1. Introduction
The genus
Tagetes, which includes plants known as ‘marigolds’, belongs to the Asteraceae family and contains more than 50 cultivated and wild species. Marigolds are native to America, but several species are naturalised in Africa, Asia, and Europe [
1,
2]. Plants of the genus
Tagetes are amongst the most widespread garden flowers worldwide. Marigolds are popular amongst gardeners due to their easy cultivation, wide adaptability, low demands for ecological and technological factors, and flower production throughout the year [
3].
As ornamental plants, marigolds have several advantages through which they have become particularly appreciated and widely cultivated in the world. These include: their suitability for a large type of cultivation, i.e., for bedding, edges, and pots, but also as cut flowers, bouquets, different floral arrangements, and other applications recognised in social or even religious life; long flowering period extending throughout the summer and fall; easy reproduction by seeds; and the great diversity of varieties within several species of
Tagetes [
3,
4,
5]. Furthermore, due to their multiple utilities, the short production period of decorative and marketable flowers, a broad spectrum of attractive colour palettes, shapes, and sizes of flowers, and also plant habits, marigolds have become one of the preferred species of many amateur and commercial flower growers [
6].
As a crop species, with a short period necessary for cultivation, marigolds can be grown in a multi-crop system, rotated with other crops, or as a mixed crop. In the latest case, cultivated on the borders with other plants can assure beneficial effects in organic agriculture management [
7], mainly in the production of important horticultural species, e.g., tomatoes [
4]. Due to their bactericidal, nematicidal, fungicide and insecticide action,
Tagetes species can be used in organic agriculture, especially in the culture of vegetables. Thiophenes, alkaloids, polyacetylenes, fatty acids, flavonoids and terpenes compounds present in
Tagetes confer antimicrobial and nematicidal effects, which may be of interest in organic farming. The use of natural or biological origin pesticides has the advantages of low toxicity, efficient control, and reduced cost. In addition, these products do not promote resistance to pests and diseases in agricultural crops [
7]. Highlighting the role and importance of
Tagetes plants in Mexican culture, Tapia-Vázquez et al. [
8] note their widespread use as an antioxidant, medicine, food pigment, flavouring, perfume, resin, ornamental and insecticide. Biocidal extracts from roots, stems, leaves, inflorescences, or from the whole plant allow wide use in agriculture as a nematicide, larvicide, attractant or insecticide and as a fertilizer [
4,
9].
Tagetes can be used in crop rotation and, in pest control, can be applied as organic manure to plants or as aqueous extracts and powders [
8]. The variation in
Tagetes cultivars belonging to different species allows the identification of genotypes with resistance or tolerance to saline and drought stresses and other adverse conditions [
4,
10,
11].
Marigolds are widely used for industrial and medicinal purposes and also as natural dyes and insecticides or herbicides [
4,
12,
13,
14,
15]. Due to their importance as medicinal plants in health care and herbal preparations, marigolds are cultivated and produced at an industrial scale, particularly in developing countries from the Far East. In addition, marigolds have received increasing attention in scientific research because of their multiple uses and their phytochemistry and bioactivity importance [
6,
16]. However, marigolds are extremely appreciated as annual ornamental plants,
Tagetes patula L.,
T. erecta L. and
T. tenuifolia Cav. being amongst the best known and most widespread. In addition to their value as ornamental plants, these three species have diverse and extremely numerous uses and properties, making them very interesting species to be used in marigold breeding works [
17].
The species
T. patula is native to the highlands of Mexico and Argentina but is now widespread throughout the world, known as the French marigold. The date of its introduction into Europe is uncertain, but it was first reported in France around 1880 [
18]. The species is also well acclimatised in Romania, where it is widely cultivated in gardens and green spaces. The plants of this species are small, about 40 cm, and have strong branches and elongated leaves (dark green), with pinnate compositions. Their floral heads are small, with thick peduncles, and the flowers are usually hermaphroditic, yellow, orange or a combination of colours. Flowering takes place gradually, starting in July and extending until September.
Tagetes erecta originates from Mexico and was first introduced to Europe in Spain in the 16th century [
19,
20]. It is known as the African marigold, Aztec marigold, American marigold, or big marigold. The species is well known in Romania. In Mexico, the species can reach up to 1.5 m or more, but the average of the most commonly used varieties is 60–80 cm [
21]. Excessive growth often complicates its growth in pots. For this reason, new commercial varieties have a height of only about 25–38 cm [
21]. The stems are glabrous or pubescent with green branches and numerous long ribs. The leaves are pinnately compounded. It blooms from July to October. The flower heads are large, with a diameter of 4–7 cm, comprising many individual flowers, usually orange in colour [
21].
Tagetes erecta has multiple uses as an ornamental plant in decorating green spaces and various floral arrangements and garlands; it is planted in crops as an insect repellent and has use in pharmacy and medicine, industry, and so forth [
22]. African marigold meal and extracts are used in poultry feed to colour the skin, meat (fat) and egg yolks and as colourants in a wide variety of food products [
23]. Yellow-orange flowers are sources of great interest in the extraction of natural oils and pigments. Lutein is extracted from flowers, the main pigment used to produce the colour orange,
T. erecta being the main source of this pigment for commercial uses [
20]. The species can be used as natural air filters or sinks to alleviate air pollution [
24]; it also has a strong invasive potential [
20] and, from cultivated gardens, can easily migrate into spontaneous flora and spread freely, Romania being recorded as being invaded by the alien flora [
25].
The origin of
T. tenuifolia is found in America, and it occurs from Mexico throughout Central America to central Colombia [
26]; compared to the two previous species, it is less common. It is known as signet marigold, golden marigold or lemon marigold. It has smaller dark green compound leaves and smaller, yellow, orange, golden or multicoloured floral heads. It gradually blooms from June to October [
27,
28].
Marigold seeds are usually sown directly in the field [
29]. The optimal sowing period varies depending on the geographical area. In Romania, this period corresponds to mid-April. Seed germination is influenced by various environmental factors, especially soil temperature and humidity, as well as watering frequency and water quality used. Seed quality is paramount in marigolds to achieve rapid and uniform seedling emergence, avoid gaps in the field and ensure uniform field culture [
30].
There are different studies of morphological and genetic diversity within and among species of
Tagetes. The knowledge of morphological and decorative characters or useful biochemical components and their genetic variability could support new breeding programmes for different species of interest [
31,
32,
33,
34,
35] as well as for marigold [
36]. Genetic variation and relationships among genotypes are essential in classifying and utilising germplasm resources in marigold breeding [
37]. Using RAPD markers in the fingerprinting of marigold, 22 cultivars of
T. patula were considered a suitable reservoir of useful genes that could be utilized for interspecific and intervarietal crosses to obtain variation, which can be exploited to create new valuable ornamental cultivars [
38]. In another study, both RAPD and STS markers were successful in estimating the genetic diversity of
Tagetes species and genotypes (
T. patula,
T. erecta,
T. minuta) and in selecting parents for breeding and mapping purposes [
39]. SSR markers also proved effective for genetic diversity analysis, species classification and individual identification in commercial cultivars and landraces of
T. patula and
T. erecta [
40]. The trends in modern plant breeding programmes may have contributed to the genetic vulnerability of all cultivated species of
Tagetes. Moreover, the genetic background of new cultivars in marigolds must be diversified to reduce the risk or possible crises in the future and minimise vulnerability. New hybrids possessing an intense heterosis effect, with superior ornamental traits and market values, can be produced using interspecific hybridisation, e.g., between African marigolds and French marigolds [
41,
42]. In this way, the success of new breeding programmes in
Tagetes depends on the genetic variation of the germplasm as well as on its efficient use [
43].
Based on the above considerations, this study aims to characterise several cultivars of T. erecta, T. patula and T. tenuifolia. Information on seed characteristics was analysed in relation to germination capacity and seedling appearance. Another question addressed in this study was to assess how these characteristics affect the growth and vegetative development of plants and their reproductive efficiency (number of flowers and set of seeds). The analysis of the plants’ phenophases of growth and fruiting and the main traits of the plants was performed comparatively both inside the species (within the tested cultivars) and between the species. Furthermore, the genetic variation and phylogenetic relations of the cultivars were investigated with SSR markers.
2. Results
2.1. Seed Characteristics
Both for the main characteristics of the seeds and the germination of the seeds, there were obvious differences between the analysed varieties (
Table 1). In
T. patula, the seed length ranged between 8.8 mm (Bolero) and 14.0 mm (Orion). The trait variation amongst the nine cultivars of the species was very different, with values of the coefficient of variation between 1.1% (Queen Sophia) and 22.3% (Bolero). Significant differences between cultivars were also recorded for the width and thickness of the seeds. The best germination capacity of the seeds was recorded in the Orange Flame, Robuszta and Orion cultivars (all with 100% germinated seeds). Instead, the seeds of the cultivar Mr. Majestik had low germination (43.5%). In the five cultivars of
T. tenuifolia, the amplitude of variation for seed traits was much lower (e.g., between 6.6–8.8 mm for seeds’ length), as was the germination of seeds (this varied between 87.0% in Sarga and 100.0% in Luna Gold and Luna Lemon), although significant differences between cultivars were detected. In
T. erecta, the seed length ranged between 10.9 and 15.1 mm, these extreme values being obtained on Aranysarga and Antigua Orange cultivars, respectively. For all seed traits, there were significant differences between cultivars. Seed germination was 100% in a single cultivar (Magas Citromsarga) but low in Alacsony Citromsarga (65.2%) and extremely low in Arctic (8.7%).
The variation of traits of seeds within the varieties was generally small (CV% below 10%) in a few cases, registering values of CV% that indicated an intermediate (CV% between 10.1–20%) or high (CV% over 20.1%) variation. The last category included Bolero for seed length (CV% = 22.3) and Orange Flame, with CV% just over 20%, for seed width and thickness.
Boxplots and Duncan multiple range tests (
Figure 1) highlight the differences between the three species of
Tagetes for the three main traits of the seeds. Between
T. patula and
T. erecta, there were no significant differences between the means of seed length, width and thickness. In contrast, the mean values of these traits in
T. tenuifolia were significantly lower than those of the first two species mentioned. Only for seed germination,
T. tenuifolia does not differ significantly from the other two species,
T. patula and
T. erecta. The graphs also suggestively indicate the dispersion of data within and between species. For the characteristics of the seeds, the whiskers of the boxplots suggest a large dispersion of the cultivar values in
T. patula. Outliers or abnormal values, marked by small dots, appear only in
T. erecta for seed thickness. Instead, such aberrant values are highlighted by boxplots for seed germination in both
T. patula and
T. erecta.
2.2. Plant Growth Traits
Significant differences between varieties within species were detected for plant height, stem diameter number of branches per stem, and angle of branches (
Table 2).
The amplitude of the variation for plant height ranged between 25.4 and 47.8 cm for T. patula, 21.0 and 21.8 cm for T. tenuifolia, and 24.4 and 69.8 cm for T. erecta. The extreme mean values of the stem diameter were comprised between 8.5 and 12.9 mm (T. patula), 8.7 and 9.2 mm (T. tenuifolia), and 8.9 and 15.8 mm (T. erecta). The branching of the plants was very different, the average number of branches on the stem being between the following values: 7.6 and 13.7 (T. patula), 16.0 and 16.9 (T. tenuifolia), and 8.9 and 14.2 (T. erecta). Finally, the fluctuation of the average values for the insertion angle of the main branches oscillated between the following limits: 40.0 and 60.6 degrees (T. patula), 51.5 and 52.8 degrees (T. tenuifolia), and 37.3 and 71.1 degrees (T. erecta).
The coefficients of variation of these traits generally had values appreciated as small (CV% below 10.1%) or even very small, i.e., for the angle of branch insertion, for all three species.
The boxplots with plant growth (
Figure 2) evidenced a higher plant height of
T. erecta, with a mean of cultivars close to 60 cm. Duncan’s multiple range tests indicated that the mean of the analysed cultivars of this species is significantly higher than the means of cultivars of
T. patula and
T. tenuifolia. In addition, it can be seen that the mean plant height of
T. erecta is reduced by a value that the boxplot highlights as an outlier. From
Table 1, it can be found that the respective cultivar is Antigua Orange, with an average plant height of only 24.4 cm, much lower compared to the other cultivars of
T. erecta species. An opposite situation occurred in
T. patula, in which an outlier (Queen Sophia cv., with 47.8 cm) raised the mean of the species.
For all the plant growth and branching characteristics, the boxplots indicate a fairly high homogeneity of T. tenuifolia cultivars. Of the three species, T. erecta stands out for its significantly higher differences in stem diameter and T. tenuifolia for the number of branches per stem. For the angle of branches insertion, there were no real differences within the three species.
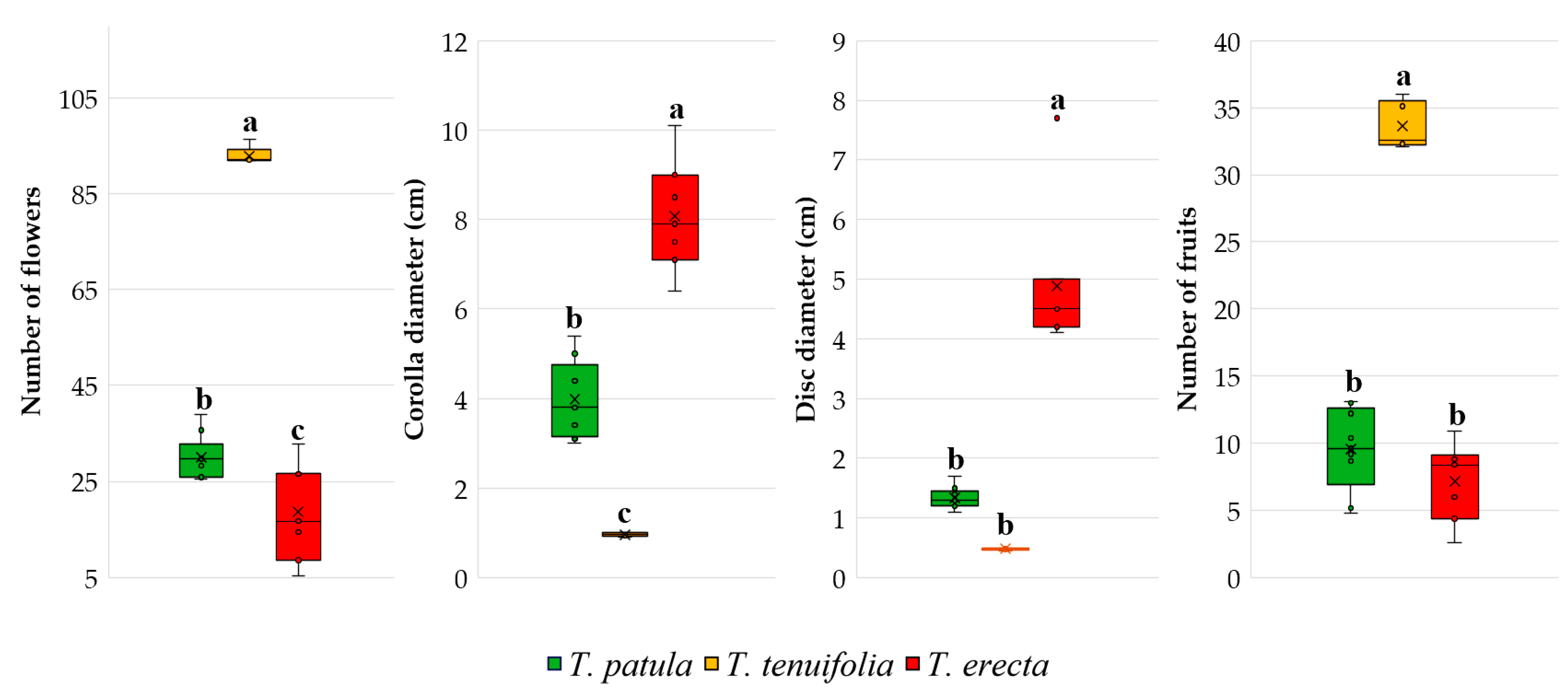
2.3. Main Traits Related to Flowers and Fruits
For the traits analysed and presented in
Table 3 (number of flowers per plant, corolla diameter, disc diameter, number of fruits), only in the case of disc diameter in
T. tenuifolia species there were no differences between cultivars within the same species. A higher number of flowers per plant was recorded in
T. tenuifolia (between 91.9 and 96.4), but a greater variation in trait per species was noted in
T. erecta (5.3 and 32.7). The corolla diameter was small and uniform in the varieties belonging to the species
T. tenuifolia, of medium size in
T. patula (with an amplitude of varieties between 3.0 and 5.4 cm) and large in
T. erecta (with limits between 6.4 and 10.1 cm). Disc diameter of the flowers was small and uniform in the cultivars of
T. tenuifolia and
T. patula, but large compared to
T. erecta cultivars. The numbers of fruits (achenes) in the heads oscillated, depending on the species:
T. patula (4.8 and 13.1);
T. tenuifolia (32.1 and 36.0);
T. erecta (2.6 and 10.9). For this last trait, the most significant variability was registered especially in the cultivars Robuszta, Cresto Yellow and Bolero (all of the
T. patula species, with CV% over 20); Hawaii, Arctic, Aranysarga, and Antigua Orange (all of the
T. erecta species; the first two with CV% over 30, and the next two over 20).
Amongst the main phenological phases (
Table 4), seedling emergence showed a higher amplitude of the number of days required in
T. patula (10.0–27.0), followed by
T. erecta (14.0–24.0). A smaller variation was recorded for
T. tenuifolia cultivars (13.0–15.0). Stem formation required a more variable number of days to
T. patula (19–30 days) and
T. erecta (29–39 days) cultivars, whereas in
T. tenuifolia cultivars, this phase unfolded more homogeneous (22–25 days). The cultivars of
T. tenuifolia also had the following vegetative phenophases (branching, budding) and were more uniform in their progress compared to
T. patula and
T. erecta. The cultivars of
T. tenuifolia showed a great uniformity for developing the main generative phenophases: full flowering, flowering duration, fruit formation, and fruit ripening, respectively. Probably, this particularity was due to the group of cultivars from the ‘Luna’ range (four cultivars), with close origin.
The long duration of the number of days required from sowing to reach full flowering in some cultivars of T. patula was noted, as well as the variation of this trait between cultivars (41–100 days). Even in T. erecta (70–83 days), the number of days to reach full flowering was much higher than in T. tenuifolia (45–47 days).
Flowering duration, reported as an average of the number of days a flower maintains its decorative features at an appropriate level, was relatively close for the three species; the highest amplitude was obtained in T. patula cultivars (15–26 days). Regarding fruit formation and fruit ripening, the longest period required, but also the highest amplitude of the two characters, was recorded in T. patula (between 54–118 days for fruit formation and between 67–145 days for fruit ripening).
The species
T. tenuifolia differs from the other two species for the main traits of the flowers and the number of fruits (
Figure 3). Only for the diameter of the floral disc,
T. tenuifolia does not differ significantly from
T. patula. The boxplots and Duncan’s multiple range tests also highlight the high number of flowers and fruits in
T. tenuifolia compared to the other two species and the extremely narrow variability of flower traits recorded in
T. tenuifolia.
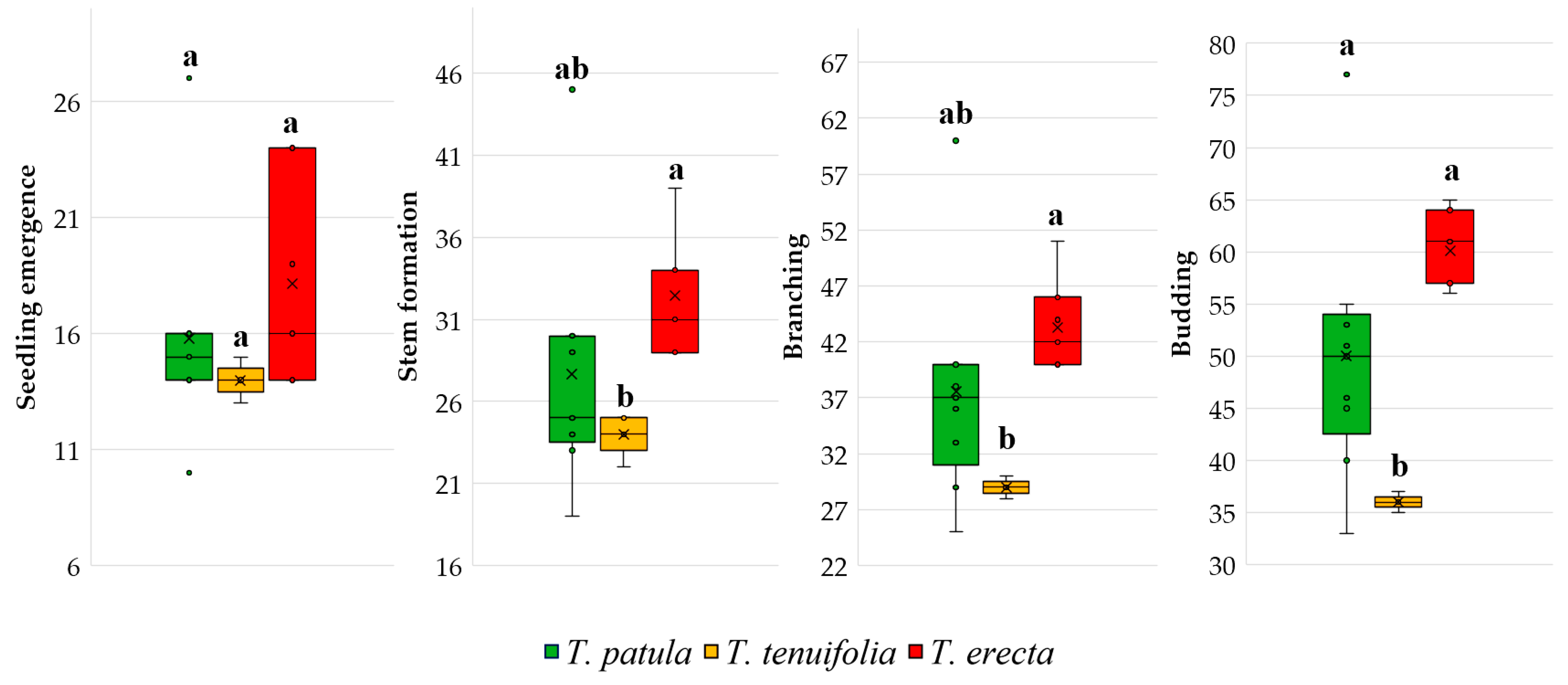
In
Figure 4, the range and distribution of the area for the vegetative phenophases of the three species of
Tagetes are presented. There is a smaller variability for
T. tenuifolia for all traits, whereas a more significant variability is highlighted for seedling emergence of
T. erecta. In addition, the boxplots emphasise the apparition of some larger outliers of
T. patula, visible for all four traits. Duncan’s multiple range tests showed that there were no significant differences between the three species for seedling emergence; instead, such differences were recorded for stem formation, branching and budding.
Regarding the main generative phenophases, the lowest variability was also recorded in the
T. tenuifolia species (
Figure 5). Even if there were no significant differences between the three species for the duration of flowering, unlike
T. tenuifolia, in which the cultivars showed a remarkable uniformity of flowering period, in
T. patula and
T. erecta cultivars, the flowering period was longer. Boxplots suggestively differentiate the dispersal of flowering days in the last two species, respectively, by the trait variation, depending on the cultivars’ genotypes.
2.4. Phenotypic Correlations between the Analyzed Traits
By calculating the phenotypic correlation coefficients between the analysed traits, numerous positive or negative relations, statistically significant as
p < 0.05, 0.01 and 0.001 (
Table 5), were highlighted between different traits of seeds, plants, flowers, or the vegetative and reproductive phenophases, fruit formation or seed ripening, respectively.
Thus, significant positive correlations were detected amongst seed traits (length, width, and thickness). In addition, these were positively correlated with corolla diameter and disc diameter, stem formation, branching, budding, and also with full flowering, fruit formation and fruit ripening. On the contrary, the three seed traits were negatively correlated with the number of branches per stem, the number of flowers, and the number of fruits.
Amongst some elements of plant vigour, such as plant height and stem diameter, there were positive correlations with decorative elements of flowers (i.e., corolla diameter, disc diameter, full flowering), or even with fruit formation and fruit ripening. However, there were also some inversely proportional relations (negative correlations) with the number of flowers and fruits. The emergence of seedlings was closely correlated with stem formation, branching, budding, full flowering, flowering duration, fruit formation, and fruit ripening. Generally, significant, positive correlations were identified amongst all these characters.
It is interesting that between a trait of great ornamental interest, such as the number of flowers, significant negative correlations were registered, some of them unexpected, with numerous analysed traits, for example, the morphological particularities of the seeds (length, width and thickness), plants (height and stem diameter), flowers (corolla diameter and disc diameter), or vegetative and reproductive phenophases (stem formation, branching, budding, full flowering, fruit formation and fruit ripening). Contrary to these negative correlations, the number of flowers was positively correlated with the number of branches on the plant and the number of fruits. The correlations registered between the number of flowers and different traits analysed were quite close also for the number of fruits (as the direction, positive or negative, and significance of correlations). Unlike the number of flowers, the corolla diameter and the disc diameter were positively correlated with most of the traits, except for the number of flowers and fruits.
2.5. Multivariate Analysis of Traits (Principal Component Analysis) and Genetic Variation
The principal component analysis (PCA) plot provides an interesting visual projection and essential information of the relationships between the three species of
Tagetes based on the multivariate variation of the 20 analysed traits (
Figure 6a). The PCA illustrates a compact grouping of variables, which are grouped together and appear as positively correlated (i.e., fruit ripening, fruit formation, budding, full flowering, branching—in quadrant I; seed width, seed length, seed thickness, stem diameter, plant height, corolla diameter, disc diameter—quadrant II).
Also closely grouped and positively correlated are the number of flowers and fruits and the number of branches per stem, all situated in quadrant IV. Their opposite positioning (and negative correlations) can be seen compared to the traits in quadrant II, including the branching angle of the main branches on the plant. PCA confirms the relations identified with Pearson correlations and may suggest that the fastigiate growth of bushes and branches with a basitone tendency is in a relatively inverse proportion to the abundant flowering and fruiting capacity.
The PCA of the 21 cultivars (
Figure 6b) highlights that the cultivars of
T. tenuifolia are very close to each other and form a particularly homogeneous group, contrasting with some cultivars of
T. patula (e.g., Bolero, Cresto Flame, Cresto Yellow) or
T. erecta (Alacsony Citromsarga). Some of the
T. erecta cultivars are located quite compactly in quadrant I, but two are included in quadrant II, one even at a great distance (Arctic). In the
T. patula species, most cultivars are grouped close to the centre, respectively, to the origin of the plot. This could indicate that they have average properties, but this assertion is contradicted by the opposite positioning towards the extremities of the two opposite quadrants of Mr. Majestic and Orion cultivars.
The first and second components of the PCA accounted for 62.19% and 20.28%, respectively, of the total variation observed.
Hierarchical clustering analyses illustrated a homogeneous grouping of cultivars within each species of
Tagetes (
Figure 7). Only one exception was recorded for the Orion and Mr. Majestic cultivars, belonging to
T. patula, which were situated on two different clusters, clearly spaced from the compact group of the other species cultivars (these being placed on a common subcluster).
Based on the phenotypic traits and phenological attributes conferring general peculiarities of greater ornamental and landscape importance, 13 cultivars from the three species were subjected to molecular analysis. The SSR technique was considered optimal for phylogenetic analysis and for testing the genetic diversity among cultivars.
In order to assess the genetic diversity present amongst the different
Tagetes genotypes, a set of six SSRs developed by Whankaew et al. [
40] were selected, taking into account the number of alleles detected and the PIC (
Table 6). A total of 13 alleles were detected using six SSR combinations with a mean of 3.5 alleles per locus. Although the expected heterozygosity has a mean of 0.46, the observed heterozygosity was very low in general, ranging from 0.00 to 0.23. This indicates a high degree of homozygosity, probably due to the various floral mechanisms adapted in Asteraceae to reduce the likelihood of self-pollination within co-sexual flowers [
44]. The PIC (polymorphic information content) value, which assesses the informativeness of the markers in a population, ranged from 0.18 in the marker TE11, which was much lower than the others, to 0.66 in the marker TE59, with a general mean of 0.40.
Similar results were observed by Whankaew et al. [
40], analysing the genetic diversity of 40 individuals of two different
Tagetes species (
T. erecta and
T. patula). This work detected a mean of three alleles per locus, with an expected heterozygosity average value of 0.48, and the markers had an average PIC of 0.48. The observed heterozygosity was higher (0.32) than that observed here, most likely due to the inclusion of commercial hybrids in their study.
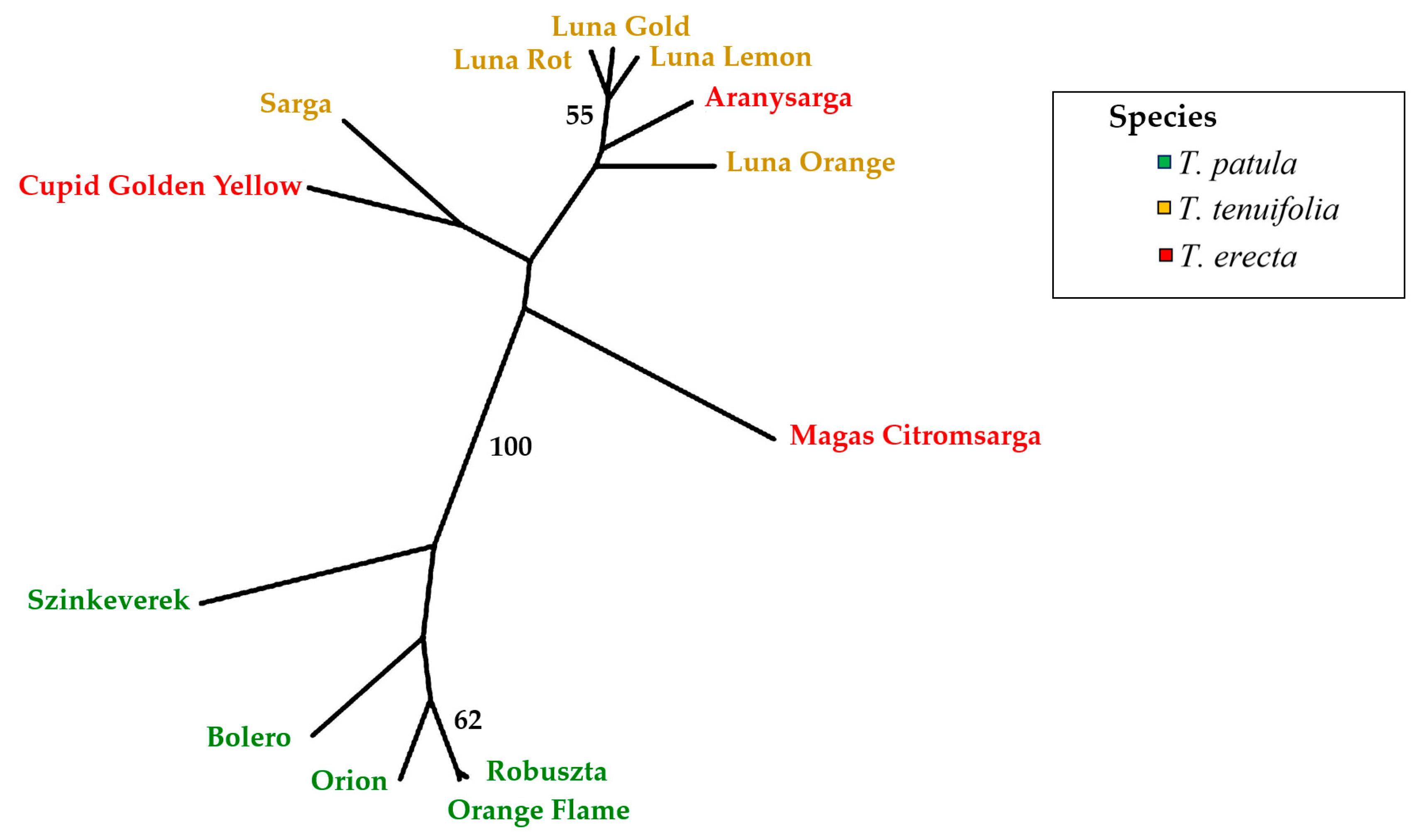
The UPGMA dendrogram using Nei’s genetic distance [
45] distributed the three species into two main clusters (
Figure 8) based on 1000 bootstrap randomisations. The first cluster separates all
T. patula cultivars from the rest with a bootstrap value of 100%. The second cluster is composed of the rest of the individuals belonging to the other two species. No clear separation by species was observed in this cluster, although all ‘Luna’ cultivars (
T. tenuifolia) clustered together with the
T. erecta cultivar Aranysarga. The pairwise genetic distance values (
Table S1) confirm the results obtained with the UPGMA dendrogram. In this way, genetic distances within
T. patula were generally lower than the genetic distances with cultivars from the two other species, and
T. tenuifolia ‘Luna’ cultivars also displayed low genetic distances among them.
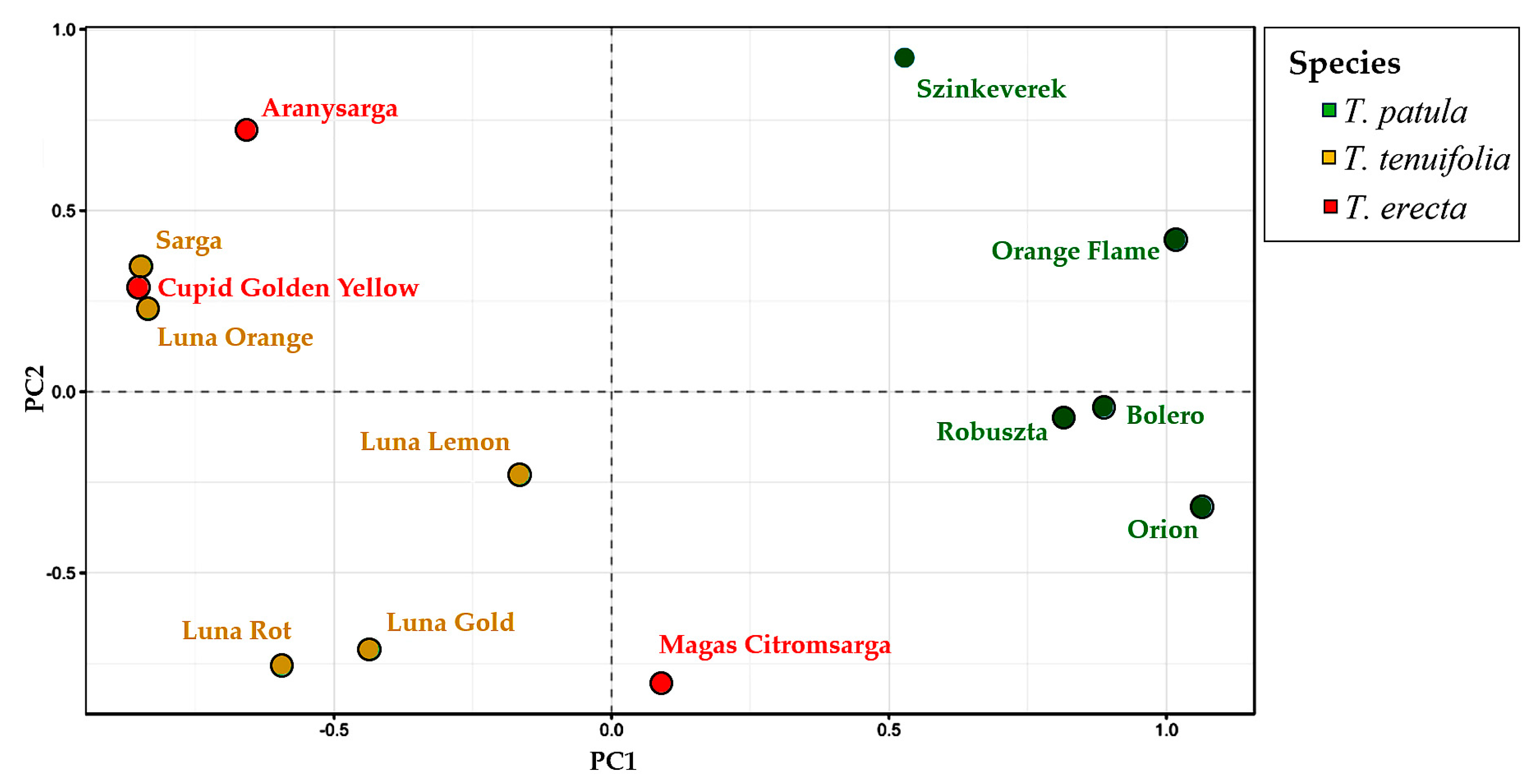
A multivariate principal coordinate analysis (PCoA) confirmed the grouping of genotypes based on UPGMA. The first two axes explain 51.35% of the genetic variation. The first component explains 33.75% of the variation (
Figure 9) and clearly separate
T. patula from
T. erecta and
T. tenuifolia, which are spread in the same area of the PCoA plot. The second component explains 17.61% of the total variation and allows a better separation of the accessions within the species. The groups observed agree with the ones detected in the UPGMA dendrogram. Among
T. patula individuals, Szinkeverek is the most genetically different. The three individuals of
T. erecta are very dispersed, indicating a wide genetic diversity. The individuals of
T. tenuifolia can be separated into two subgroups (
Figure 9). The first one is composed by Sarga and Luna Orange, and the second includes Luna Lemon, Luna Root and Luna Gold.



 ,
, 













{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}