
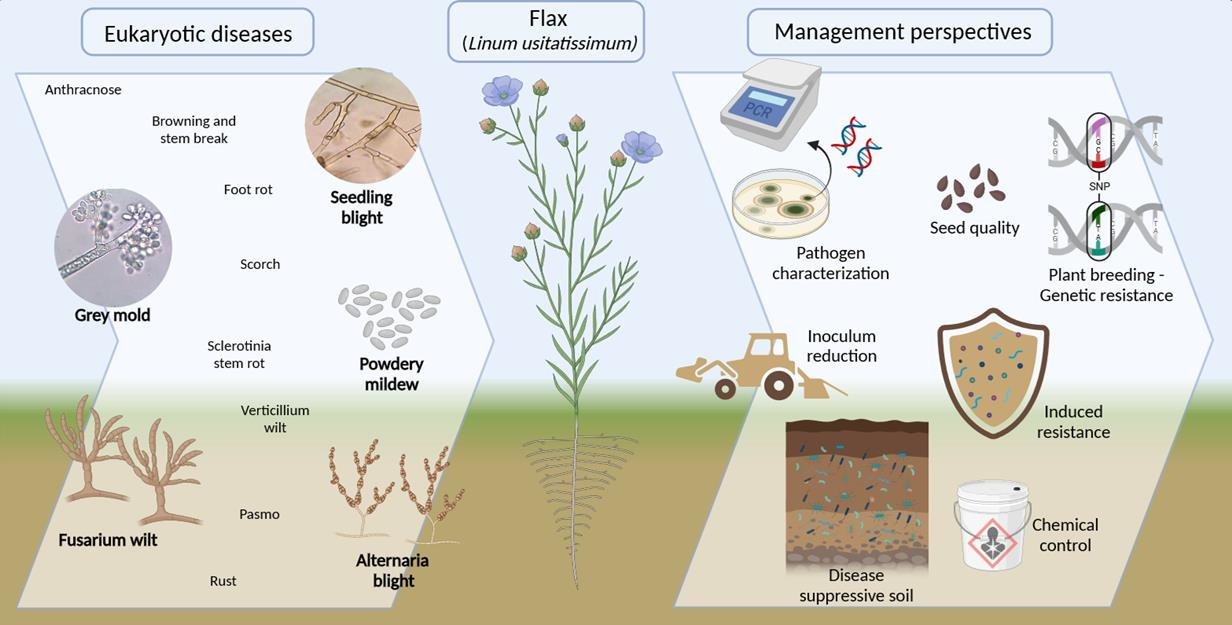
Overview and Management of the Most Common Eukaryotic Diseases of Flax (Linum usitatissimum)

 , , and
, , and
Abstract
:
1. Introduction
2. Air-Borne Diseases
2.1. Powdery Mildew—Golovinomyces orontii/Podosphaera lini/Erysiphe lini/Leveillula taurica
2.2. Rust—Melampsora lini
3. Seed-Borne Diseases
3.1. Alternaria Blight—Alternaria linicola
3.2. Anthracnose—Colletotrichum lini
3.3. Basal Stem Blight/Foot Rot—Boeremia linicola
3.4. Browning and Stem Break—Aureobasidium pullulans var. lini
3.5. Gray Mold—Botrytis cinerea
3.6. Pasmo—Septoria linicola (syn.: Mycosphaerella linicola)
4. Soil-Borne Diseases
4.1. Fusarium wilt—Fusarium oxysporum f. sp. lini
4.2. Sclerotinia Stem Rot—Sclerotinia sclerotiorum
4.3. Scorch—Globisporangium megalacanthum and Berkeleyomyces basicola
4.4. Seedling Blight/Root Rot—Rhizoctonia solani
4.5. Verticillium wilt—Verticillium dahliae
5. Perspectives in Flax Disease Management
5.1. Pathogen Characterization
5.2. Sanitary Quality of Seeds
5.3. Inoculum Reduction
5.4. Disease Suppressive Soils
5.5. Chemical Control
- An increasing amount of fungicide-resistant pathogen strains;
- A rising demand by consumers and plant product retailers for very low or even no synthetic chemical residues at all;
- More and more restrictive international regulations on the permitted levels of chemical residues in the environment and on the registration and eco-toxicological impact of pesticides (e.g., EU Directive 2009/128 on the sustainable use of pesticides and EU Green Deal 2019 Farm to Fork Strategy).
- Even if a global tendency tends toward a “zero” pesticide agriculture, another way is considered by researchers: integrated pest management. As described by Ons et al. (2020) [115] in a review, the increasing global food demand requires efficient ways to fight crop diseases. Combining synthetic pesticides and biocontrol products could be a suitable option.
5.6. Plant Breeding and Genetic Resistance
5.7. Induced Resistance
6. Conclusions
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
Appendix A

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Disease | Gene Families | Chromosome Location |
|---|---|---|
| Alternaria Blight | Heat shock protein | 14 |
| Fusarium wilt | AAA-ATPase | 8 |
| CYP709B2 Cytochrome P450 | 1 | |
| DA1-related protein 4 nucleotide-binding LRR protein | 1 | |
| Exportin 1A | 1 | |
| KIP1-like protein | 1 | |
| NADP-malic enzyme 4 | 1 | |
| Pectate lyase family protein | 8 | |
| Protein kinase | 1 | |
| Receptor-like protein | 13 | |
| Rubredoxin-like superfamily protein | 11 | |
| SPFH/B and 7/PHB domain-containing membrane-associated protein | 8 | |
| Sucrose nonfermenting 1 (SNF1)-related protein kinase | 13 | |
| Voltage-dependant anion channel 1 | 8 | |
| Powdery mildew | AP2—Integrase-type DNA-binding superfamily protein | 13 |
| C-8,7 sterol isomerase | 1 | |
| Cellulose synthase-like D5 | 15 | |
| Coiled-coil-nucleotide-binding site domain leucin rich repeat | 10, 13, 14 | |
| Dirigent protein | 5, 15 | |
| DNAJ homologue | 13 | |
| Early responsive to dehydration stress family protein | 7 | |
| Homeodomain-like superfamily protein | 11 | |
| Hydroxymethylglutaryl-CoA synthase | 5 | |
| Leucine Rich Repeat Protein | 4 | |
| Lysin motif receptor-like kinases | 13 | |
| Mildew locus O | 7, 9, 13 | |
| Nitrate transporter | 4 | |
| Pathogenesis-related protein | 14 | |
| Pectin lyase-like superfamily protein | 5 | |
| Receptor-like kinase protein | 1, 2, 3, 4, 6, 7, 8, 9, 10, 12, 13, 14, 15 | |
| Receptor-like protein | 5, 6, 12, 13 | |
| Resistance to powdery mildew 8 | 13 | |
| Sucrose synthase | 3 | |
| Toll/interleukin 1 receptor-like domain-nucleotide binding site-Leucine rich repeat | 2, 4, 5, 7, 8, 9, 12, 13, 14 | |
| Transmembrane coiled-coil protein | 5, 7, 9, 11, 12, 14 | |
| WRKY | 4, 5, 6, 11, 15 | |
| ZPR1 zinc-finger domain protein | 7 | |
| Pasmo | Coiled-coil-nucleotide-binding site domain-leucin rich repeat | 4, 5, 8, 12 |
| Nucleotide-binding site domain | 4 | |
| Nucleotide-binding site–leucine-rich repeats | 8 | |
| Receptor-like kinase protein | 1, 2, 3, 4, 6, 7, 8, 9, 10, 12, 13, 15 | |
| Receptor-like protein | 3, 6, 8, 10 | |
| Toll/interleukin 1 receptor-like domain-nucleotide binding site | 12 | |
| Toll/interleukin 1 receptor-like domain-nucleotide binding site-leucine rich repeat | 3, 4, 5, 8 | |
| Toll/interleukin 1 receptor-nucleotide binding site | 5 | |
| Toll/interleukin 1 receptor-unknown/random | 4, 5, 8, 13, 15 | |
| Transmembrane coiled-coil protein | 4, 5, 8, 9, 11, 12, 15 |
References
- Rashid, K.Y. Principal Diseases of Flax, 1st ed.; CRC Press: Boca Raton, FL, USA, 2003; pp. 104–135. ISBN 978-0-429-20585-9. [Google Scholar]
- van Zeist, W.; Bakker-Heeres, J.A.H. Evidence for Linseed Cultivation before 6000 Bc. J. Archaeol. Sci. 1975, 2, 215–219. [Google Scholar] [CrossRef]
- FAOSTAT. Available online: https://www.fao.org/faostat/en/#data/QCL (accessed on 2 June 2023).
- Allen, E.; Alvarez, S.; ISTA Secretariat. Chapter 7: Seed Health Testing. In Rules Proposals for the International Rules for Seed Testing 2023 Edition; IRST: Wallisellen, Switzerland, 2023; pp. 1–14. [Google Scholar]
- Kvavadze, E.; Bar-Yosef, O.; Belfer-Cohen, A.; Boaretto, E.; Jakeli, N.; Matskevich, Z.; Meshveliani, T. 30,000-Year-Old Wild Flax Fibers. Science 2009, 325, 1359. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Muir, A.D.; Westcott, N.D. Flax: The Genus Linum; CRC Press: Boca Raton, FL, USA, 2003; ISBN 978-0-203-43750-6. [Google Scholar]
- In Focus: An Economic Overview of Flax in 2022. Available online: https://allianceflaxlinenhemp.eu/en/flax-linen-economic-observatory (accessed on 18 July 2023).
- Lin Fibre—Une Valorisation de Toutes les Composantes de la Plante. Available online: https://www.arvalis.fr/infos-techniques/une-valorisation-de-toutes-les-composantes-de-la-plante (accessed on 10 January 2023).
- Yadav, B.; Kaur, V.; Narayan, O.P.; Yadav, S.K.; Kumar, A.; Wankhede, D.P. Integrated Omics Approaches for Flax Improvement under Abiotic and Biotic Stress: Current Status and Future Prospects. Front. Plant Sci. 2022, 13, 931275. [Google Scholar] [CrossRef] [PubMed]
- Münch, S.; Lingner, U.; Floss, D.S.; Ludwig, N.; Sauer, N.; Deising, H.B. The Hemibiotrophic Lifestyle of Colletotrichum Species. J. Plant Physiol. 2008, 165, 41–51. [Google Scholar] [CrossRef]
- Dean, R.; Van Kan, J.A.L.; Pretorius, Z.A.; Hammond-Kosack, K.E.; Di Pietro, A.; Spanu, P.D.; Rudd, J.J.; Dickman, M.; Kahmann, R.; Ellis, J.; et al. The Top 10 Fungal Pathogens in Molecular Plant Pathology. Mol. Plant Pathol. 2012, 13, 414–430. [Google Scholar] [CrossRef] [Green Version]
- Cheng, Y.; Tang, X.; Gao, C.; Li, Z.; Chen, J.; Guo, L.; Wang, T.; Xu, J. Molecular Diagnostics and Pathogenesis of Fungal Pathogens on Bast Fiber Crops. Pathogens 2020, 9, 223. [Google Scholar] [CrossRef] [Green Version]
- Novakovskiy, R.O.; Dvorianinova, E.M.; Rozhmina, T.A.; Kudryavtseva, L.P.; Gryzunov, A.A.; Pushkova, E.N.; Povkhova, L.V.; Snezhkina, A.V.; Krasnov, G.S.; Kudryavtseva, A.V.; et al. Data on Genetic Polymorphism of Flax (Linum Usitatissimum L.) Pathogenic Fungi of Fusarium, Colletotrichum, Aureobasidium, Septoria, and Melampsora Genera. Data Brief 2020, 31, 105710. [Google Scholar] [CrossRef]
- Singh, R.B.; Singh, H.K.; Parmar, A. Yield Loss Assessment Due to Alternaria Blight and Its Management in Linseed. Pak. J. Biol. Sci. 2013, 17, 511–516. [Google Scholar] [CrossRef] [PubMed]
- Gruzdevienė, E.; Brazauskienė, I.; Repečkienė, J.; Lugauskas, A. The Occurrence of Pathogenic Fungi During Flax Growing Season in Central Lithuania. J. Plant Prot. Res. 2008, 48, 255–265. [Google Scholar] [CrossRef]
- Decognet, V.; Cerceau, V.; Jouan, B. Control of Phoma Exigua Var. Linicola on Flax by Seed and Foliar Spray Treatments with Fungicides. Crop Prot. 1994, 13, 105–108. [Google Scholar] [CrossRef]
- Kanapin, A.; Bankin, M.; Rozhmina, T.; Samsonova, A.; Samsonova, M. Genomic Regions Associated with Fusarium Wilt Resistance in Flax. Int. J. Mol. Sci. 2021, 22, 12383. [Google Scholar] [CrossRef] [PubMed]
- Planchon, A.; Durambur, G.; Besnier, J.-B.; Plasson, C.; Gügi, B.; Bernard, S.; Mérieau, A.; Trouvé, J.-P.; Dubois, C.; Laval, K.; et al. Effect of a Bacillus Subtilis Strain on Flax Protection against Fusarium Oxysporum and Its Impact on the Root and Stem Cell Walls. Plant Cell Environ. 2021, 44, 304–322. [Google Scholar] [CrossRef] [PubMed]
- Ferryman, S.A.M.; Fitt, B.D.L.; Harold, J.F.S. Factors Affecting the Development of Botrytis Cinerea (Grey Mould) on Linseed (Linum Usitatissimum) Buds, Flowers and Capsules. Ann. Appl. Biol. 2002, 140, 1–12. [Google Scholar] [CrossRef]
- Williamson, B.; Tudzynski, B.; Tudzynski, P.; Van Kan, J.A.L. Botrytis Cinerea: The Cause of Grey Mould Disease. Mol. Plant Pathol. 2007, 8, 561–580. [Google Scholar] [CrossRef] [PubMed]
- Paumier, D.; Bammé, B.; Penaud, A.; Valade, R.; Suffert, F. First Report of the Sexual Stage of the Flax Pathogen Mycosphaerella Linicola in France and Its Impact on Pasmo Epidemiology. Plant Pathol. 2021, 70, 475–483. [Google Scholar] [CrossRef]
- Perryman, S.; Gladders, P.; Fitt, B.D.L. Autumn Sowing Increases Severity of Pasmo (Mycosphaerella Linicola) on Linseed in the UK. Ann. Appl. Biol. 2009, 154, 19–32. [Google Scholar] [CrossRef]
- Lapalu, N.; Simon, A.; Demenou, B.; Paumier, D.; Guillot, M.-P.; Gout, L.; Suffert, F.; Valade, R. Complete Genome Sequences of Septoria Linicola: A Resource for Studying a Damaging Flax Pathogen. Mol. Plant Microbe Interact. 2023, 36, 59–63. [Google Scholar] [CrossRef]
- Halley, S.; Bradley, C.A.; Lukach, J.R.; McMullen, M.; Knodel, J.J.; Endres, G.J.; Gregoire, T. Distribution and Severity of Pasmo on Flax in North Dakota and Evaluation of Fungicides and Cultivars for Management. Plant Dis. 2004, 88, 1123–1126. [Google Scholar] [CrossRef] [Green Version]
- Aly, A.A.; Mohamed, H.I.; Mansour, M.T.M.; Omar, M.R. Suppression of Powdery Mildew on Flax by Foliar Application of Essential Oils. J. Phytopathol. 2013, 161, 376–381. [Google Scholar] [CrossRef]
- Saharan, G.S.; Mehta, N. Sclerotinia Diseases of Crop Plants: Biology, Ecology and Disease Management; Springer: Dordrecht, The Netherlands, 2008; ISBN 978-1-4020-8407-2. [Google Scholar]
- Wiersema, H.T. Flax Scorch. Euphytica 1955, 4, 197–205. [Google Scholar] [CrossRef]
- Kangatharalingam, N. Variability in Rhizoctonia Solani Kühn Pathogenic on Flax; Electronic Theses and Dissertations; South Dakota State University: Brookings, SD, USA, 1987. [Google Scholar]
- Aly, A.; Hussein, E.; Asran, A.; El-Hawary, O.; Mosa, A.; Mostafa, M. Non-Differential Interaction between Isolates of Rhizoctonia Solani and Flax Cultivars. Icel. Agric. Sci. 2013, 26, 37–44. [Google Scholar]
- Blum, A.; Bressan, M.; Zahid, A.; Trinsoutrot-Gattin, I.; Driouich, A.; Laval, K. Verticillium Wilt on Fiber Flax: Symptoms and Pathogen Development in planta. Plant Dis. 2018, 102, 2421–2429. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Braun, U.; Cook, R.T.A. Taxonomic Manual of the Erysiphales (Powdery Mildews); CBS-KNAW Fungal Biodiversity Centre: Utrecht, The Netherlands, 2012; ISBN 978-90-70351-89-2. [Google Scholar]
- Ajithkumar, K.; Savitha, A.S.; Mahadevakumar, S.; Maharachchikumbura, S.S.N.; Sreenivasa, M.Y.; Rathnakumar, A.L.; Sujatha, M. First Report of Molecular Detection of Leveillula Taurica Associated with Powdery Mildew of Linseed (Linum Usitatissimum) from India. Plant Dis. 2022, 106, 1529. [Google Scholar] [CrossRef]
- Braun, U. Podosphaera Lini (Ascomycota, Erysiphales) Revisited and Reunited with Oidium Lini. PPQ 2019, 9, 128–138. [Google Scholar] [CrossRef]
- Preston, C.D.; Cook, R.T.A. Podosphaera Lini, an Overlooked Powdery Mildew in Britain. Field Mycol. 2019, 20, 55–57. [Google Scholar] [CrossRef]
- Melloy, P.; Del Ponte, E.M.; Sparks, A.H. Timing of Triazole-Based Spray Schedules for Managing Mungbean Powdery Mildew in Australia: A Meta-Analysis. Plant Dis. 2022, 106, 918–924. [Google Scholar] [CrossRef] [PubMed]
- Rashid, K.; Duguid, S. Inheritance of Resistance to Powdery Mildew in Flax. Can. J. Plant Pathol. 2005, 27, 404–409. [Google Scholar] [CrossRef]
- You, F.M.; Rashid, K.Y.; Zheng, C.; Khan, N.; Li, P.; Xiao, J.; He, L.; Yao, Z.; Cloutier, S. Insights into the Genetic Architecture and Genomic Prediction of Powdery Mildew Resistance in Flax (Linum Usitatissimum L.). Int. J. Mol. Sci. 2022, 23, 4960. [Google Scholar] [CrossRef]
- Speck, A.; Trouvé, J.-P.; Enjalbert, J.; Geffroy, V.; Joets, J.; Moreau, L. Genetic Architecture of Powdery Mildew Resistance Revealed by a Genome-Wide Association Study of a Worldwide Collection of Flax (Linum Usitatissimum L.). Front. Plant Sci. 2022, 13, 871633. [Google Scholar] [CrossRef]
- EU Pesticides Database. Available online: https://food.ec.europa.eu/plants/pesticides/eu-pesticides-database_en (accessed on 14 June 2023).
- CTGB Admissions. Available online: https://pesticidesdatabase.ctgb.nl/en/authorisations (accessed on 14 June 2023).
- State Institution “Main State Inspectorate for Seed Production, Quarantine and Plant Protection”. Available online: https://www.ggiskzr.by/ (accessed on 14 June 2023).
- Ephy. Available online: https://ephy.anses.fr/ (accessed on 14 June 2023).
- Pesticides Register of Authorised Plant Protection Products. Available online: https://secure.pesticides.gov.uk/pestreg/ (accessed on 14 June 2023).
- Consult Authorisations of Plant Protection Products. Available online: https://fytoweb.be/en/plant-protection-products/consult-authorisations-plant-protection-products (accessed on 14 June 2023).
- Government of Canada, H.C. Pesticide Product Information—Health Canada. Available online: https://pest-control.canada.ca/pesticide-registry/en/index.html (accessed on 14 June 2023).
- Lawrence, G.J.; Dodds, P.N.; Ellis, J.G. Rust of Flax and Linseed Caused by Melampsora Lini. Mol. Plant Pathol. 2007, 8, 349–364. [Google Scholar] [CrossRef]
- Ellis, J.G.; Dodds, P.N.; Lawrence, G.J. Flax Rust Resistance Gene Specificity Is Based on Direct Resistance-Avirulence Protein Interactions. Annu. Rev. Phytopathol. 2007, 45, 289–306. [Google Scholar] [CrossRef] [PubMed]
- The Flax Genome; Compendium of Plant Genomes; You, F.M.; Fofana, B. (Eds.) Springer: Cham, Switzerland, 2023; ISBN 978-3-031-16060-8. [Google Scholar]
- Sran, R.S.; Paul, S.; Kumar, A.; Sekhon, B.S. Genetics of Resistance to Rust and Powdery Mildew in Linseed (Linum Usitatissimum L.). Indian Phytopathol. 2021, 74, 633–637. [Google Scholar] [CrossRef]
- Vloutoglou, I.; Fitt, B.D.L.; Lucas, J.A. Survival and Seed to Seedling Transmission of Alternaria Linicola on Linseed. Ann. Appl. Biol. 1995, 127, 33–47. [Google Scholar] [CrossRef]
- Singh, N.; Kumar, R.; Kumar, S.; Singh, P.K.; Yadav, H.K. Mapping QTLs for Alternaria Blight in Linseed (Linum Usitatissimum L.). 3 Biotech 2021, 11, 91. [Google Scholar] [CrossRef] [PubMed]
- Chapter 7: Seed Health Testing. Int. Rules Seed Test. 2022, 2022, i-7-6. [CrossRef]
- Council Directive 2002/57/EC of 13 June 2002 on the Marketing of Seed of Oil and Fibre Plants; European Commission: Brussels, Belgium, 2022.
- Pest Lists. Available online: https://pestlist.worldseed.org/public/pestlist.jsp (accessed on 18 July 2023).
- Hawksworth, D.L. CMI Descriptions of Fungi and Bacteria. Mycopathologia 1990, 111, 109. [Google Scholar] [CrossRef]
- Yurlova, N.; Hoog, S.; Gerrits van den Ende, B. Taxonomy of Aureobasidium and Allied Genera. Stud. Mycol. 1999, 1999, 63–69. [Google Scholar]
- Bozoudi, D.; Tsaltas, D. The Multiple and Versatile Roles of Aureobasidium Pullulans in the Vitivinicultural Sector. Fermentation 2018, 4, 85. [Google Scholar] [CrossRef] [Green Version]
- You, F.M.; Rashid, K.Y.; Cloutier, S. Genomic Designing for Genetic Improvement of Biotic Stress Resistance in Flax. In Genomic Designing for Biotic Stress Resistant Oilseed Crops; Kole, C., Ed.; Springer: Cham, Switzerland, 2022; pp. 311–345. ISBN 978-3-030-91035-8. [Google Scholar]
- Mbengue, M.; Navaud, O.; Peyraud, R.; Barascud, M.; Badet, T.; Vincent, R.; Barbacci, A.; Raffaele, S. Emerging Trends in Molecular Interactions between Plants and the Broad Host Range Fungal Pathogens Botrytis Cinerea and Sclerotinia Sclerotiorum. Front. Plant Sci. 2016, 7, 422. [Google Scholar] [CrossRef] [Green Version]
- He, L.; Xiao, J.; Rashid, K.Y.; Yao, Z.; Li, P.; Jia, G.; Wang, X.; Cloutier, S.; You, F.M. Genome-Wide Association Studies for Pasmo Resistance in Flax (Linum Usitatissimum L.). Front. Plant Sci. 2019, 9, 1982. [Google Scholar] [CrossRef] [PubMed]
- He, L.; Xiao, J.; Rashid, K.Y.; Jia, G.; Li, P.; Yao, Z.; Wang, X.; Cloutier, S.; You, F.M. Evaluation of Genomic Prediction for Pasmo Resistance in Flax. Int. J. Mol. Sci. 2019, 20, 359. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Samsonova, A.; Kanapin, A.; Bankin, M.; Logachev, A.; Gretsova, M.; Rozhmina, T.; Samsonova, M. A Genomic Blueprint of Flax Fungal Parasite Fusarium Oxysporum f. Sp. Lini. Int. J. Mol. Sci. 2021, 22, 2665. [Google Scholar] [CrossRef]
- Baayen, R.P.; O’Donnell, K.; Bonants, P.J.M.; Cigelnik, E.; Kroon, L.P.N.M.; Roebroeck, E.J.A.; Waalwijk, C. Gene Genealogies and AFLP Analyses in the Fusarium Oxysporum Complex Identify Monophyletic and Nonmonophyletic Formae Speciales Causing Wilt and Rot Disease. Phytopathology 2000, 90, 891–900. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Spielmeyer, W.; Green, A.G.; Bittisnich, D.; Mendham, N.; Lagudah, E.S. Identification of Quantitative Trait Loci Contributing to Fusarium Wilt Resistance on an AFLP Linkage Map of Flax (Linum Usitatissimum). Theor. Appl. Genet. 1998, 97, 633–641. [Google Scholar] [CrossRef]
- Mazurier, S.; Corberand, T.; Lemanceau, P.; Raaijmakers, J.M. Phenazine Antibiotics Produced by Fluorescent Pseudomonads Contribute to Natural Soil Suppressiveness to Fusarium Wilt. ISME J. 2009, 3, 977–991. [Google Scholar] [CrossRef]
- Mitchell, S.J.; Jellis, G.J.; Cox, T.W. Sclerotinia Sclerotiorum on Linseed. Plant Pathol. 1986, 35, 403–405. [Google Scholar] [CrossRef]
- Mederick, F.; Piening, L.J. Sclerotinia Sclerotiorum on Oil and Fibre Flax in Alberta. Can. Plant Dis. Surv. 1982, 62, 11. [Google Scholar]
- Roy, J.; del Río Mendoza, L.E.; Bandillo, N.; McClean, P.E.; Rahman, M. Genetic Mapping and Genomic Prediction of Sclerotinia Stem Rot Resistance to Rapeseed/Canola (Brassica Napus L.) at Seedling Stage. Theor. Appl. Genet. 2022, 135, 2167–2184. [Google Scholar] [CrossRef] [PubMed]
- Talukder, Z.I.; Underwood, W.; Misar, C.G.; Seiler, G.J.; Cai, X.; Li, X.; Qi, L. A Quantitative Genetic Study of Sclerotinia Head Rot Resistance Introgressed from the Wild Perennial Helianthus Maximiliani into Cultivated Sunflower (Helianthus Annuus L.). Int. J. Mol. Sci. 2022, 23, 7727. [Google Scholar] [CrossRef]
- Zeng, W.; Kirk, W.; Hao, J. Field Management of Sclerotinia Stem Rot of Soybean Using Biological Control Agents. Biol. Control 2012, 60, 141–147. [Google Scholar] [CrossRef]
- Whipps, J.M.; Sreenivasaprasad, S.; Muthumeenakshi, S.; Rogers, C.W.; Challen, M.P. Use of Coniothyrium Minitans as a Biocontrol Agent and Some Molecular Aspects of Sclerotial Mycoparasitism. Eur. J. Plant Pathol. 2008, 121, 323–330. [Google Scholar] [CrossRef]
- Zeng, W.; Wang, D.; Kirk, W.; Hao, J. Use of Coniothyrium Minitans and Other Microorganisms for Reducing Sclerotinia Sclerotiorum. Biol. Control 2012, 60, 225–232. [Google Scholar] [CrossRef]
- Albert, D.; Dumonceaux, T.; Carisse, O.; Beaulieu, C.; Filion, M. Combining Desirable Traits for a Good Biocontrol Strategy against Sclerotinia Sclerotiorum. Microorganisms 2022, 10, 1189. [Google Scholar] [CrossRef] [PubMed]
- Cariou, E.; Brochard, M.; Raynal, G. Flax scorch due to Chalara elegans (syn. Thielaviopsis basicola) and several Pythium species. Phytoma. Défense Végétaux 2003, 566, 6–9. [Google Scholar]
- Nel, W.J.; Duong, T.A.; de Beer, Z.W.; Wingfield, M.J. Black Root Rot: A Long Known but Little Understood Disease. Plant Pathol. 2019, 68, 834–842. [Google Scholar] [CrossRef]
- Cariou-Pham, E.; Bonnan, S. Biological Control of Flax Scorch Using Glomus Intraradices and Trichoderma Atroviride. In Modern Multidisciplinary Applied Microbiology; John Wiley & Sons, Ltd.: Hoboken, NJ, USA, 2006; pp. 380–384. ISBN 978-3-527-61190-4. [Google Scholar]
- Brentzel, W.E. Disease of Flax Caused by a Species of Rhizoctonia. Phytopathology 1923, 13, 53. [Google Scholar]
- Pederson, V.D. Etiology and Control of a Seedling Blight of Flax Caused by Rhizoctonia Solani Kühn. Master’s Thesis, South Dakota State College of Agriculture and Mechanic Arts, Brookings, SD, USA, 1957. [Google Scholar]
- Horne, C.W. Texas Plant Disease Handbook. Chapter: Diseases on Flax (Linum Usitatissimum L.); Texas A&M: College Station, TX, USA, 1983. [Google Scholar]
- Sharon, M.; Kuninaga, S.; Hyakumachi, M.; Sneh, B. The Advancing Identification and Classification of Rhizoctonia Spp. Using Molecular and Biotechnological Methods Compared with the Classical Anastomosis Grouping. Mycoscience 2006, 47, 299–316. [Google Scholar] [CrossRef]
- Hwang, S.F.; Gossen, B.D.; Chang, K.F.; Turnbull, G.D.; Howard, R.J.; Blade, S.F. Etiology, Impact and Control of Rhizoctonia Seedling Blight and Root Rot of Chickpea on the Canadian Prairies. Can. J. Plant Sci. 2003, 83, 959–967. [Google Scholar] [CrossRef]
- Yang, J.; Karbanda, P.D.; Wang, H.; McAndrew, D.W. Characterization, Virulence, and Genetic Variation of Rhizoctonia Solani AG-9 in Alberta. Plant Dis. 1996, 80, 513–518. [Google Scholar] [CrossRef]
- Nelson, B.; Helms, T.; Christianson, T.; Kural, I. Characterization and Pathogenicity of Rhizoctonia from Soybean. Plant Dis. 1996, 80, 74–80. [Google Scholar] [CrossRef]
- Misawa, T.; Kurose, D.; Mori, M.; Toda, T. Characterization of Japanese Rhizoctonia Solani AG-2-1 Isolates Using RDNA-ITS Sequences, Culture Morphology, and Growth Temperature. J. Gen. Plant Pathol. 2018, 84, 387–394. [Google Scholar] [CrossRef]
- Yu, H.; Zhou, Q.; Hwang, S.F.; Ho, A.J.; Chang, K.F.; Strelkov, S.E.; He, Y.; Conner, R.L.; Harding, M.W. Pathogenicity, Anastomosis Groups, Host Range, and Genetic Diversity of Rhizoctonia Species Isolated from Soybean, Pea, and Other Crops in Alberta and Manitoba, Canada. Can. J. Plant Sci. 2022, 102, 301–315. [Google Scholar] [CrossRef]
- Anderson, N.A. Evaluation of the Rhizoctonia Complex in Relation to Seedling Blight of Flax. Plant Dis. Rep. 1977, 61, 140–142. [Google Scholar]
- Aly, A.A.; El-Samawaty, A.E.R.M.A.; Yassin, M.A. Non-Differential Interaction between Isolates of Rhizoctonia Solani and Egyptian Cotton Cultivars. Res. Crops 2014, 15, 655–661. [Google Scholar] [CrossRef]
- Rashid, K.Y. Principal Diseases of Flax. In Flax: The genus Linum; Muir, A.D., Westcott, N.D., Eds.; CRC Press: London, UK, 2015; p. 32. ISBN 978-0-429-20585-9. [Google Scholar]
- Keene, C.; Dakota, N. Flax Production in North Dakota. NDSU Ext. 2020, 1038, 1–8. [Google Scholar]
- Vest, G.; Comstock, V.E. Resistance in Flax to Seedling Blight Caused by Rhizoctonia Solani. Phytopathology 1968, 58, 1161. [Google Scholar]
- Ashour, A.Z.A.; Aly, A.A.; Asran, A.A.; Zayed, S.M.E. Effect of Seed-Dressing Fungicides and Seed-Rate on Incidence of Flax Seedling Blight. J. Agric. Chem. Biotechnol. 2019, 10, 141–143. [Google Scholar] [CrossRef]
- Song, R.; Li, J.; Xie, C.; Jian, W.; Yang, X. An Overview of the Molecular Genetics of Plant Resistance to the Verticillium Wilt Pathogen Verticillium Dahliae. Int. J. Mol. Sci. 2020, 21, 1120. [Google Scholar] [CrossRef] [Green Version]
- Chen, D.; Zebarth, B.J.; Goyer, C.; Comeau, L.-P.; Nahar, K.; Dixon, T. Effect of Biofumigation on Population Densities of Pratylenchus Spp. and Verticillium Spp. and Potato Yield in Eastern Canada. Am. J. Potato Res. 2022, 99, 229–242. [Google Scholar] [CrossRef]
- Deketelaere, S.; Spiessens, K.; Pollet, S.; Tyvaert, L.; Rooster, L.D.; Callens, D.; França, S.C.; Höfte, M. Towards Practical Application of Verticillium Isaacii Vt305 to Control Verticillium Wilt of Cauliflower: Exploring Complementary Biocontrol Strategies. Plants 2020, 9, 1469. [Google Scholar] [CrossRef]
- Bressan, M.; Blum, A.; Castel, L.; Trinsoutrot-Gattin, I.; Laval, K.; Gangneux, C. Assessment of Verticillium Flax Inoculum in Agroecosystem Soils Using Real-Time PCR Assay. Agric. Ecosyst. Environ. Appl. Soil Ecol. 2016, 108, 176–186. [Google Scholar] [CrossRef]
- Kanapin, A.; Samsonova, A.; Rozhmina, T.; Bankin, M.; Logachev, A.; Samsonova, M. The Genome Sequence of Five Highly Pathogenic Isolates of Fusarium Oxysporum f. Sp. Lini. Mol. Plant Microbe Interact. 2020, 33, 1112–1115. [Google Scholar] [CrossRef]
- Krasnov, G.S.; Pushkova, E.N.; Novakovskiy, R.O.; Kudryavtseva, L.P.; Rozhmina, T.A.; Dvorianinova, E.M.; Povkhova, L.V.; Kudryavtseva, A.V.; Dmitriev, A.A.; Melnikova, N.V. High-Quality Genome Assembly of Fusarium Oxysporum f. Sp. Lini. Front. Genet. 2020, 11, 959. [Google Scholar] [CrossRef] [PubMed]
- Dvorianinova, E.M.; Pushkova, E.N.; Novakovskiy, R.O.; Povkhova, L.V.; Bolsheva, N.L.; Kudryavtseva, L.P.; Rozhmina, T.A.; Melnikova, N.V.; Dmitriev, A.A. Nanopore and Illumina Genome Sequencing of Fusarium Oxysporum f. Sp. Lini Strains of Different Virulence. Front. Genet. 2021, 12, 662928. [Google Scholar] [CrossRef] [PubMed]
- Sigova, E.A.; Pushkova, E.N.; Rozhmina, T.A.; Kudryavtseva, L.P.; Zhuchenko, A.A.; Novakovskiy, R.O.; Zhernova, D.A.; Povkhova, L.V.; Turba, A.A.; Borkhert, E.V.; et al. Assembling Quality Genomes of Flax Fungal Pathogens from Oxford Nanopore Technologies Data. J. Fungi 2023, 9, 301. [Google Scholar] [CrossRef]
- Nemri, A.; Saunders, D.; Anderson, C.; Upadhyaya, N.; Win, J.; Lawrence, G.; Jones, D.; Kamoun, S.; Ellis, J.; Dodds, P. The Genome Sequence and Effector Complement of the Flax Rust Pathogen Melampsora Lini. Front. Plant Sci. 2014, 5, 98. [Google Scholar] [CrossRef]
- Anderson, C.; Khan, M.A.; Catanzariti, A.-M.; Jack, C.A.; Nemri, A.; Lawrence, G.J.; Upadhyaya, N.M.; Hardham, A.R.; Ellis, J.G.; Dodds, P.N.; et al. Genome Analysis and Avirulence Gene Cloning Using a High-Density RADseq Linkage Map of the Flax Rust Fungus, Melampsora Lini. BMC Genom. 2016, 17, 667. [Google Scholar] [CrossRef] [Green Version]
- Cui, Z.; Yan, B.; Gao, Y.; Wu, B.; Wang, Y.; Wang, H.; Xu, P.; Zhao, B.; Cao, Z.; Zhang, Y.; et al. Agronomic Cultivation Measures on Productivity of Oilseed Flax: A Review. Oil Crop Sci. 2022, 7, 53–62. [Google Scholar] [CrossRef]
- Lopez-Escudero, F.J.; Blanco-Lopez, M.A. Effect of a Single or Double Soil Solarization to Control Verticillium Wilt in Established Olive Orchards in Spain. Plant Dis. 2001, 85, 489–496. [Google Scholar] [CrossRef] [Green Version]
- Agrios, G.N. Control of Plant Diseases. In Plant Pathology; Elsevier: Amsterdam, The Netherlands, 2005; pp. 293–353. ISBN 978-0-12-044565-3. [Google Scholar]
- Ostos, E.; Garcia-Lopez, M.T.; Porras, R.; Lopez-Escudero, F.J.; Trapero-Casas, A.; Michailides, T.J.; Moral, J. Effect of Cultivar Resistance and Soil Management on Spatial–Temporal Development of Verticillium Wilt of Olive: A Long-Term Study. Front. Plant Sci. 2020, 11, 584496. [Google Scholar] [CrossRef]
- Ma, L.; Rocha, F.I.; Lee, J.; Choi, J.; Tejera, M.; Sooksa-Nguan, T.; Boersma, N.; VanLoocke, A.; Heaton, E.; Howe, A. The Impact of Stand Age and Fertilization on the Soil Microbiome of Miscanthus × Giganteus. Phytobiomes J. 2021, 5, 51–59. [Google Scholar] [CrossRef]
- Raaijmakers, J.M.; Mazzola, M. Soil Immune Responses. Science 2016, 352, 1392–1393. [Google Scholar] [CrossRef] [PubMed]
- Jayaraman, S.; Naorem, A.K.; Lal, R.; Dalal, R.C.; Sinha, N.K.; Patra, A.K.; Chaudhari, S.K. Disease-Suppressive Soils—Beyond Food Production: A Critical Review. J. Soil Sci. Plant Nutr. 2021, 21, 1437–1465. [Google Scholar] [CrossRef] [PubMed]
- Schlatter, D.; Kinkel, L.; Thomashow, L.; Weller, D.; Paulitz, T. Disease Suppressive Soils: New Insights from the Soil Microbiome. Phytopathology 2017, 107, 1284–1297. [Google Scholar] [CrossRef] [Green Version]
- Were, E.; Viljoen, A.; Rasche, F. Back to the Roots: Understanding Banana below-Ground Interactions Is Crucial for Effective Management of Fusarium Wilt. Plant Pathol. 2023, 72, 19–38. [Google Scholar] [CrossRef]
- Catambacan, D.G.; Cumagun, C.J.R. Weed-Associated Fungal Endophytes as Biocontrol Agents of Fusarium Oxysporum f. Sp. Cubense TR4 in Cavendish Banana. J. Fungi 2021, 7, 224. [Google Scholar] [CrossRef]
- Vinayarani, G.; Prakash, H.S. Fungal Endophytes of Turmeric (Curcuma Longa L.) and Their Biocontrol Potential against Pathogens Pythium Aphanidermatum and Rhizoctonia Solani. World J. Microbiol. Biotechnol. 2018, 34, 49. [Google Scholar] [CrossRef]
- Yuan, X.; Hong, S.; Xiong, W.; Raza, W.; Shen, Z.; Wang, B.; Li, R.; Ruan, Y.; Shen, Q.; Dini-Andreote, F. Development of Fungal-Mediated Soil Suppressiveness against Fusarium Wilt Disease via Plant Residue Manipulation. Microbiome 2021, 9, 200. [Google Scholar] [CrossRef]
- Palmieri, D.; Ianiri, G.; Del Grosso, C.; Barone, G.; De Curtis, F.; Castoria, R.; Lima, G. Advances and Perspectives in the Use of Biocontrol Agents against Fungal Plant Diseases. Horticulturae 2022, 8, 577. [Google Scholar] [CrossRef]
- Ons, L.; Bylemans, D.; Thevissen, K.; Cammue, B.P.A. Combining Biocontrol Agents with Chemical Fungicides for Integrated Plant Fungal Disease Control. Microorganisms 2020, 8, 1930. [Google Scholar] [CrossRef]
- Leach, J.E.; Leung, H.; Tisserat, N.A. Plant Disease and Resistance. In Encyclopedia of Agriculture and Food Systems; Elsevier: Amsterdam, The Netherlands, 2014; pp. 360–374. ISBN 978-0-08-093139-5. [Google Scholar]
- Robb, E.J.; Nazar, R.N. Tomato Ve-Resistance Locus: Resilience in the Face of Adversity? Planta 2021, 254, 126. [Google Scholar] [CrossRef]
- Zhang, Y.; Lubberstedt, T.; Xu, M. The Genetic and Molecular Basis of Plant Resistance to Pathogens. J. Genet. Genom. 2013, 40, 23–35. [Google Scholar] [CrossRef] [Green Version]
- Diederichsen, A.; Kusters, P.M.; Kessler, D.; Bainas, Z.; Gugel, R.K. Assembling a Core Collection from the Flax World Collection Maintained by Plant Gene Resources of Canada. Genet. Resour. Crop Evol. 2013, 60, 1479–1485. [Google Scholar] [CrossRef]
- Soto-Cerda, B.J.; Diederichsen, A.; Ragupathy, R.; Cloutier, S. Genetic Characterization of a Core Collection of Flax (Linum Usitatissimum L.) Suitable for Association Mapping Studies and Evidence of Divergent Selection between Fiber and Linseed Types. BMC Plant Biol. 2013, 13, 78. [Google Scholar] [CrossRef] [Green Version]
- Lawrence, G.J.; Anderson, P.A.; Dodds, P.N.; Ellis, J.G. Relationships between Rust Resistance Genes at the M Locus in Flax. Mol. Plant Pathol. 2010, 11, 19. [Google Scholar] [CrossRef] [PubMed]
- Haroon, M.; Wang, X.; Afzal, R.; Zafar, M.M.; Idrees, F.; Batool, M.; Khan, A.S.; Imran, M. Novel Plant Breeding Techniques Shake Hands with Cereals to Increase Production. Plants 2022, 11, 1052. [Google Scholar] [CrossRef] [PubMed]
- Asgarinia, P.; Cloutier, S.; Duguid, S.; Rashid, K.; Mirlohi, A.; Banik, M.; Saeidi, G. Mapping Quantitative Trait Loci for Powdery Mildew Resistance in Flax (Linum Usitatissimum L.). Crop Sci. 2013, 53, 2462–2472. [Google Scholar] [CrossRef] [Green Version]
- Mohamed, A.A.; El-Hefny, M.; El-Shanhorey, N.A.; Ali, H.M. Foliar Application of Bio-Stimulants Enhancing the Production and the Toxicity of Origanum Majorana Essential Oils Against Four Rice Seed-Borne Fungi. Molecules 2020, 25, 2363. [Google Scholar] [CrossRef]
- Shukla, P.S.; Borza, T.; Critchley, A.T.; Prithiviraj, B. Seaweed-Based Compounds and Products for Sustainable Protection against Plant Pathogens. Mar. Drugs 2021, 19, 59. [Google Scholar] [CrossRef]
- Pršić, J.; Ongena, M. Elicitors of Plant Immunity Triggered by Beneficial Bacteria. Front. Plant Sci. 2020, 11, 594530. [Google Scholar] [CrossRef]
- Rodríguez, A.B.F.; Menéndez, D.C.; Delgado, E.O.; Díaz, O.L.; Pino, J.C.C. Evaluation of Chitosan as an Inhibitor of Soil-Borne Pathogens and as an Elicitor of Defence Markers and Resistance in Tobacco Plants. Span. J. Agric. Res. 2007, 5, 533–541. [Google Scholar] [CrossRef] [Green Version]
- Saberi Riseh, R.; Hassanisaadi, M.; Vatankhah, M.; Soroush, F.; Varma, R.S. Nano/Microencapsulation of Plant Biocontrol Agents by Chitosan, Alginate, and Other Important Biopolymers as a Novel Strategy for Alleviating Plant Biotic Stresses. Int. J. Biol. Macromol. 2022, 222, 1589–1604. [Google Scholar] [CrossRef] [PubMed]




| Ranking | Country or Area | Total Production (ton) | Acreage (ha) | Crop Yield (ton/ha) |
|---|---|---|---|---|
| A | ||||
| 1 | Russia | 1,300,232 | 1,492,119 | 0.9 |
| 2 | Kazakhstan | 775,517 | 1,366,068 | 0.6 |
| 3 | Canada | 345,719 | 403,500 | 0.9 |
| 4 | China, mainland | 340,002 | 260,000 | 1.3 |
| 5 | India | 111,007 | 183,725 | 0.6 |
| 6 | Ethiopia | 82,000 | 80,000 | 1.0 |
| 7 | France | 72,941 | 37,500 | 1.9 |
| 8 | United Kingdom and Northern Ireland | 70,998 | 40,907 | 1.7 |
| 9 | United States of America | 68,785 | 108,460 | 0.6 |
| 10 | Ukraine | 42,231 | 27,600 | 1.5 |
| B | ||||
| 1 | France | 678,385 | 112,580 | 6.0 |
| 2 | Belgium | 87,000 | 15,390 | 5.7 |
| 3 | Belarus | 35,680 | 42,300 | 0.8 |
| 4 | China, mainland | 27,130 | 6317 | 4.3 |
| 5 | Russia | 25,947 | 36,483 | 0.7 |
| 6 | United Kingdom and Northern Ireland | 14,745 | 10,095 | 1.5 |
| 7 | The Netherlands | 11,330 | 1800 | 6.3 |
| 8 | Egypt | 7601 | 8609 | 0.9 |
| 9 | Chile | 3065 | 2769 | 1.1 |
| 10 | Argentina | 2620 | 2904 | 0.9 |
| Disease Common Name | Causal Agent | NCBI Taxonomy ID EPPO Code | Lifestyle | Affected Organs * | References | |
|---|---|---|---|---|---|---|
| Phylum | Species | |||||
| Anthracnose | Ascomycota | Colletotrichum lini (Colletotrichum linicola) | 500171 COLLLI | Hemibiotrophic | Seedling Leaf Stem Buds/Flowers | [1,10,11,12,13] |
| Alternaria blight | Ascomycota | Alternaria linicola | 75319 ALTELI | Necrotrophic | Seedling Stem Bolls/Seeds | [1,14] |
| Browning and stem break | Ascomycota | Aureobasidium pullulans var. lini (Kabatiella lini, Polyspora lini, Discosphaerina fulvida, Guignardia fulvida) | 1836197 AUREPL | Necrotrophic | Seedling Stem | [1,13,15] |
| Foot rot/basal stem blight | Ascomycota | Boeremia linicola (Phoma exigua f. sp. linicola) | 2904145 PHOMEL | Necrotrophic | Root Seedling Stem Bolls/Seeds | [1,16] |
| Fusarium wilt | Ascomycota | Fusarium oxysporum f. sp. lini | 120040 FUSALI | Hemibiotrophic | Seedling Leaf Stem Buds/Flowers | [1,11,17,18] |
| Gray mold | Ascomycota | Botrytis cinerea | 40559 BOTRCI | Necrotrophic | Seedling Bolls/Seeds | [1,11,19,20] |
| Pasmo | Ascomycota | Septoria linicola (Mycosphaerella linicola) | 215465 MYCOLN | Hemibiotrophic | Seedling Leaf Stem Bolls/Seeds | [1,21,22,23,24] |
| Powdery mildew | Ascomycota | Podosphaera lini (Oidium lini), Golovinomyces orontii and others | 683379 62715 ERYSPP | Biotrophic | Leaf Stem | [1,25] |
| Rust | Basidiomycota | Melampsora lini | 5261 MELMLI | Biotrophic | Leaf Stem Bolls/Seeds | [1,11] |
| Sclerotinia stem rot | Ascomycota | Sclerotinia sclerotiorum | 5180 SCLESC | Necrotrophic | Stem | [1,26] |
| Scorch | Oomycota Ascomycota | Globisporangium megalacanthum (Pythium megalacanthum) and Berkeleyomyces basicola (Thielaviopsis basicola, Chalara elegans) | 147705 PYTHME 124036 THIEBA | Necrotrophic Hemibiotrophic | Root Seedling Leaf Stem | [1,27] |
| Seedling blight/root rot | Basidiomycota | Rhizoctonia solani | 456999 RHIZSO | Necrotrophic | Root Seedling Stem | [1,28,29] |
| Verticillium wilt | Ascomycota | Verticillium dahliae | 27337 VERTDA | Hemibiotrophic | Leaf Stem Windrow | [1,12,30] |
| FRAC Acknowledged Mode of Action— Target Site Code | Chemical Group— FRAC Code | Active Substance | Resistance Risk | Target Disease * | Areas of Authorization |
|---|---|---|---|---|---|
| A | |||||
| Contact fungicide action | |||||
| Chemicals with multi-site activity | Inorganic—M02 | Sulfur | Low | Powdery mildew | BE |
| Data not available | Inorganic | Potassium hydrogen carbonate | Data not available | Powdery mildew | NL |
| Signal transduction—E2 | Phenylpyrroles—12 | Fludioxonil a | Low to medium | Gray mold | NL |
| Unknown | Phenyl-acetamides—U06 | Cyflufenamid | Unknown | Powdery mildew | BE, FR |
| Systemic fungicide action | |||||
| Cytoskeleton and motor protein—B1 | MBC—Thiophanate—1 | Thiophanate-methyl | High | Anthracnose Fusarium wilt Pasmo | BY |
| Respiration—C2 | SDHI—pyrazole-4-carboxamides—7 | Benzovindiflupyr b | Medium to high | Pasmo Powdery mildew | GB, IE |
| Bixafen | Sclerotinia stem rot | NL, CA | |||
| Fluxapyroxad c | |||||
| SDHI—pyridine-carboxamides—7 | Boscalid | Medium to high | Basal stem blight Foot rot | FR | |
| Powdery mildew | NL | ||||
| SDHI—pyridinyl-ethyl-benzamides—7 | Fluopyram d | Medium to high | Gray mold Sclerotinia stem rot | NL | |
| Respiration—C3 | QoI—methoxy-acrylates—11 | Azoxystrobin e | High | Anthracnose | BY |
| Browning and stem break | BY, FR | ||||
| Fusarium wilt | BY | ||||
| Pasmo | BY, CA, FR | ||||
| Powdery mildew | CA, FR | ||||
| Sclerotinia stem rot | CA, FR, NL | ||||
| Picoxystrobin | High | Pasmo | CA | ||
| QoI- methoxy-carbamates—11 | Pyraclostrobin c | High | Anthracnose | BY | |
| Fusarium wilt | BY | ||||
| Pasmo | BY, CA | ||||
| Powdery mildew | NL | ||||
| Sclerotinia stem rot | CA | ||||
| QoI—oximino-acetates—11 | Trifloxystrobin | High | Alternaria blight Powdery mildew | NL | |
| Amino acid and protein synthesis—D1 | AP—anilino-pyrimidines—9 | Cyprodinil a | Medium | Gray mold | NL |
| Sterol biosynthesis in membranes—G1 | DMI—Triazoles—3 | Cyproconazole | Medium | Anthracnose Fusarium wilt Pasmo | BY |
| Difenoconazole e | Medium | Anthracnose | BY | ||
| Basal stem blight | FR | ||||
| Browning and stem break | FR | ||||
| Foot rot | FR | ||||
| Fusarium wilt | BY | ||||
| Pasmo | BY, CA, FR | ||||
| Powdery mildew | BE, CA, FR, NL | ||||
| Sclerotinia stem rot | CA, FR | ||||
| Epoxiconazole | Medium | Anthracnose Fusarium wilt Pasmo | BY | ||
| Systemic fungicide action | |||||
| Sterol biosynthesis in membranes—G1 | DMI—Triazoles—3 | Flutriafol | Medium | Anthracnose Fusarium wilt Pasmo | BY |
| Propiconazole | Medium | Anthracnose Browning and stem break Pasmo | BY | ||
| Spiroxamine | Low to medium | Anthracnose Fusarium wilt | BY | ||
| Tebuconazole d | Medium | Anthracnose | BY | ||
| Browning and stem break | FR | ||||
| Fusarium wilt | BY | ||||
| Gray mold | GB, IE, NL | ||||
| Pasmo | FR | ||||
| Powdery mildew | BE, FR | ||||
| Sclerotinia stem rot | NL | ||||
| DMI—Triazolinthione—3 | Prothioconazole b | Medium | Anthracnose | BY | |
| Fusarium wilt | BY | ||||
| Pasmo | GB, IE | ||||
| Powdery mildew | BE, FR, GB, IE | ||||
| Sclerotinia stem rot | CA | ||||
| B | |||||
| Contact fungicide action | |||||
| signal transduction—E2 | Phenylpyrroles—12 | Fludioxonil | Low to medium | Alternaria blight | BE, NL |
| Anthracnose | NL | ||||
| Basal stem blight | BE, NL | ||||
| Foot rot | BE, NL | ||||
| Fusarium wilt | BE, FR, NL | ||||
| Gray mold | BE | ||||
| Sclerotinia stem rot | BE, NL | ||||
| Seedling blight | NL | ||||
| chemicals with multi-site activity | Dithiocarbamates—M03 | Thiram f | Low | Fusarium wilt Seedling blight | CA |
| Systemic fungicide action | |||||
| nucleic acid metabolism—A1 | PA—Acylalanines—4 | Metalaxyl-M | High | Alternaria blight | NL |
| Anthracnose | NL | ||||
| Basal stem blight | NL | ||||
| Foot rot | NL | ||||
| Sclerotinia stem rot | NL | ||||
| Scorch | BE, FR | ||||
| Seedling blight | NL | ||||
| cytoskeleton and motor protein—B1 | MBC—Benzimidazoles—1 | Carbendazim | High | Anthracnose Fusarium wilt Pasmo | BY |
| respiration—C2 | SDHI—Oxathiin-carboxamides—7 | Carboxin a | Medium to high | Fusarium wilt Seedling blight | CA |
| Causal Agent (Strain) | Cultivar/Type | Plant Cultivation Details | Disease Assessment Details | Technique—Results | Chr Involved * | Reference |
|---|---|---|---|---|---|---|
| Alternaria blight Alternaria linicola | F2 population (disease-resistant genotype × disease-susceptible genotype) | Field (India)—2014 | Disease rating Score: 0–5 Stages: flowering harvesting | Simple sequence repeats (SSR) genotyping—2 QTLs | 14 | [51] |
| Fusarium wilt Fusarium oxysporum f.sp. lini (MI39) | Russian flax genetic collection (179 fiber flax accessions and 117 linseed accessions) | Greenhouse—3 years in a row (2019–2021)—Infection of soil using pure culture of fungus | Disease Severity Index Score: 0–3 Stage: early yellow ripeness | GWAS— 15 QTNs | 1, 8, 11, 13 | [17,48] |
| Powdery mildew Oïdium lini | F3 and F4 populations (disease-resistant genotype × disease-susceptible genotype) | F3: field (Canada, 2012), under natural powdery mildew infection F4: growth chamber, infection with isolate PM97 propagated on flax plants in controlled conditions | Disease rating Score: 0–9 Stages: F3: end of flowering/mid- and late green boll F4: pre-flowering | Simple sequence repeats (SSR) genotyping—3 QTLs | 1, 7, 9 | [123] |
| Worldwide collection (38 countries) 173 fiber flax 110 linseed 2 wild flax 19 unknown type flax | Field (France)—2018 and 2019 Natural field infection | Disease rating with AUDPC Score: 0–9 Stage: flowering | GWAS—10 QTLs | 1, 2, 4, 13, 14 | [38] | |
| Worldwide collection (39 countries) 80 fiber flax 292 linseed | Field (Canada)—2012 to 2016 Greenhouse-inoculated diseased plants (10) every ten rows | Disease rating Score: 0–9 Stages: early flowering (8 to 9 weeks after sowing) late flowering (9 to 10 weeks after sowing) green boll (10 to 11 weeks after sowing) early brown boll (11 to 12 weeks after sowing) | GWAS— 349 QTNs | All | [37,48] | |
| Pasmo Septoria linicola | Field (Canada)—2012 to 2016 Spread of infested chopped straw between rows and misting | GWAS— 692 QTNs | All | [48,58,60] |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Moyse, J.; Lecomte, S.; Marcou, S.; Mongelard, G.; Gutierrez, L.; Höfte, M. Overview and Management of the Most Common Eukaryotic Diseases of Flax (Linum usitatissimum). Plants 2023, 12, 2811. https://0-doi-org.brum.beds.ac.uk/10.3390/plants12152811
Moyse J, Lecomte S, Marcou S, Mongelard G, Gutierrez L, Höfte M. Overview and Management of the Most Common Eukaryotic Diseases of Flax (Linum usitatissimum). Plants. 2023; 12(15):2811. https://0-doi-org.brum.beds.ac.uk/10.3390/plants12152811
Chicago/Turabian StyleMoyse, Julie, Sylvain Lecomte, Shirley Marcou, Gaëlle Mongelard, Laurent Gutierrez, and Monica Höfte. 2023. "Overview and Management of the Most Common Eukaryotic Diseases of Flax (Linum usitatissimum)" Plants 12, no. 15: 2811. https://0-doi-org.brum.beds.ac.uk/10.3390/plants12152811