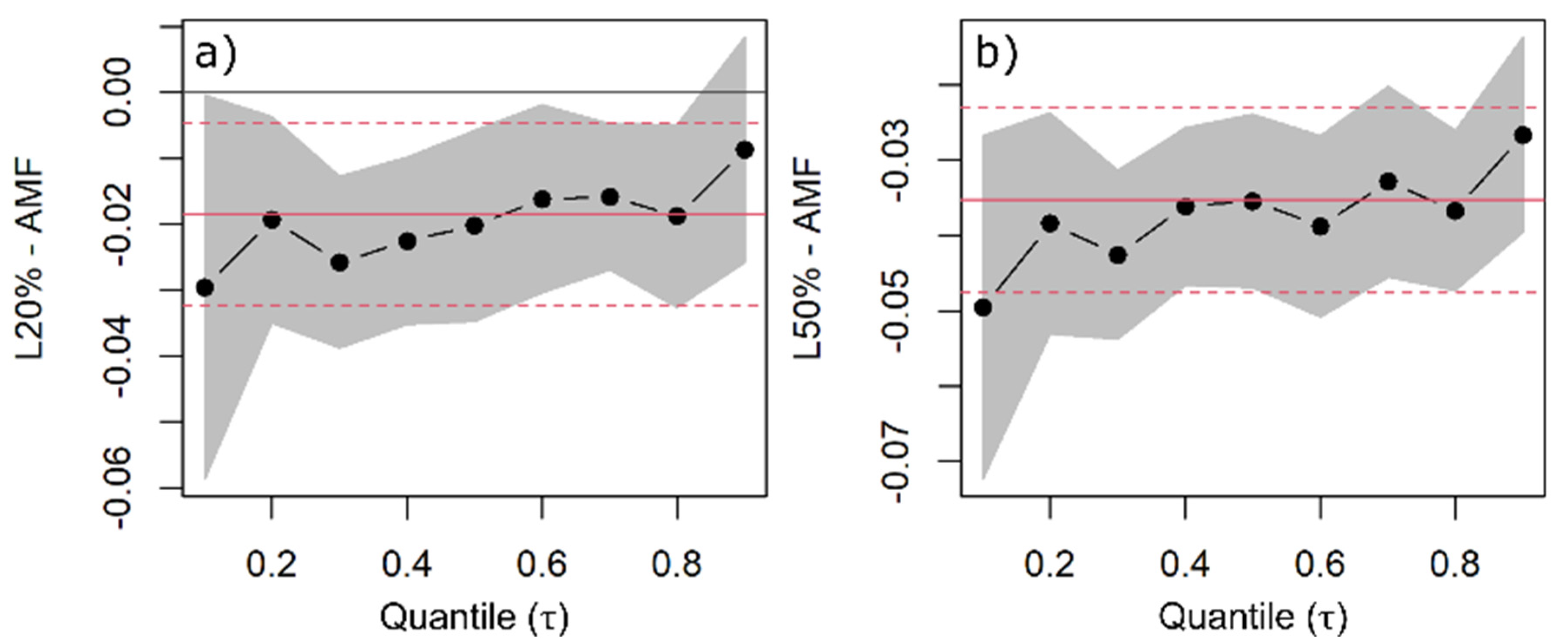
2.1. Specific Leaf Area and Leaf Mass Fraction
2.2. Woody Aboveground and Root Mass Fractions
2.3. Competitor’s Effect on Biomass Allocation
3. Discussion
3.1. Specific Leaf Area and Leaf Mass Fraction
3.2. Woody Aboveground and Root Mass Fractions
3.3. Competitor’s Effect on Biomass Allocation
4. Materials and Methods
4.1. Study Site and Experimental Setup
4.2. Sampling Procedure
4.3. Data Processing
4.4. Statistical Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Chapin, F.S.; Bloom, A.J.; Field, C.B.; Waring, R.H. Plant Responses to Multiple Environmental Factors. BioScience 1987, 37, 49–57. [Google Scholar] [CrossRef]
- Lambers, H.; Chapin, F.S.; Pons, T.L. Plant Physiological Ecology; Springer: New York, NY, USA, 2008; ISBN 978-0-387-78340-6. [Google Scholar]
- White, A.C.; Rogers, A.; Rees, M.; Osborne, C.P. How Can We Make Plants Grow Faster? A Source–Sink Perspective on Growth Rate. J. Exp. Bot. 2016, 67, 31–45. [Google Scholar] [CrossRef] [PubMed]
- Niklas, K.J. A Phyletic Perspective on the Allometry of Plant Biomass-partitioning Patterns and Functionally Equivalent Organ-categories. New Phytol. 2006, 171, 27–40. [Google Scholar] [CrossRef]
- Poorter, H.; Niklas, K.J.; Reich, P.B.; Oleksyn, J.; Poot, P.; Mommer, L. Biomass Allocation to Leaves, Stems and Roots: Meta-Analyses of Interspecific Variation and Environmental Control: Tansley Review. New Phytol. 2012, 193, 30–50. [Google Scholar] [CrossRef] [PubMed]
- Gedroc, J.J.; McConnaughay, K.D.M.; Coleman, J.S. Plasticity in Root/Shoot Partitioning: Optimal, Ontogenetic, or Both? Funct. Ecol. 1996, 10, 44. [Google Scholar] [CrossRef]
- Markovic, D.; Nikolic, N.; Glinwood, R.; Seisenbaeva, G.; Ninkovic, V. Plant Responses to Brief Touching: A Mechanism for Early Neighbour Detection? PLoS ONE 2016, 11, e0165742. [Google Scholar] [CrossRef] [Green Version]
- Valladares, F.; Gianoli, E.; Gómez, J.M. Ecological Limits to Plant Phenotypic Plasticity. New Phytol. 2007, 176, 749–763. [Google Scholar] [CrossRef]
- Pham, B.; McConnaughay, K. Environmental Selection and Convergent Evolution of Plant Phenotypes. In Ecology and the Environment; Springer: New York, NY, USA, 2013; pp. 1–20. ISBN 978-1-4614-7612-2. [Google Scholar]
- Messier, C.; Nikinmaa, E. Effects of Light Availability and Sapling Size on the Growth, Biomass Allocation, and Crown Morphology of Understory Sugar Maple, Yellow Birch, and Beech. Écoscience 2000, 7, 345–356. [Google Scholar] [CrossRef] [Green Version]
- Liu, Y.; Dawson, W.; Prati, D.; Haeuser, E.; Feng, Y.; van Kleunen, M. Does Greater Specific Leaf Area Plasticity Help Plants to Maintain a High Performance When Shaded? Ann. Bot. 2016, 118, 1329–1336. [Google Scholar] [CrossRef] [Green Version]
- Shipley, B.; Meziane, D. The Balanced-Growth Hypothesis and the Allometry of Leaf and Root Biomass Allocation. Funct. Ecol. 2002, 16, 326–331. [Google Scholar] [CrossRef]
- Bloom, A.J.; Chapin, F.S.; Mooney, H.A. Resource Limitation in Plants-An Economic Analogy. Annu. Rev. Ecol. Syst. 1985, 16, 363–392. [Google Scholar] [CrossRef]
- Lugli, L.F. Digging Deeper? Biomass Allocation Patterns in Trees and Lianas in Tropical Seasonal Forests. New Phytol. 2020, 226, 639–640. [Google Scholar] [CrossRef] [PubMed]
- Mccarthy, M.C.; Enquist, B.J. Consistency between an Allometric Approach and Optimal Partitioning Theory in Global Patterns of Plant Biomass Allocation. Funct. Ecol. 2007, 21, 713–720. [Google Scholar] [CrossRef]
- Annighöfer, P.; Petritan, A.M.; Petritan, I.C.; Ammer, C. Disentangling Juvenile Growth Strategies of Three Shade-Tolerant Temperate Forest Tree Species Responding to a Light Gradient. For. Ecol. Manag. 2017, 391, 115–126. [Google Scholar] [CrossRef]
- Delagrange, S.; Montpied, P.; Dreyer, E.; Messier, C.; Sinoquet, H. Does Shade Improve Light Interception Efficiency? A Comparison among Seedlings from Shade-Tolerant and -Intolerant Temperate Deciduous Tree Species. New Phytol. 2006, 172, 293–304. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Schall, P.; Lödige, C.; Beck, M.; Ammer, C. Biomass Allocation to Roots and Shoots Is More Sensitive to Shade and Drought in European Beech than in Norway Spruce Seedlings. For. Ecol. Manag. 2012, 266, 246–253. [Google Scholar] [CrossRef]
- Chen, H. Interspecific Responses of Planted Seedlings to Light Availability in Interior British Columbia: Survival, Growth, Allometric Patterns, and Specific Leaf Area. Can. J. For. Res. 1997, 27, 1383–1393. [Google Scholar] [CrossRef]
- Kunstler, G.; Curt, T.; Bouchaud, M.; Lepart, J. Growth, Mortality, and Morphological Response of European Beech and Downy Oak along a Light Gradient in Sub-Mediterranean Forest. Can. J. For. Res. 2005, 35, 1657–1668. [Google Scholar] [CrossRef]
- Montgomery, R. Relative Importance of Photosynthetic Physiology and Biomass Allocation for Tree Seedling Growth across a Broad Light Gradient. Tree Physiol. 2004, 24, 155–167. [Google Scholar] [CrossRef] [Green Version]
- Poorter, H.; Nagel, O. The Role of Biomass Allocation in the Growth Response of Plants to Different Levels of Light, CO2, Nutrients and Water: A Quantitative Review. Funct. Plant Biol. 2000, 27, 595. [Google Scholar] [CrossRef]
- Kobe, R.K.; Iyer, M.; Walters, M.B. Optimal Partitioning Theory Revisited: Nonstructural Carbohydrates Dominate Root Mass Responses to Nitrogen. Ecology 2010, 91, 166–179. [Google Scholar] [CrossRef]
- Gommers, C.M.M.; Visser, E.J.W.; Onge, K.R.S.; Voesenek, L.A.C.J.; Pierik, R. Shade Tolerance: When Growing Tall Is Not an Option. Trends Plant Sci. 2013, 18, 65–71. [Google Scholar] [CrossRef] [PubMed]
- Annighöfer, P. Stress Relief through Gap Creation? Growth Response of a Shade Tolerant Species (Fagus Sylvatica L.) to a Changed Light Environment. For. Ecol. Manag. 2018, 415–416, 139–147. [Google Scholar] [CrossRef]
- Zhang, L.; Copini, P.; Weemstra, M.; Sterck, F. Functional Ratios among Leaf, Xylem and Phloem Areas in Branches Change with Shade Tolerance, but Not with Local Light Conditions, across Temperate Tree Species. New Phytol. 2016, 209, 1566–1575. [Google Scholar] [CrossRef]
- Fang, Y.; Zou, X.; Lie, Z.; Xue, L. Variation in Organ Biomass with Changing Climate and Forest Characteristics across Chinese Forests. Forests 2018, 9, 521. [Google Scholar] [CrossRef] [Green Version]
- Bond, W.J. The Tortoise and the Hare: Ecology of Angiosperm Dominance and Gymnosperm Persistence. Biol. J. Linn. Soc. 1989, 36, 227–249. [Google Scholar] [CrossRef]
- Zhang, L.; Chen, Y.; Hao, G.; Ma, K.; Bongers, F.; Sterck, F.J. Conifer and Broadleaved Trees Differ in Branch Allometry but Maintain Similar Functional Balances. Tree Physiol. 2020, 40, 511–519. [Google Scholar] [CrossRef]
- Poorter, L. Growth Responses of 15 Rain–Forest Tree Species to a Light Gradient: The Relative Importance of Morphological and Physiological Traits. Funct. Ecol. 1999, 13, 396–410. [Google Scholar] [CrossRef]
- Canham, C.D.; Berkowitz, A.R.; Kelly, V.R.; Lovett, G.M.; Ollinger, S.V.; Schnurr, J. Biomass Allocation and Multiple Resource Limitation in Tree Seedlings. Can. J. For. Res. 1996, 26, 1521–1530. [Google Scholar] [CrossRef]
- Goldberg, D.E. 3-Components of Resource Competition in Plant Communities. In Perspectives on Plant Competition; Grace, J.B., Tilman, D., Eds.; Academic Press: Cambridge, MA, USA, 1990; pp. 27–49. ISBN 978-0-12-294452-9. [Google Scholar]
- Connell, J.H. 2-Apparent versus “Real” Competition in Plants. In Perspectives on Plant Competition; Grace, J.B., Tilman, D., Eds.; Academic Press: Cambridge, MA, USA, 1990; pp. 9–26. ISBN 978-0-12-294452-9. [Google Scholar]
- Ammer, C. Diversity and Forest Productivity in a Changing Climate. New Phytol. 2019, 221, 50–66. [Google Scholar] [CrossRef] [Green Version]
- del Río, M.; Condés, S.; Pretzsch, H. Analyzing Size-Symmetric vs. Size-Asymmetric and Intra- vs. Inter-Specific Competition in Beech (Fagus Sylvatica L.) Mixed Stands. For. Ecol. Manag. 2014, 325, 90–98. [Google Scholar] [CrossRef] [Green Version]
- Pretzsch, H.; Block, J.; Dieler, J.; Dong, P.H.; Kohnle, U.; Nagel, J.; Spellmann, H.; Zingg, A. Comparison between the Productivity of Pure and Mixed Stands of Norway Spruce and European Beech along an Ecological Gradient. Ann. For. Sci. 2010, 67, 712. [Google Scholar] [CrossRef] [Green Version]
- Pretzsch, H.; Schütze, G. Transgressive Overyielding in Mixed Compared with Pure Stands of Norway Spruce and European Beech in Central Europe: Evidence on Stand Level and Explanation on Individual Tree Level. Eur. J. For. Res. 2009, 128, 183–204. [Google Scholar] [CrossRef]
- Amoroso, M.M.; Turnblom, E.C. Comparing Productivity of Pure and Mixed Douglas-Fir and Western Hemlock Plantations in the Pacific Northwest. Can. J. For. Res. 2006, 36, 1484–1496. [Google Scholar] [CrossRef]
- Radosevich, S.R.; Hibbs, D.E.; Ghersa, C.M. Effects of Species Mixtures on Growth and Stand Development of Douglas-Fir and Red Alder. Can. J. For. Res. 2006, 36, 768–782. [Google Scholar] [CrossRef]
- Borrass, L.; Kleinschmit, D.; Winkel, G. The “German Model” of Integrative Multifunctional Forest Management—Analysing the Emergence and Political Evolution of a Forest Management Concept. For. Policy Econ. 2017, 77, 16–23. [Google Scholar] [CrossRef]
- Yamamoto, S.-I. Forest Gap Dynamics and Tree Regeneration. J. For. Res. 2000, 5, 223–229. [Google Scholar] [CrossRef]
- Spiecker, H.; Linder, M.; Schuler, J. Douglas-Fir—An Option for Europe; What Science Can Tell Us; European Forest Institute: Joensuu, Finland, 2019; ISBN 978-952-5980-65-3. [Google Scholar]
- Brunet, J.; Fritz, Ö.; Richnau, G. Biodiversity in European Beech Forests—A Review with Recommendations for Sustainable Forest Management. Ecol. Bull. 2010, 53, 77–94. [Google Scholar]
- Antúnez, I.; Retamosa, E.C.; Villar, R. Relative Growth Rate in Phylogenetically Related Deciduous and Evergreen Woody Species. Oecologia 2001, 128, 172–180. [Google Scholar] [CrossRef]
- Walters, M.B.; Kruger, E.L.; Reich, P.B. Growth, Biomass Distribution and CO2 Exchange of Northern Hardwood Seedlings in High and Low Light: Relationships with Successional Status and Shade Tolerance. Oecologia 1993, 94, 7–16. [Google Scholar] [CrossRef]
- Delucia, E.H.; Sipe, T.W.; Herrick, J.; Maherali, H. Sapling Biomass Allocation and Growth in the Understory of a Deciduous Hardwood Forest. Am. J. Bot. 1998, 85, 955–963. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Petriţan, A.M.; von Lüpke, B.; Petriţan, I.C. Influence of Light Availability on Growth, Leaf Morphology and Plant Architecture of Beech (Fagus Sylvatica L.), Maple (Acer Pseudoplatanus L.) and Ash (Fraxinus Excelsior L.) Saplings. Eur. J. For. Res. 2009, 128, 61–74. [Google Scholar] [CrossRef] [Green Version]
- Sevillano, I.; Short, I.; Grant, J.; O’Reilly, C. Effects of Light Availability on Morphology, Growth and Biomass Allocation of Fagus Sylvatica and Quercus Robur Seedlings. For. Ecol. Manag. 2016, 374, 11–19. [Google Scholar] [CrossRef] [Green Version]
- Valladares, F.; Niinemets, Ü. Shade Tolerance, a Key Plant Feature of Complex Nature and Consequences. Annu. Rev. Ecol. Evol. Syst. 2008, 39, 237–257. [Google Scholar] [CrossRef] [Green Version]
- Gruntman, M.; Groß, D.; Májeková, M.; Tielbörger, K. Decision-Making in Plants under Competition. Nat. Commun. 2017, 8, 2235. [Google Scholar] [CrossRef]
- Reich, P.B.; Tjoelker, M.G.; Walters, M.B.; Vanderklein, D.W.; Buschena, C. Close Association of RGR, Leaf and Root Morphology, Seed Mass and Shade Tolerance in Seedlings of Nine Boreal Tree Species Grown in High and Low Light. Funct. Ecol. 1998, 12, 327–338. [Google Scholar] [CrossRef]
- Aerts, R.; Boot, R.G.A.; van der Aart, P.J.M. The Relation between Above- and Belowground Biomass Allocation Patterns and Competitive Ability. Oecologia 1991, 87, 551–559. [Google Scholar] [CrossRef]
- Kawaletz, H.; Mölder, I.; Annighöfer, P.; Terwei, A.; Zerbe, S.; Ammer, C. Pot Experiments with Woody Species—A Review. Forestry 2014, 87, 482–491. [Google Scholar] [CrossRef] [Green Version]
- Kawaletz, H.; Mölder, I.; Annighöfer, P.; Terwei, A.; Zerbe, S.; Ammer, C. Back to the Roots: How Do Seedlings of Native Tree Species React to the Competition by Exotic Species? Ann. For. Sci. 2014, 71, 337–347. [Google Scholar] [CrossRef] [Green Version]
- Passioura, J.B. Viewpoint: The Perils of Pot Experiments. Funct. Plant Biol. 2006, 33, 1075. [Google Scholar] [CrossRef]
- Poorter, H.; Bühler, J.; van Dusschoten, D.; Climent, J.; Postma, J.A. Pot Size Matters: A Meta-Analysis of the Effects of Rooting Volume on Plant Growth. Funct. Plant Biol. 2012, 39, 839. [Google Scholar] [CrossRef] [Green Version]
- McConnaughay, K.D.M.; Coleman, J.S. Biomass Allocation in Plants: Ontogeny or Optimality? A Test along Three Resource Gradients. Ecology 1999, 80, 2581–2593. [Google Scholar] [CrossRef]
- Weiner, J. Allocation, Plasticity and Allometry in Plants. Perspect. Plant Ecol. Evol. Syst. 2004, 6, 207–215. [Google Scholar] [CrossRef]
- Annighöfer, P.; Seidel, D.; Ammer, C.; Stephens, S.L.; York, R.A. Silvicultural Implications from Analyzing Light Induced Height Growth Development of Eight North American Juvenile Tree Species in Mixed-Conifer Forests. Forestry 2019, 92, 616–626. [Google Scholar] [CrossRef]
- Delagrange, S.; Messier, C.; Lechowicz, M.J.; Dizengremel, P. Physiological, Morphological and Allocational Plasticity in Understory Deciduous Trees: Importance of Plant Size and Light Availability. Tree Physiol. 2004, 24, 775–784. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lödige, C.; Schall, P.; Ammer, C. How Do Size and Resource Availability Control Aboveground Biomass Allocation of Tree Seedlings? For. Res. Open Access 2014, 3, 123. [Google Scholar] [CrossRef]
- Cohen, J. A Power Primer. Psychol. Bull. 1992, 112, 155–159. [Google Scholar] [CrossRef] [PubMed]
- Nakagawa, S. A Farewell to Bonferroni: The Problems of Low Statistical Power and Publication Bias. Behav. Ecol. 2004, 15, 1044–1045. [Google Scholar] [CrossRef]
- Becker, P. Competition in the Regeneration Niche between Conifers and Angiosperms: Bond’s Slow Seedling Hypothesis. Funct. Ecol. 2000, 14, 401–412. [Google Scholar] [CrossRef]
- Dieler, J.; Pretzsch, H. Morphological Plasticity of European Beech (Fagus Sylvatica L.) in Pure and Mixed-Species Stands. For. Ecol. Manag. 2013, 295, 97–108. [Google Scholar] [CrossRef]
- Girard, F.; Vennetier, M.; Ouarmim, S.; Caraglio, Y.; Misson, L. Polycyclism, a Fundamental Tree Growth Process, Decline with Recent Climate Change: The Example of Pinus Halepensis Mill. in Mediterranean France. Trees 2011, 25, 311–322. [Google Scholar] [CrossRef]
- Parhizkar, P.; Sagheb-Talebi, K.; Mataji, A.; Nyland, R.; Namiranian, M. Silvicultural Characteristics of Oriental Beech (Fagus Orientalis Lipsky) Regeneration under Different RLI and Positions within Gaps. Forestry 2011, 84, 177–185. [Google Scholar] [CrossRef] [Green Version]
- Rajender Rao, K.; Acheré, V.; Bastien, C.; Bastien, J.-C.; Guérin, V.; Faivre-Rampant, P.; Favre, J.-M.; Jorge, V.; Leplé, J.-C.; Verger, M.; et al. Linkage Mapping and QTL Analysis for Growth of Young Pseudotsuga Menziesii Plants. In Sustainable Forestry, Wood products and Biotechnology, BIOFOR 02; DFA-AFA Press: Vitoria-Gasteiz, Spain, 2002; pp. 295–304. [Google Scholar]
- Longuetaud, F.; Caraglio, Y. Pith: A Marker of Primary Growth in Picea abies (L.) Karst. Trees 2009, 23, 325–334. [Google Scholar] [CrossRef]
- Barthélémy, D.; Caraglio, Y. Plant Architecture: A Dynamic, Multilevel and Comprehensive Approach to Plant Form, Structure and Ontogeny. Ann. Bot. 2007, 99, 375–407. [Google Scholar] [CrossRef] [Green Version]
- Goldberg, D.E. Competitive Ability: Definitions, Contingency and Correlated Traits. Philos. Trans. R. Soc. B Biol. Sci. 1996, 351, 1377–1385. [Google Scholar] [CrossRef]
- Henry, H.A.L.; Aarssen, L.W. The Interpretation of Stem Diameter-Height Allometry in Trees: Biomechanical Constraints, Neighbour Effects, or Biased Regressions? Ecol. Lett. 1999, 2, 89–97. [Google Scholar] [CrossRef]
- Bebre, I.; Riebl, H.; Annighöfer, P. Seedling Growth and Biomass Production under Different Light Availability Levels and Competition Types. Forests 2021, 12, 1376. [Google Scholar] [CrossRef]
- Welander, N.T.; Ottosson, B. The Influence of Shading on Growth and Morphology in Seedlings of Quercus robur L. and Fagus sylvatica L. For. Ecol. Manag. 1998, 107, 117–126. [Google Scholar] [CrossRef]
- Köppen, W. The Thermal Zones of the Earth According to the Duration of Hot, Moderate and Cold Periods and to the Impact of Heat on the Organic World. Meteorol. Z. 2011, 20, 351–360. [Google Scholar] [CrossRef]
- Gálhidy, L.; Mihók, B.; Hagyó, A.; Rajkai, K.; Standovár, T. Effects of Gap Size and Associated Changes in Light and Soil Moisture on the Understorey Vegetation of a Hungarian Beech Forest. Plant Ecol. 2006, 183, 133–145. [Google Scholar] [CrossRef]
- Walters, M.B.; Reich, P.B. Are Shade Tolerance, Survival, and Growth Linked? Low Light and Nitrogen Effects on Hardwood Seedlings. Ecology 1996, 77, 841–853. [Google Scholar] [CrossRef]
- Collet, C.; Chenost, C. Using Competition and Light Estimates to Predict Diameter and Height Growth of Naturally Regenerated Beech Seedlings Growing under Changing Canopy Conditions. Forestry 2006, 79, 489–502. [Google Scholar] [CrossRef] [Green Version]
- Bebre, I.; Riebl, H.; Annighöfer, P. Height, Diameter, Biomass, Leaf Area, and Relative Height Volume Growth of European Beech, Norway Spruce, and Douglas Fir Grown in Monospecific and Mixed Pots under Different Light Availability Levels. Pangaea 2021. [Google Scholar] [CrossRef]
- Fahrmeir, L.; Kneib, T.; Lang, S.; Marx, B. Regression; Springer: Berlin/Heidelberg, Germany, 2013; ISBN 978-3-642-34332-2. [Google Scholar]
- Cade, B.S.; Noon, B.R. A Gentle Introduction to Quantile Regression for Ecologists. Front. Ecol. Environ. 2003, 1, 412–420. [Google Scholar] [CrossRef]
- Scharf, F.S.; Juanes, F.; Sutherland, M. Inferring Ecological Relationships from the Edges of Scatter Diagrams: Comparison of Regression Techniques. Ecology 1998, 79, 448–460. [Google Scholar] [CrossRef]
- Yu, K.; Lu, Z.; Stander, J. Quantile Regression: Applications and Current Research Areas. J. R. Stat. Soc. D 2003, 52, 331–350. [Google Scholar] [CrossRef]
- Wooldridge, J.M. Introductory Econometrics: A Modern Approach, 5th ed.; South-Western Cengage Learning: Mason, OH, USA, 2013; ISBN 978-1-111-53104-1. [Google Scholar]
- R Core Team. R: A Language and Environment for Statistical Computing; R Foundation for Statistical Computing: Vienna, Austria, 2020. [Google Scholar]
- Koenker, R. Quantreg: Quantile Regression; R Foundation for Statistical Computing: Urbana, IL, USA, 2021. [Google Scholar]










Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Bebre, I.; Marques, I.; Annighöfer, P. Biomass Allocation and Leaf Morphology of Saplings Grown under Various Conditions of Light Availability and Competition Types. Plants 2022, 11, 305. https://0-doi-org.brum.beds.ac.uk/10.3390/plants11030305
Bebre I, Marques I, Annighöfer P. Biomass Allocation and Leaf Morphology of Saplings Grown under Various Conditions of Light Availability and Competition Types. Plants. 2022; 11(3):305. https://0-doi-org.brum.beds.ac.uk/10.3390/plants11030305
Chicago/Turabian StyleBebre, Ieva, Isa Marques, and Peter Annighöfer. 2022. "Biomass Allocation and Leaf Morphology of Saplings Grown under Various Conditions of Light Availability and Competition Types" Plants 11, no. 3: 305. https://0-doi-org.brum.beds.ac.uk/10.3390/plants11030305