
Effects of Spring Drought and Nitrogen Addition on Productivity and Community Composition of Degraded Grasslands
 and
and {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
2.1. Responses of Soil Moisture
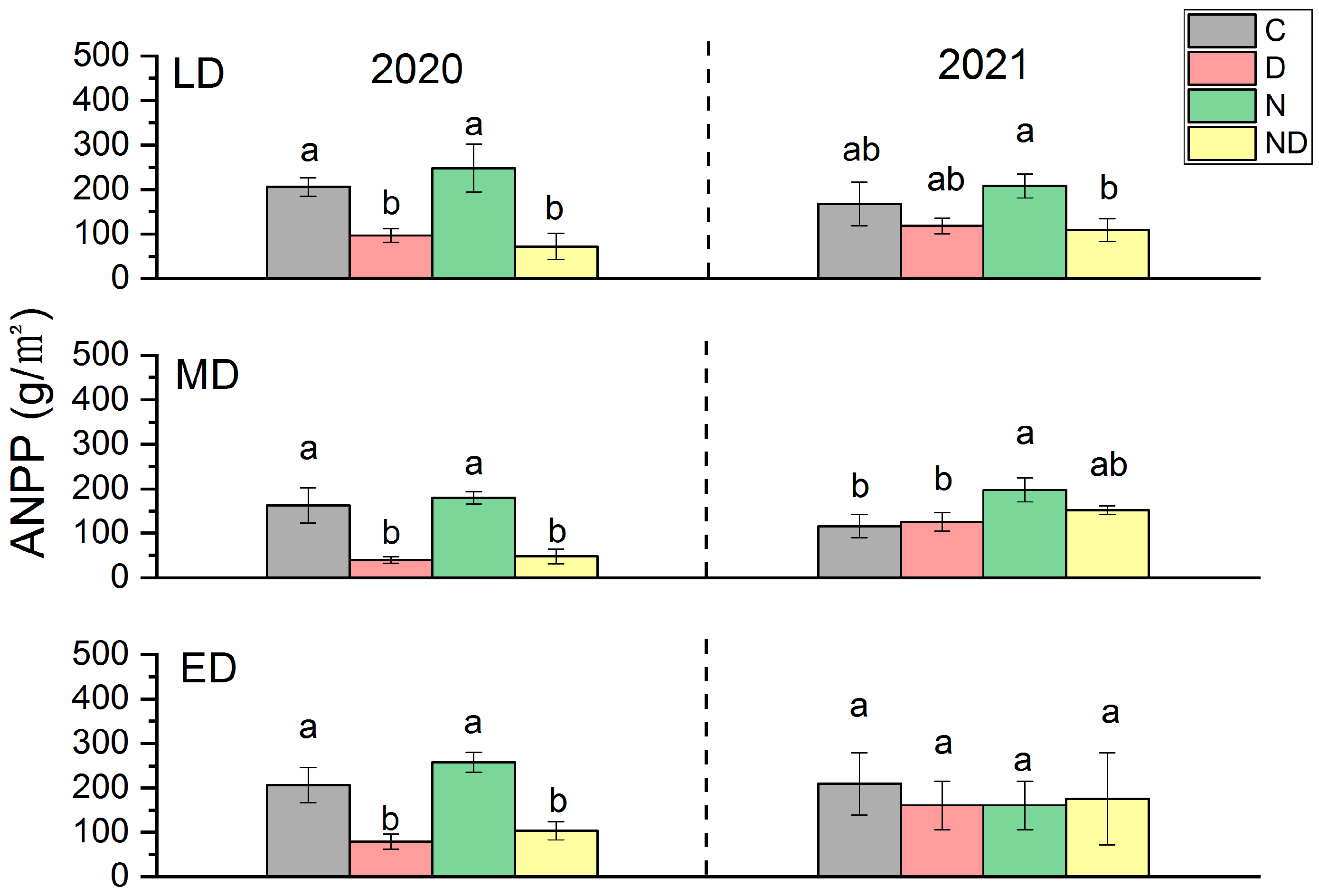
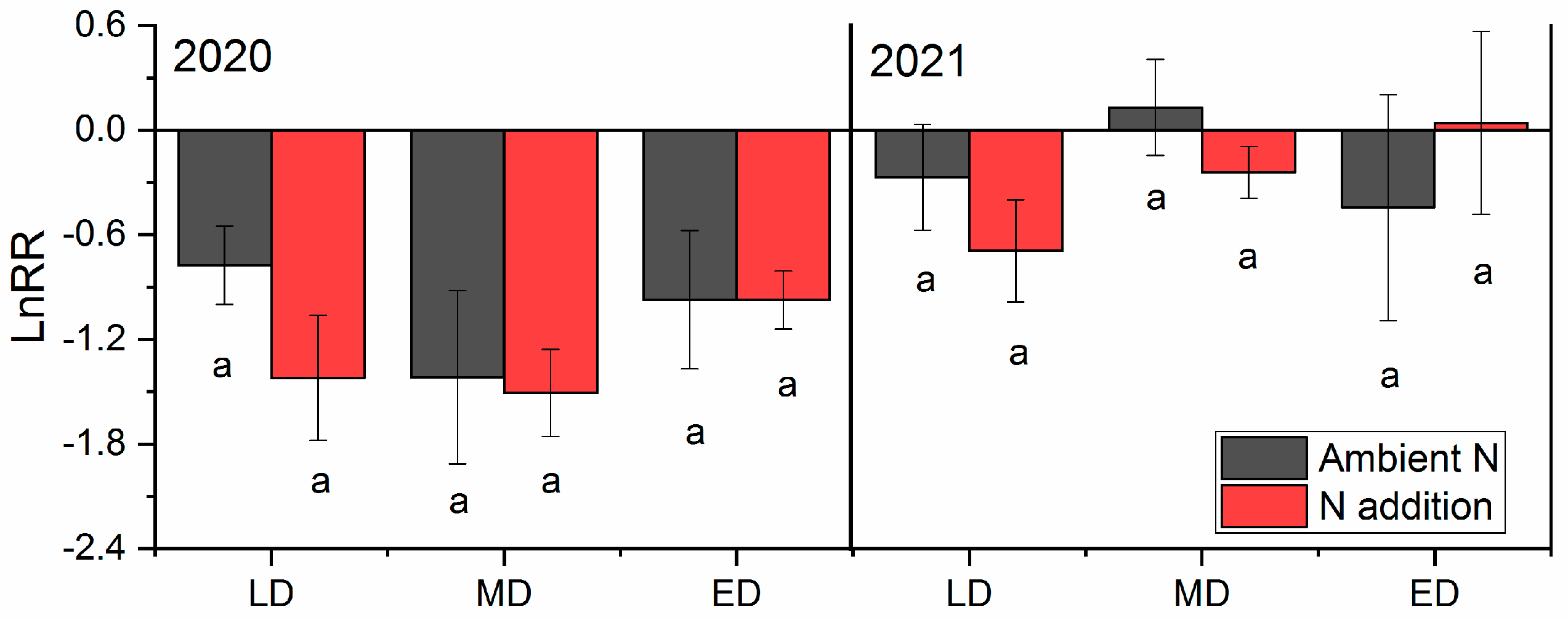
2.2. Responses of ANPP
2.3. Responses of the Composition of Vegetation from the Functional Groups and Degradation of the Indicator Species
2.4. Relationships between ANPP Drought Sensitivity, the Composition of Vegetation of Plant Functional Groups, and Degradation Indicator Species
3. Discussion
3.1. Effects of the Interactions of Spring Drought and Nitrogen Addition on the ANPP of Degraded Grasslands
3.2. Regulation of Drought Sensitivity of Degraded Grassland by Plant Functional Groups and Degradation Indicator Species
3.3. Suggestions on Grassland Restoration and Management
4. Conclusions
5. Materials and Methods
5.1. Study Areas
5.2. Sample Site Selection
5.3. Experimental Design
5.4. Data Collection
5.5. Statistical Analysis
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Kang, L.; Han, X.; Zhang, Z.; Sun, O.J. Grassland ecosystems in China: Review of current knowledge and research advancement. Philos. Trans. R. Soc. B Biol. Sci. 2007, 362, 997–1008. [Google Scholar] [CrossRef] [PubMed]
- Bardgett, R.D.; Bullock, J.M.; Lavorel, S.; Manning, P.; Schaffner, U.; Ostle, N.; Chomel, M.; Durigan, G.; Fry, E.L.; Johnson, D.; et al. Combatting global grassland degradation. Nat. Rev. Earth Environ. 2021, 2, 720–735. [Google Scholar] [CrossRef]
- Dong, C.C.; Wang, W.; Liu, H.Y.; Xu, X.T.; Zeng, H. Temperate grassland shifted from nitrogen to phosphorus limitation induced by degradation and nitrogen deposition: Evidence from soil extracellular enzyme stoichiometry. Ecol. Indic. 2019, 101, 453–464. [Google Scholar] [CrossRef]
- Dietrich, J.D.; Smith, M.D. The effect of timing of growing season drought on flowering of a dominant C4 grass. Oecologia 2016, 181, 391–399. [Google Scholar] [CrossRef] [PubMed]
- Ren, H.; Xu, Z.; Isbell, F.; Huang, J.; Han, X.; Wan, S.; Chen, S.; Wang, R.; Zeng, D.-H.; Jiang, Y.; et al. Exacerbated nitrogen limitation ends transient stimulation of grassland productivity by increased precipitation. Ecol. Monogr. 2017, 87, 457–469. [Google Scholar] [CrossRef]
- Griffin-Nolan, R.J.; Carroll, C.J.W.; Denton, E.M.; Johnston, M.K.; Collins, S.L.; Smith, M.D.; Knapp, A.K. Legacy effects of a regional drought on aboveground net primary production in six central US grasslands. Plant Ecol. 2018, 219, 505–515. [Google Scholar] [CrossRef]
- Harrison, S.; LaForgia, M. Seedling traits predict drought-induced mortality linked to diversity loss. Proc. Natl. Acad. Sci. USA 2019, 116, 5576–5581. [Google Scholar] [CrossRef] [Green Version]
- Hoover, D.L.; Duniway, M.C.; Belnap, J. Testing the apparent resistance of three dominant plants to chronic drought on the Colorado Plateau. J. Ecol. 2017, 105, 152–162. [Google Scholar] [CrossRef] [Green Version]
- Gessler, A.; Schaub, M.; McDowell, N.G. The role of nutrients in drought—Induced tree mortality and recovery. New Phytol. 2017, 214, 513–520. [Google Scholar] [CrossRef] [Green Version]
- Knapp, A.K.; Fay, P.A.; Blair, J.M.; Collins, S.L.; Smith, M.D.; Carlisle, J.D.; Harper, C.W.; Danner, B.T.; Lett, M.S.; McCarron, J.K. Rainfall Variability, Carbon Cycling, and Plant Species Diversity in a Mesic Grassland. Science 2002, 298, 2202–2205. [Google Scholar] [CrossRef] [Green Version]
- Knapp, A.K.; Ciais, P.; Smith, M.D. Reconciling inconsistencies in precipitation–productivity relationships: Implications for climate change. New Phytol. 2017, 214, 41–47. [Google Scholar] [CrossRef] [Green Version]
- Bobbink, R.; Hicks, K.; Galloway, J.; Spranger, T.; Alkemade, R.; Ashmore, M.; Bustamante, M.; Cinderby, S.; Davidson, E.; Dentener, F.; et al. Global assessment of nitrogen deposition effects on terrestrial plant diversity: A synthesis. Ecol. Appl. 2010, 20, 30–59. [Google Scholar] [CrossRef] [Green Version]
- Ma, Y.; Xu, Z.; Zeng, Q.; Meng, J.; Hu, Y.; Su, J. Effects of nitrogen addition on nutrient stoichiometric characteristics of herbaceous plants in deser-tification steppe. Acta Prataculturae Sin. 2021, 30, 64–72. (In Chinese) [Google Scholar]
- Bai, Y.; Wu, J.; Clark, C.M.; Naeem, S.; Pan, Q.; Huang, J.; Zhang, L.; Han, X. Tradeoffs and thresholds in the effects of nitrogen addition on biodiversity and ecosystem functioning: Evidence from inner Mongolia Grasslands. Glob. Chang. Biol. 2010, 16, 358–372. [Google Scholar] [CrossRef]
- Isbell, F.; Reich, P.B.; Tilman, D.; Hobbie, S.E.; Polasky, S.; Binder, S. Nutrient enrichment, biodiversity loss, and consequent declines in ecosystem productivity. Proc. Natl. Acad. Sci. USA 2013, 110, 11911–11916. [Google Scholar] [CrossRef]
- Xu, X.; Liu, H.; Song, Z.; Wang, W.; Hu, G.; Qi, Z. Response of aboveground biomass and diversity to nitrogen addition along a degradation gradient in the Inner Mongolian steppe, China. Sci. Rep. 2015, 5, 10284. [Google Scholar] [CrossRef] [Green Version]
- Bharath, S.; Borer, E.T.; Biederman, L.A.; Blumenthal, D.M.; Fay, P.A.; Gherardi, L.A.; Knops, J.M.H.; Leakey, A.D.B.; Yahdjian, L.; Seabloom, E.W. Nutrient addition increases grassland sensitivity to droughts. Ecology 2020, 101, e02981. [Google Scholar] [CrossRef] [Green Version]
- Xu, Q.; Yang, X.; Song, J.; Ru, J.; Xia, J.; Wang, S.; Wan, S.; Jiang, L. Nitrogen enrichment alters multiple dimensions of grassland functional stability via changing compositional stability. Ecol. Lett. 2022, 25, 2713–2725. [Google Scholar] [CrossRef]
- Bloor, J.M.; Bardgett, R.D. Stability of above-ground and below-ground processes to extreme drought in model grassland ecosystems: Interactions with plant species diversity and soil nitrogen availability. Perspect. Plant Ecol. Evol. Syst. 2012, 14, 193–204. [Google Scholar] [CrossRef]
- Nogueira, C.; Werner, C.; Rodrigues, A.; Caldeira, M.C. A prolonged dry season and nitrogen deposition interactively affect CO2 fluxes in an annual Mediterranean grassland. Sci. Total. Environ. 2019, 654, 978–986. [Google Scholar] [CrossRef]
- Zhang, Y.; Ren, Z.; Zhang, Y. Winter nitrogen enrichment does not alter the sensitivity of plant communities to precipitation in a semiarid grassland. Sci. Total Environ. 2021, 790, 148264. [Google Scholar] [CrossRef]
- Shi, B.; Wang, Y.; Meng, B.; Zhong, S.; Sun, W. Effects of nitrogen addition on the drought susceptibility of the Leymus chinensis meadow eco-system vary with drought duration. Front. Plant Sci. 2018, 9, 254. [Google Scholar] [CrossRef]
- Zhang, X.; Tang, H.; Dong, X.; Li, B.; Huang, Y.; Gong, J. The dilemma of Chinese grassland and its transformation. Chin. Sci. Bull. 2016, 61, 165–177. (In Chinese) [Google Scholar]
- Berdugo, M.; Delgado-Baquerizo, M.; Soliveres, S.; Hernández-Clemente, R.; Zhao, Y.; Gaitán, J.J.; Gross, N.; Saiz, H.; Maire, V.; Lehmann, A.; et al. Global ecosystem thresholds driven by aridity. Science 2020, 367, 787–790. [Google Scholar] [CrossRef] [Green Version]
- Luo, W.; Muraina, T.O.; Griffin-Nolan, R.J.; Ma, W.; Song, L.; Fu, W.; Yu, Q.; Knapp, A.K.; Wang, Z.; Han, X.; et al. Responses of a semiarid grassland to recurrent drought are linked to community functional composition. Ecology 2022, 104, e3920. [Google Scholar] [CrossRef]
- Han, Q.; Li, Y.; Peng, K.; Li, C.; Huang, X.; Xu, W. Effects of atmospheric nitrogen deposition on net primary productivity of grassland ecosystems in Central Asia. Acta Ecol. Sin. 2021, 41, 8545–8555. (In Chinese) [Google Scholar]
- Chen, Z.; Lu, S.; Li, S.; Zhao, N.; Xu, X. Resistance and resilience of different plant functional groups to simulated drought in temperate meadow steppe. Chin. J. Grassl. 2023, 45, 10–19. (In Chinese) [Google Scholar]
- You, C.; Wu, F.; Gan, Y.; Yang, W.; Hu, Z.; Xu, Z.; Tan, B.; Liu, L.; Ni, X. Grass and forbs respond differently to nitrogen addition: A meta-analysis of global grassland eco-systems. Sci. Rep. 2017, 7, 1563. [Google Scholar] [CrossRef] [Green Version]
- Stampfli, A.; Bloor, J.M.G.; Fischer, M.; Zeiter, M. High land-use intensity exacerbates shifts in grassland vegetation composition after severe experimental drought. Glob. Chang. Biol. 2018, 24, 2021–2034. [Google Scholar] [CrossRef] [Green Version]
- Xu, X.; Hu, G.; Liu, X.; Lu, S.; Li, S.; Zhao, N. Impacts of nitrogen enrichment on vegetation growth dynamics are regulated by grassland degradation status. Land Degrad. Dev. 2021, 32, 4056–4066. [Google Scholar] [CrossRef]
- Meng, B.; Shi, B.; Zhong, S.; Chai, H.; Li, S.; Wang, Y.; Henry, H.A.L.; Ma, J.-Y.; Sun, W. Drought sensitivity of aboveground productivity in Leymus chinensis meadow steppe depends on drought timing. Oecologia 2019, 191, 685–696. [Google Scholar] [CrossRef]
- Huxman, T.E.; Smith, M.D.; Fay, P.A.; Knapp, A.K.; Shaw, M.R.; Loik, M.E.; Smith, S.D.; Tissue, D.T.; Zak, J.C.; Weltzin, J.F.; et al. Convergence across biomes to a common rain-use efficiency. Nature 2004, 429, 651–654. [Google Scholar] [CrossRef]
- Hofer, D.; Suter, M.; Buchmann, N.; Lüscher, A. Nitrogen status of functionally different forage species explains resistance to severe drought and post-drought overcompensation. Agric. Ecosyst. Environ. 2017, 236, 312–322. [Google Scholar] [CrossRef]
- Friedrich, U.; von Oheimb, G.; Kriebitzsch, W.U.; Schleßelmann, K.; Weber, M.S.; Härdtle, W. Nitrogen deposition increases susceptibility to drought-experimental evi-dence with the perennial grass Molinia caerulea (L.) Moench. Plant and Soil 2012, 353, 59–71. [Google Scholar] [CrossRef]
- Walter, J.; Jentsch, A.; Beierkuhnlein, C.; Kreyling, J. Ecological stress memory and cross stress tolerance in plants in the face of climate extremes. Environ. Exp. Bot. 2013, 94, 3–8. [Google Scholar] [CrossRef]
- Müller, L.M.; Bahn, M. Drought legacies and ecosystem responses to subsequent drought. Glob. Chang. Biol. 2022, 28, 5086–5103. [Google Scholar] [CrossRef]
- Li, X.; Jimoh, S.O.; Li, Y.; Duan, J.; Cui, Y.; Jin, K.; Wang, Z.; Zhang, Y. Stress memory and phyllosphere/soil legacy underlie tolerance and plasticity of Leymus chinensis to periodic drought risk. Agric. For. Meteorol. 2022, 312, 108717. [Google Scholar] [CrossRef]
- De Boeck, H.J.; Hiltbrunner, E.; Verlinden, M.; Bassin, S.; Zeiter, M. Legacy Effects of Climate Extremes in Alpine Grassland. Front. Plant Sci. 2018, 9, 1586. [Google Scholar] [CrossRef]
- Wilcox, K.R.; Blair, J.M.; Smith, M.D.; Knapp, A.K. Does ecosystem sensitivity to precipitation at the site-level conform to regional-scale predictions? Ecology 2016, 97, 561–568. [Google Scholar] [CrossRef]
- Xu, C.; Ke, Y.; Zhou, W.; Luo, W.; Ma, W.; Song, L.; Smith, M.D.; Hoover, D.L.; Wilcox, K.R.; Fu, W.; et al. Resistance and resilience of a semi-arid grassland to multi-year extreme drought. Ecol. Indic. 2021, 131, 108139. [Google Scholar] [CrossRef]
- Van Sundert, K.; Khan, M.A.S.A.; Bharath, S.; Buckley, Y.M.; Caldeira, M.C.; Donohue, I.; Dubbert, M.; Ebeling, A.; Eisenhauer, N.; Eskelinen, A.; et al. Fertilized graminoids intensify negative drought effects on grassland productivity. Glob. Chang. Biol. 2021, 27, 2441–2457. [Google Scholar] [CrossRef]
- Grime, J.P. Vegetation classification by reference to strategies. Nature 1974, 250, 26–31. [Google Scholar] [CrossRef]
- Zhang, Y.; Wang, R.; Sardans, J.; Wang, B.; Gu, B.; Li, Y.; Liu, H.; Peñuelas, J.; Jiang, Y. Resprouting ability differs among plant functional groups along a soil acidification gradient in a meadow: A rhizosphere perspective. J. Ecol. 2023, 111, 631–644. [Google Scholar] [CrossRef]
- Tang, H.; Liu, R. List of C4 plants in Inner Mongolia. J. Inn. Mong. Univ. (Nat. Sci. Ed.) 2001, 32, 431–438. (In Chinese) [Google Scholar]
- Smith, M.D. The ecological role of climate extremes: Current understanding and future prospects. J. Ecol. 2011, 99, 651–655. [Google Scholar] [CrossRef]
- Wang, R.; Tian, Y.; Ouyang, S.; Xu, X.; Xu, F.; Zhang, Y. Nitrogen acquisition strategies used by Leymus chinensis and Stipa grandis in temperate steppes. Biol. Fertil. Soils 2016, 52, 951–961. [Google Scholar] [CrossRef]
- Xu, X.; Liu, H.; Liu, X.; Song, Z.; Wang, W.; Qiu, S. Differentiated seasonal vegetation cover dynamics of degraded grasslands in Inner Mongolia recorded by continuous photography technique. Int. J. Biometeorol. 2019, 63, 671–677. [Google Scholar] [CrossRef]
- Wang, M. Ecological vulnerability of dam in Hebei province and suggestions on its treatment. Econ. Forum 2018, 10, 11–15. (In Chinese) [Google Scholar]
- Liu, H.; Yin, Y.; Tian, Y.; Ren, J.; Wang, H. Climatic and anthropogenic controls of topsoil features in the semi-arid East Asian steppe Grassland. Geophys. Res. Lett. 2008, 35, 1–17. [Google Scholar] [CrossRef]
- Li, B. The rangeland degradation in north China and its preventive strategy. Sci. Agric. Sin. 1997, 6, 2–10. (In Chinese) [Google Scholar]
- Inner Mongolia Ningxia Comprehensive Investigation Team of Chinese Academy of Sciences. Inner Mongolia Vegetation; Science Press: Beijing, China, 1985. (In Chinese) [Google Scholar]
- Zhao, N.; Li, S.; Lu, S.; Wang, X.; Chen, Z.; Xu, X. High resistance to spring drought of grasslands under different degradation levels on a Leymus chinensis steppe. Glob. Ecol. Conserv. 2023, 43, e02471. [Google Scholar] [CrossRef]
- Xu, X.; Liu, H.; Wang, W.; Song, Z. Patterns and determinants of the response of plant biomass to addition of nitrogen in semi-arid and alpine grasslands of China. J. Arid. Environ. 2018, 153, 11–17. [Google Scholar] [CrossRef]
- Zhao, Y.; Song, Z.; Xu, X.; Liu, H.; Wu, X.; Li, Z.; Guo, F.; Pan, W. Nitrogen application increases phytolith carbon sequestration in degraded grasslands of North China. Ecol. Res. 2016, 31, 117–123. [Google Scholar] [CrossRef]
- Xu, X.; Liu, H.; Wang, W.; Hu, G.; Wu, X.; Song, Z. Effects of manipulated precipitation on aboveground net primary productivity of grassland fields: Controlled rainfall experiments in Inner Mongolia, China. Land Degrad. Dev. 2021, 32, 1981–1992. [Google Scholar] [CrossRef]






Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Li, S.; Lu, S.; Li, X.; Hou, X.; Zhao, X.; Xu, X.; Zhao, N. Effects of Spring Drought and Nitrogen Addition on Productivity and Community Composition of Degraded Grasslands. Plants 2023, 12, 2836. https://0-doi-org.brum.beds.ac.uk/10.3390/plants12152836
Li S, Lu S, Li X, Hou X, Zhao X, Xu X, Zhao N. Effects of Spring Drought and Nitrogen Addition on Productivity and Community Composition of Degraded Grasslands. Plants. 2023; 12(15):2836. https://0-doi-org.brum.beds.ac.uk/10.3390/plants12152836
Chicago/Turabian StyleLi, Shaoning, Shaowei Lu, Xiaohui Li, Xingchen Hou, Xi Zhao, Xiaotian Xu, and Na Zhao. 2023. "Effects of Spring Drought and Nitrogen Addition on Productivity and Community Composition of Degraded Grasslands" Plants 12, no. 15: 2836. https://0-doi-org.brum.beds.ac.uk/10.3390/plants12152836