
Changes in Carbon and Nitrogen Metabolites before, at, and after Anthesis for Wheat Cultivars in Response to Reduced Soil Water and Zinc Foliar Application



Abstract
:1. Introduction
2. Results
2.1. Variances
2.1.1. Soluble Sugars
2.1.2. Soluble Proteins
2.1.3. Free Amino Acids
2.2. Means of Zn Levels
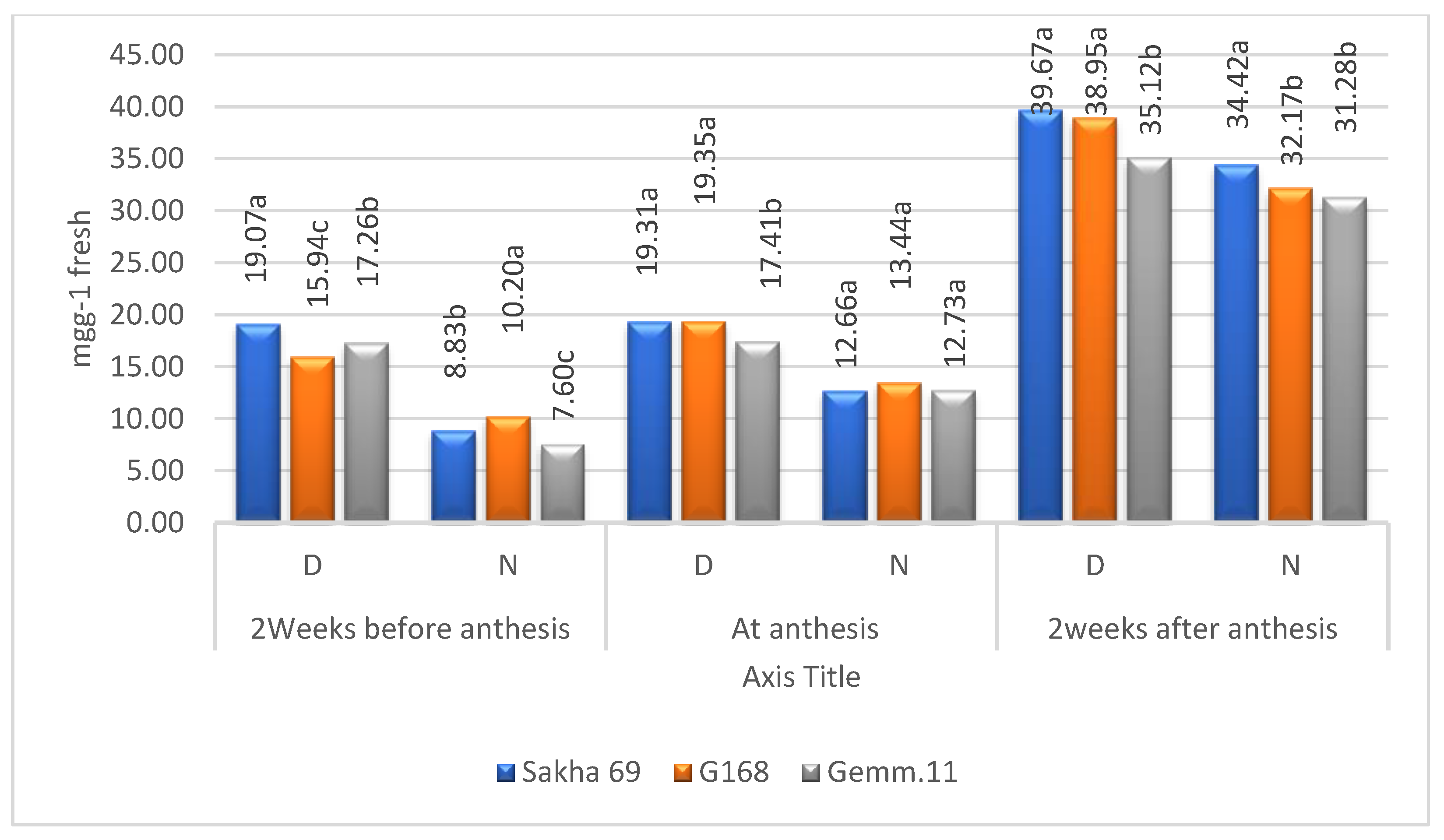
2.3. Means of Cultivars
3. Correlation between Different Metabolites in Wheat Cultivars under the Effect of Various Zinc Concentrations
4. Discussion
5. Materials and Methods
5.1. The Genetic Materials
5.2. Irrigation
5.3. Fertilization
5.4. Preparation of Plant Extracts for Analysis
5.5. Determination of Water-Soluble Metabolites
5.6. Statistical Analysis
6. Conclusions
Author Contributions
Funding
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Sehgal, A.; Sita, K.; Kumar, J.; Kumar, S.; Singh, S.; Siddique, K.H.M.; Nayyar, H. Effects of Drought, Heat and Their Interaction on the Growth, Yield and Photosynthetic Function of Lentil (Lens Culinaris Medikus) Genotypes Varying in Heat and Drought Sensitivity. Front. Plant Sci. 2017, 8, 1776. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sehgal, A.; Sita, K.; Siddique, K.H.M.; Kumar, R.; Bhogireddy, S.; Varshney, R.K.; HanumanthaRao, B.; Nair, R.M.; Prasad, P.V.V.; Nayyar, H. Drought or/and Heat-Stress Effects on Seed Filling in Food Crops: Impacts on Functional Biochemistry, Seed Yields, and Nutritional Quality. Front. Plant Sci. 2018, 9, 1705. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Szegletes, Z.; Erdei, L.; Tari, I.; Cseuz, L. Accumulation of Osmoprotectants in Wheat Cultivars of Different Drought Tolerance. Cereal Res. Commun. 2000, 28, 403–410. [Google Scholar] [CrossRef]
- Yordanov, I.; Velikova, V.; Tsonev, T. Plant Responses to Drought, Acclimation, and Stress Tolerance. Photosynthetica 2000, 38, 171–186. [Google Scholar] [CrossRef]
- Marček, T.; Hamow, K.Á.; Végh, B.; Janda, T.; Darko, E. Metabolic Response to Drought in Six Winter Wheat Genotypes. PLoS ONE 2019, 14, e0212411. [Google Scholar] [CrossRef] [Green Version]
- Abid, M.; Ali, S.; Qi, L.K.; Zahoor, R.; Tian, Z.; Jiang, D.; Snider, J.L.; Dai, T. Physiological and Biochemical Changes during Drought and Recovery Periods at Tillering and Jointing Stages in Wheat (Triticum Aestivum L.). Sci. Rep. 2018, 8, 4615. [Google Scholar] [CrossRef]
- Dietz, K.-J.; Zörb, C.; Geilfus, C.-M. Drought and Crop Yield. Plant Biol. 2021, 23, 881–893. [Google Scholar] [CrossRef]
- Blum, A. Osmotic Adjustment Is a Prime Drought Stress Adaptive Engine in Support of Plant Production. Plant Cell Environ. 2017, 40, 4–10. [Google Scholar] [CrossRef]
- Keyvan, S. The Effects of Drought Stress on Yield, Relative Water Content, Proline, Soluble Carbohydrates and Chlorophyll of Bread Wheat Cultivars. J. Anim. Plant Sci. 2010, 8, 1051–1060. [Google Scholar]
- Sarker, A.M.; Rahman, M.S.; Paul, N.K. Effect of Soil Moisture on Relative Leaf Water Content, Chlorophyll, Proline and Sugar Accumulation in Wheat. J. Agron. Crop Sci. 1999, 183, 225–229. [Google Scholar] [CrossRef]
- Kunjadia, K.R. and S.H. and B.A. Effect of Zinc and Iron Application on Leaf Chlorophyll, Carotenoid, Grain Yield and Quality of Wheat in Calcareous Soil of Saurashtra Region. Int. J. Chem. Stud. 2018, 6, 2092–2096. [Google Scholar]
- Husnain Gondal, A.; Farooq, Q.; Sohail, S.; Shasang Kumar, S.; Danish Toor, M.; Zafar, A.; Rehman, B. Adaptability of Soil PH through Innovative Microbial Approach. Curr. Res. Agric. Sci. 2021, 8, 71–79. [Google Scholar] [CrossRef]
- Gondal, A.H. A Detailed Review Study of Zinc Involvement in Animal, Plant and Human Nutrition. Indian J. Pure Appl. Biosci. 2021, 9, 262–271. [Google Scholar] [CrossRef]
- Umair Hassan, M.; Aamer, M.; Umer Chattha, M.; Haiying, T.; Shahzad, B.; Barbanti, L.; Nawaz, M.; Rasheed, A.; Afzal, A.; Liu, Y.; et al. The Critical Role of Zinc in Plants Facing the Drought Stress. Agriculture 2020, 10, 396. [Google Scholar] [CrossRef]
- Tayyiba, L.; Zafar, H.; Gondal, A.H.; Farooq, Q.; Mukhtar, M.M.; Hussain, R.; Aslam, N.; Muzaffar, A.; Sattar, I. Efficiency of Zinc in Plants, Its Deficiency and Sensitivity for Different Crops. Curr. Res. Agric. Sci. 2021, 8, 128–134. [Google Scholar] [CrossRef]
- Thabet, S.G.; Alqudah, A.M. Crops and Drought. In eLS; Wiley: Hoboken, NJ, USA, 2019; pp. 1–8. ISBN 9780470015902. [Google Scholar]
- Castillo, R.R.; Lozano, D.; González, B.; Manzano, M.; Izquierdo-Barba, I.; Vallet-Regí, M. Advances in Mesoporous Silica Nanoparticles for Targeted Stimuli-Responsive Drug Delivery: An Update. Expert Opin. Drug Deliv. 2019, 16, 415–439. [Google Scholar] [CrossRef]
- Ali, A.; Bhat, B.A.; Rather, G.A.; Malla, B.A.; Ganie, S.A. Proteomic Studies of Micronutrient Deficiency and Toxicity. In Plant Micronutrients; Springer International Publishing: Cham, Switzerland, 2020; pp. 257–284. [Google Scholar]
- El-Dahshouri, M.F.; El-Fouly, M.M.; Khalifa, R.K.M.; El-Ghany, H.M.A. Effect of Zinc Foliar Application at Different Physiological Growth Stages on Yield and Quality of Wheat under Sandy Soil Conditions. Agric. Eng. Int. CIGR J. 2017, 2017, 193–200. [Google Scholar]
- Sarker, U.; Oba, S. Catalase, Superoxide Dismutase and Ascorbate-Glutathione Cycle Enzymes Confer Drought Tolerance of Amaranthus Tricolor. Sci. Rep. 2018, 8, 16496. [Google Scholar] [CrossRef] [Green Version]
- Ma, D.; Sun, D.; Wang, C.; Ding, H.; Qin, H.; Hou, J.; Huang, X.; Xie, Y.; Guo, T. Physiological Responses and Yield of Wheat Plants in Zinc-Mediated Alleviation of Drought Stress. Front. Plant Sci. 2017, 8, 860. [Google Scholar] [CrossRef] [Green Version]
- Sun, L.; Song, F.; Guo, J.; Zhu, X.; Liu, S.; Liu, F.; Li, X. Nano-ZnO-Induced Drought Tolerance Is Associated with Melatonin Synthesis and Metabolism in Maize. Int. J. Mol. Sci. 2020, 21, 782. [Google Scholar] [CrossRef] [Green Version]
- Dubois, M.; Gilles, K.; Hamilton, J.K.; Rebers, P.A.; Smith, F. A Colorimetric Method for the Determination of Sugars. Nature 1951, 168, 167. [Google Scholar] [CrossRef] [PubMed]
- Sattar, A.; Wang, X.; Abbas, T.; Sher, A.; Ijaz, M.; Ul-Allah, S.; Irfan, M.; Butt, M.; Wahid, M.A.; Cheema, M.; et al. Combined Application of Zinc and Silicon Alleviates Terminal Drought Stress in Wheat by Triggering Morpho-Physiological and Antioxidants Defense Mechanisms. PLoS ONE 2021, 16, e0256984. [Google Scholar] [CrossRef] [PubMed]
- Yadav, A.K.; Carroll, A.J.; Estavillo, G.M.; Rebetzke, G.J.; Pogson, B.J. Wheat Drought Tolerance in the Field Is Predicted by Amino Acid Responses to Glasshouse-Imposed Drought. J. Exp. Bot. 2019, 70, 4931–4948. [Google Scholar] [CrossRef] [PubMed]
- Hammad, S.A.R.; Ali, O.A.M. Physiological and Biochemical Studies on Drought Tolerance of Wheat Plants by Application of Amino Acids and Yeast Extract. Ann. Agric. Sci. 2014, 59, 133–145. [Google Scholar] [CrossRef] [Green Version]
- Parida, A.K.; Dagaonkar, V.S.; Phalak, M.S.; Umalkar, G.V.; Aurangabadkar, L.P. Alterations in Photosynthetic Pigments, Protein and Osmotic Components in Cotton Genotypes Subjected to Short-Term Drought Stress Followed by Recovery. Plant Biotechnol. Rep. 2007, 1, 37–48. [Google Scholar] [CrossRef]
- Sankar, B.; Jaleel, C.A.; Annan, P.A.; Kishorekumar, A.; Somasundaram, R.; Vam, R.P. Drought-Induced Biochemical Modifications and Proline Metabolism in Abelmoschus Esculentus (L.) Moench. Acta Bot. Croat. 2007, 66, 43–56. [Google Scholar]
- El-Sharkawi, H.M.; Farghali, K.A.; Sayed, S.A. Interactive Effects of Water Stress, Temperature and Nutrients in the Seed Germination of Three Desert Plants. J. Arid Environ. 1989, 17, 307–317. [Google Scholar] [CrossRef]
- Lee, Y.P.; Takahashi, T. An Improved Colorimetric Determination of Amino Acids with the Use of Ninhydrin. Anal. Biochem. 1966, 14, 71–77. [Google Scholar] [CrossRef]
- Lowry, O.; Rosebrough, N.; Farr, A.L.; Randall, R. PROTEIN MEASUREMENT WITH THE FOLIN PHENOL REAGENT. J. Biol. Chem. 1951, 193, 265–275. [Google Scholar] [CrossRef]
- David, F.N.; Steel, R.G.D.; Torrie, J.H. Principles and Procedures of Statistics. Biometrika 1961, 48, 234. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Item | Soluble Sugars under Drought | Soluble Proteins under Drought | Free Amino Acids under Drought | ||||||
| Year 1 | |||||||||
| S.V. | 2wb | Anthesis | 2wa | 2wb | Anthesis | 2wa | 2wb | Anthesis | 2wa |
| Reps | 11.30 | 1.050 | 6.230 | 6.290 * | 49.58 | 15.00 | 1.130 | 4.640 | 0.600 |
| Zn levels (A) | 240.5 ** | 413.8 ** | 24.14 | 21.61 ** | 81.41 ** | 57.09 * | 2.110 | 41.39 ** | 13.96 ** |
| Error A | 10.27 | 28.53 | 58.96 | 2.150 | 8.360 | 12.60 | 0.670 | 0.810 | 1.110 |
| Cultivars (B) | 128.3 ** | 384.1 ** | 297.4 ** | 12.77 * | 29.42 | 158.3 ** | 2.690 | 0.150 | 4.940 ** |
| A × B | 61.45 ** | 181.3 * | 539.6 ** | 33.16 ** | 10.46 ** | 130.3 ** | 3.100 | 12.63 ** | 3.710 ** |
| Error B | 8.400 | 61.14 | 40.36 | 2.260 | 8.420 | 13.70 | 1.470 | 1.110 | 0.790 |
| Year 2 | |||||||||
| Reps | 12.74 | 25.36 | 0.170 | 6.920 | 3.730 | 1.830 | 0.041 | 0.580 | 0.690 |
| Zn levels (A) | 12.74 ** | 200.7 ** | 49.30 ** | 145.0 ** | 40.30 ** | 300.3 * | 0.520 ** | 2.460 * | 3.280 ** |
| Error A | 0.860 | 0.080 | 2.860 | 3.000 | 3.240 | 53.39 | 0.055 | 0.400 | 0.140 |
| Cultivars (B) | 35.85 ** | 1.22 | 15.69 ** | 94.27 ** | 4.180 | 105.6 ** | 0.570 | 1.260 * | 1.350 |
| A × B | 5.850 * | 69.15 ** | 23.73 ** | 72.51 ** | 47.08 ** | 62.01 * | 1.920 ** | 2.990 ** | 1.990 * |
| Error B | 1.880 | 4.150 | 1.370 | 0.760 | 3.700 | 15.14 | 0.330 | 0.290 | 0.560 |
| Combined | |||||||||
| Years (Y) | 1340 ** | 1874 ** | 1802 ** | 229.7 ** | 1203 ** | 42.96 ** | 363.7 ** | 197.9 ** | 47.57 ** |
| Reps/years | 7.260 | 13.21 | 3.200 | 6.610 | 26.66 | 8.410 | 0.590 | 2.610 | 0.640 |
| Zn levels (A) | 154.1 ** | 74.98 * | 68.54 | 128.5 ** | 66.53 ** | 60.45 | 1.440 * | 30.95 ** | 4.240 ** |
| A × Y | 99.14 ** | 539.5 ** | 4.890 | 38.08 ** | 55.18 ** | 297.0 ** | 1.200 | 12.90 ** | 13.00 ** |
| Error A | 5.56 | 14.3 | 30.91 | 2.580 | 5.800 | 32.99 | 0.360 | 0.610 | 0.630 |
| Cultivars (B) | 27.51 ** | 213.2 ** | 199.0 ** | 59.21 ** | 29.54 | 143.8 ** | 2.870 | 0.590 | 0.590 |
| B × Y | 136.6 ** | 172.1 * | 114.1 ** | 47.83 ** | 5.060 | 120.2 ** | 0.380 | 0.810 | 5.710 ** |
| A × B | 47.33 ** | 82.03 * | 363.88 ** | 45.44 ** | 105.0 ** | 118.42 ** | 3.85 ** | 11.30 ** | 5.350 ** |
| A × B × Y | 19.97 ** | 168.4 ** | 199.5 ** | 60.23 ** | 46.66 ** | 73.93 ** | 1.170 | 4.320 ** | 0.350 |
| Error B | 5.140 | 32.64 | 20.86 | 1.510 | 6.060 | 14.42 | 0.890 | 0.700 | 0.680 |
| Soluble sugars under irrigation | Soluble proteins under irrigation | Free amino acids under irrigation | |||||||
| Year 1 | |||||||||
| S.V. | 2wb | Anthesis | 2waa | 2wb | Anthesis | 2wa | 2wb | Anthesis | 2w a |
| Reps | 1.190 | 12.84 | 18.88 | 2.550 | 1.590 | 20.94 | 1.240 | 0.500 | 0.220 |
| Zn levels (A) | 13.67 | 946.6 ** | 823.2 ** | 32.87 ** | 250.0 ** | 447.75 ** | 18.00 ** | 5.800 | 3.400 ** |
| Error A | 2.890 | 23.04 | 9.090 | 2.690 | 2.300 | 8.950 | 1.420 | 1.460 | 0.290 |
| Cultivars (B) | 29.30 ** | 1226.4 ** | 344.0 ** | 88.67 ** | 7.980 | 65.79 * | 5.510 * | 21.75 ** | 11.96 ** |
| A × B | 11.11 ** | 1656 ** | 342.4 ** | 87.82 ** | 37.10 ** | 485.9 ** | 2.930 | 19.62 ** | 4.500 ** |
| Error B | 0.820 | 50.25 | 7.840 | 3.560 | 5.800 | 13.58 | 1.450 | 0.620 | 0.820 |
| Year 2 | |||||||||
| Reps | 0.230 | 0.040 | 0.300 | 0.180 | 0.290 | 2.980 | 1.560 | 0.430 | 1.410 |
| Zn levels (A) | 53.67 ** | 44.46 ** | 67.70 ** | 0.660 ** | 15.65 ** | 14.74 | 13.10 * | 9.680 | 2.370 |
| Error A | 0.930 | 1.610 | 0.490 | 0.050 | 0.120 | 5.250 | 1.680 | 3.580 | 0.500 |
| Cultivars (B) | 442.9 ** | 129.7 ** | 17.83 * | 0.740 * | 1.740 | 46.69 * | 36.50 ** | 33.59 ** | 1.320 |
| A × B | 14.66 ** | 25.19 ** | 38.35 ** | 2.020 ** | 1.720 ** | 22.09 * | 11.45 ** | 22.80 ** | 0.630 |
| Error B | 0.580 | 2.890 | 2.930 | 0.120 | 0.050 | 8.080 | 1.330 | 2.050 | 0.680 |
| Combined | |||||||||
| Years (Y) | 413.1 ** | 5930 ** | 216.3 ** | 4148 ** | 6459 ** | 4729 ** | 145.4 ** | 767.7 ** | 10.98 * |
| Reps/years | 0.71 | 6.44 | 9.59 | 1.37 | 0.94 | 11.96 | 1.400 | 0.460 | 0.820 |
| Zn levels (A) | 25.22 ** | 344.5 ** | 625.7 ** | 17.86 ** | 109.63 ** | 159.6 ** | 2.610 | 13.85 * | 0.280 |
| A × Y | 42.13 ** | 646.5 ** | 265.2 ** | 15.67 ** | 156.02 ** | 302.9 ** | 28.50 ** | 1.640 | 5.500 ** |
| Error A | 1.910 | 12.32 | 4.790 | 1.370 | 1.210 | 7.100 | 1.550 | 2.520 | 0.390 |
| Cultivars (B) | 3.970 ** | 1070 ** | 153.69 ** | 40.53 ** | 4.440 | 62.66 ** | 17.55 ** | 7.160 ** | 9.170 ** |
| B × Y | 68.27 ** | 286.3 ** | 208.1 ** | 48.89 ** | 5.290 | 49.81 * | 24.45 ** | 48.18 ** | 4.110 ** |
| A × B | 14.77 ** | 821.41 ** | 147.74 ** | 47.79 ** | 26.59 ** | 309.2 ** | 7.650 ** | 21.79 ** | 2.790 ** |
| A × B × Y | 11.00 ** | 860.2 ** | 233.0 ** | 42.06 ** | 12.23 * | 198.8 ** | 6.730 ** | 20.63 ** | 2.340 |
| Error B | 0.700 | 26.57 | 5.390 | 1.840 | 2.930 | 10.83 | 1.390 | 1.340 | 0.750 |
| Soluble Sugar under Drought, mg·g−1 Fresh Weight | |||||||||
| Zn Levels | 2 Weeks before Anthesis | at Anthesis | 2 Weeks after at Anthesis | ||||||
| Y1 | Y2 | Comb | Y1 | Y2 | Comb | Y1 | Y2 | Comb | |
| 500 ppm | 26.50a | 12.29a | 19.40a | 29.01a | 8.210c | 18.61a | 26.59a | 15.72b | 21.16a |
| 250 ppm | 14.08b | 11.36a | 12.72c | 21.61b | 6.240d | 13.93b | 27.20a | 18.71a | 22.96a |
| Water | 18.19bc | 9.430b | 13.81c | 12.44c | 17.07a | 14.70b | 24.05a | 13.84b | 18.94a |
| None | 19.88c | 11.06a | 15.47b | 20.51b | 11.24b | 15.87ab | 24.14a | 13.68b | 18.91a |
| Soluble sugar under irrigation, mg·g−1 fresh weight | |||||||||
| Zn levels | 2 Weeks before anthesis | at anthesis | 2 weeks after at anthesis | ||||||
| Y1 | Y2 | Comb | Y1 | Y2 | Comb | Y1 | Y2 | Comb | |
| 500 ppm | 10.72b | 6.870a | 8.780a | 34.11a | 6.22c | 20.16b | 27.14a | 19.32a | 23.23a |
| 250 ppm | 13.42a | 4.740b | 9.090a | 37.51a | 9.280b | 23.39a | 8.200b | 13.28d | 10.74c |
| Water | 9.470c | 4.220b | 6.850b | 22.40b | 9.790b | 16.09c | 9.630b | 17.92b | 13.78b |
| None | 7.600d | 6.270b | 6.940b | 15.43c | 11.56a | 13.50c | 6.670b | 14.98c | 10.82c |
| Soluble protein under drought, mg·g−1 fresh weight | |||||||||
| Zn levels | 2 Weeks before anthesis | at anthesis | 2 weeks after at anthesis | ||||||
| Y1 | Y2 | Comb | Y1 | Y2 | Comb | Y1 | Y2 | Comb | |
| 500 ppm | 18.28ab | 15.42ab | 16.85b | 23.12a | 11.60b | 17.36b | 48.58a | 21.80a | 35.19a |
| 250 ppm | 17.66b | 10.09c | 13.88c | 18.96b | 14.76a | 16.86b | 46.45a | 31.80a | 39.13a |
| Water | 19.80a | 19.19a | 19.49a | 26.30a | 15.64a | 20.97a | 42.60b | 35.02a | 38.81a |
| None | 21.10a | 17.84a | 19.48a | 22.72a | 16.42a | 19.57a | 44.91ab | 32.13a | 38.52a |
| Soluble protein under irrigation, mg·g−1 fresh weight | |||||||||
| Zn levels | 2 Weeks before anthesis | At anthesis | 2 weeks after at anthesis | ||||||
| Y1 | Y2 | Comb | Y1 | Y2 | Comb | Y1 | Y2 | Comb | |
| 500 ppm | 16.45a | 1.360a | 8.900a | 19.18c | 4.410a | 11.80c | 34.36b | 26.26a | 30.31b |
| 250 ppm | 17.14a | 1.640a | 9.390a | 18.16c | 2.450b | 10.30c | 48.55a | 23.39a | 35.97a |
| Water | 13.87b | 1.080a | 7.470b | 22.50b | 4.790a | 13.650b | 44.82a | 23.75a | 34.28a |
| None | 18.40a | 1.070a | 9.740a | 29.82a | 2.230b | 16.020a | 35.18b | 24.67a | 29.92b |
| Free amino acids under drought, mg·g−1 fresh weight | |||||||||
| Zn levels | 2 Weeks before anthesis | at anthesis | 2 weeks after at anthesis | ||||||
| Y1 | Y2 | Comb | Y1 | Y2 | Comb | Y1 | Y2 | Comb | |
| 500 ppm | 6.630a | 1.520b | 4.070a | 6.410b | 3.280b | 4.840c | 3.090ab | 1.670c | 2.380b |
| 250 ppm | 5.460a | 1.480b | 3.470b | 4.820c | 3.420b | 4.120c | 4.860a | 1.470c | 3.170a |
| Water | 6.200a | 1.480b | 3.840a | 9.950a | 4.430a | 7.190a | 4.520a | 2.210b | 3.360a |
| None | 6.150a | 1.970a | 4.060a | 7.160b | 3.940ab | 5.550b | 2.200b | 2.820a | 2.500b |
| Free amino acids under irrigation, mg·g−1 fresh weight | |||||||||
| Zn levels | 2 Weeks before anthesis | at anthesis | 2 weeks after at anthesis | ||||||
| Y1 | Y2 | Comb | Y1 | Y2 | Comb | Y1 | Y2 | Comb | |
| 500 ppm | 10.26bc | 8.390a | 9.330a | 13.44a | 6.670a | 10.05ab | 3.380b | 2.990ab | 3.190a |
| 250 ppm | 10.49b | 9.970a | 10.23a | 11.52a | 5.800a | 8.660b | 3.990ab | 2.440b | 3.210a |
| Water | 12.93a | 6.540b | 9.73a | 13.92a | 7.420a | 10.67a | 4.350a | 2.570b | 3.460a |
| None | 11.24ab | 8.660a | 9.95a | 12.85a | 5.710a | 9.280ab | 2.980b | 3.580a | 3.280a |
| Zn Levels | Plant Stages | 2 Weeks before Anthesis | at Anthesis | 2 Weeks after Anthesis | ||||||
|---|---|---|---|---|---|---|---|---|---|---|
| Compound | SS | SP | AA | SS | SP | AA | SS | SP | AA | |
| 500 ppm | SS | 0.94 ** | 0.97 ** | 0.98 ** | 0.87 * | 0.96 ** | 0.82 * | |||
| SP | −0.99 ** | 0.86 * | 0.98 ** | 0.93 ** | 0.99 ** | 0.79 | ||||
| AA | 1.00 ** | −1.00 ** | 0.98 ** | 0.97 ** | 0.41 | 0.31 | ||||
| 250 ppm | SS | 0.74 | 0.56 | 0.92 ** | 0.93 ** | 0.71 | 0.65 | |||
| SP | −0.50 | 0.97 ** | 0.99 ** | 0.89 * | −0.99 ** | 1.00 ** | ||||
| AA | 0.87 * | −0.53 | 0.98 ** | 0.97 ** | 0.93 ** | −0.89 * | ||||
| water | SS | 0.16 | 1.00 ** | −0.73 | −0.82 * | 0.65 | 0.70 | |||
| SP | −0.91 ** | 0.15 | 0.98 ** | 0.99 ** | −0.98 ** | 0.51 | ||||
| AA | 0.97 ** | −0.96 ** | 0.95 ** | 0.99 ** | 0.96 ** | −0.90 * | ||||
| none | SS | 0.99 ** | 0.98 ** | 0.98 ** | 0.90 * | 0.94 ** | −0.82 * | |||
| SP | −0.87 * | 0.96 ** | 0.61 | 0.83 * | −0.97 ** | −0.84 * | ||||
| AA | 0.96 ** | −0.90 * | 0.41 | 0.97 ** | −0.72 | 0.74 | ||||
| Cultivars | Plant Stages | 2 Weeks before Anthesis | at Anthesis | 2 Weeks after Anthesis | ||||||
|---|---|---|---|---|---|---|---|---|---|---|
| Compound | SS | SP | AA | SS | SP | AA | SS | SP | AA | |
| Sakha 69 | SS | 0.17 | 0.71 | 0.68 | 0.78 | 0.97 ** | 0.96 ** | |||
| SP | −0.96 ** | 0.24 | 0.97 ** | 0.93 ** | −0.87 * | 0.88 * | ||||
| AA | 0.99 ** | −0.97 ** | 0.96 ** | 1.00 ** | 0.50 | −0.74 | ||||
| G168 | SS | 0.94 ** | −0.02 | 0.89 * | 0.93 ** | 0.90 * | 0.79 | |||
| SP | −0.98 ** | −0.04 | 0.80 * | 0.99 ** | 0.66 | 0.91 * | ||||
| AA | 0.98 ** | −0.99 ** | 0.83 * | 0.99 ** | 0.49 | 0.34 | ||||
| Gem 11 | SS | 0.79 | 0.97 ** | 0.96 ** | 0.99 ** | 0.98 ** | 0.59 | |||
| SP | −0.68 | 0.70 | 1.00 ** | 0.94 ** | −1.00 ** | 0.61 | ||||
| AA | 0.98 ** | −0.56 | 0.98 ** | 0.98 ** | 0.86* | −0.87 * | ||||
| Item | Value | Item | Value |
|---|---|---|---|
| Sand (%) | 27.4 | Total nitrogen (%) | 0.72 |
| Silt (%) | 24.3 | KCl-extractable N (mg·kg−1) | 41.23 |
| Clay (%) | 48.3 | Fe mg/kg | 13.21 |
| Texture grade | Clay | Mn mg/kg | 5.152 |
| EC (1:1 extract) dS·m−1 | 0.47 | Cu mg/kg | 1.31 |
| pH | 8.2 | Zn mg/kg | 2.12 |
| CaCO3 (%) | 3.4 | Soil moisture at F. capacity | 46% |
| Organic matter (%) | 1.75 | Soil moisture at wilting point. | 28% |
| NaHCO3-extractable P (mg·kg−1) | 4.36 | NH4OAC-extractable K (mg·kg−1) | 49.24 |
| Seasons | Season 2018/19 | Season 2019/20 | |||
|---|---|---|---|---|---|
| Experiment | Drought Stress Experiment | Normal Irrig. Experiment | Drought Stress Experiment | Normal Irrig. Experiment | |
| Time | |||||
| Before 2nd irrigation | 37.84 | 39.84 | 38.56 | 37.12 | |
| Before 4th irrigation | 23.86 | 33.72 | 22.81 | 36.89 | |
| At anthesis | 17.15 | 38.17 | 18.52 | 37.55 | |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Mahdy, R.E.; Alghamdi, S.A.; Amro, A.; Tammam, S.A. Changes in Carbon and Nitrogen Metabolites before, at, and after Anthesis for Wheat Cultivars in Response to Reduced Soil Water and Zinc Foliar Application. Plants 2022, 11, 1261. https://0-doi-org.brum.beds.ac.uk/10.3390/plants11091261
Mahdy RE, Alghamdi SA, Amro A, Tammam SA. Changes in Carbon and Nitrogen Metabolites before, at, and after Anthesis for Wheat Cultivars in Response to Reduced Soil Water and Zinc Foliar Application. Plants. 2022; 11(9):1261. https://0-doi-org.brum.beds.ac.uk/10.3390/plants11091261
Chicago/Turabian StyleMahdy, Rasha E., Sameera A. Alghamdi, Ahmed Amro, and Suzan A. Tammam. 2022. "Changes in Carbon and Nitrogen Metabolites before, at, and after Anthesis for Wheat Cultivars in Response to Reduced Soil Water and Zinc Foliar Application" Plants 11, no. 9: 1261. https://0-doi-org.brum.beds.ac.uk/10.3390/plants11091261