
Phytochemical and Agronomic Characterization of High-Flavonoid Lettuce Lines Grown under Field Conditions


Abstract
:1. Introduction
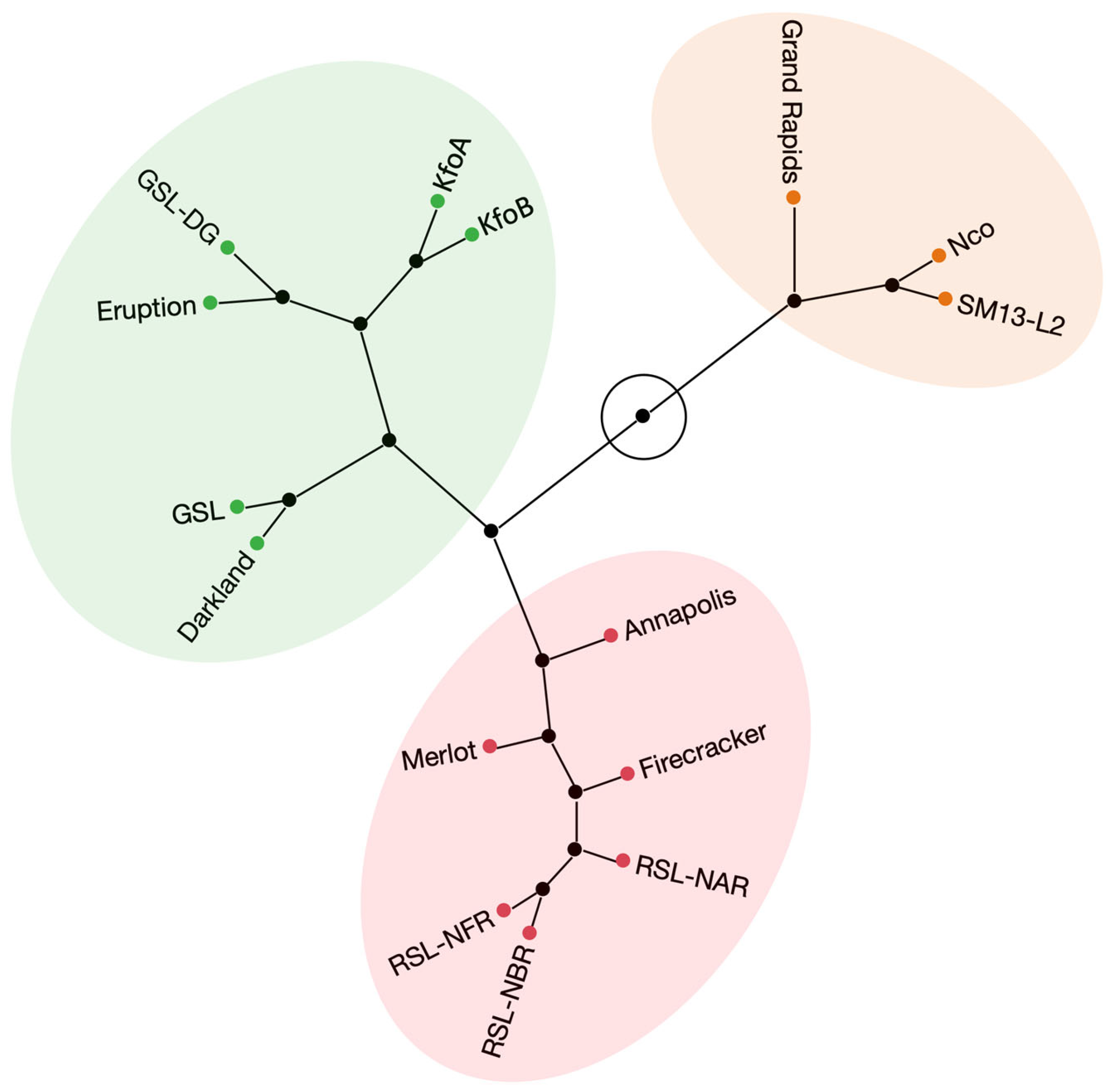
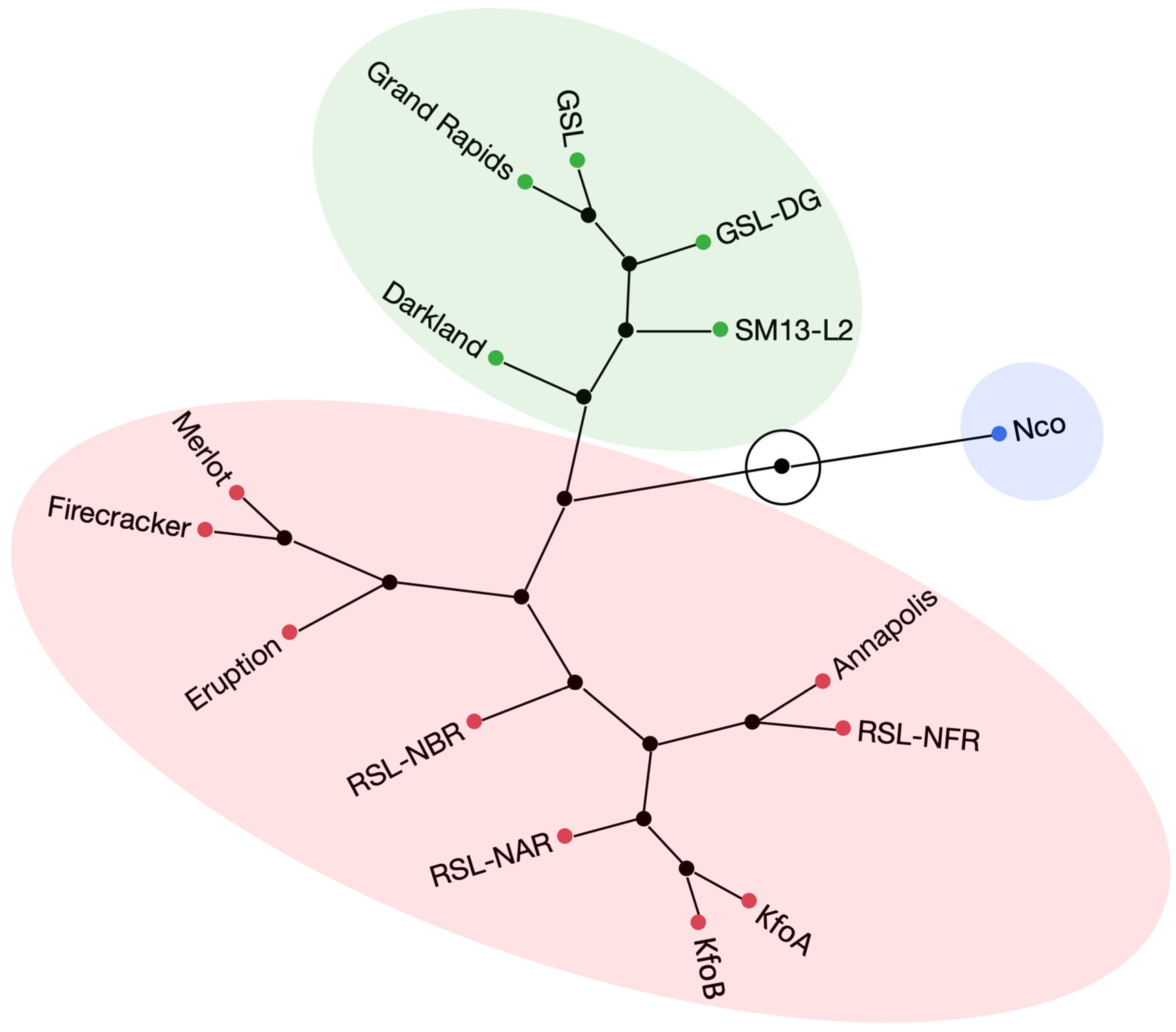
2. Results
2.1. Biomass Production
2.2. Content of Pigments in Leaves
2.3. Resistance to Diseases
2.4. Postharvest Quality
2.5. Elemental Composition
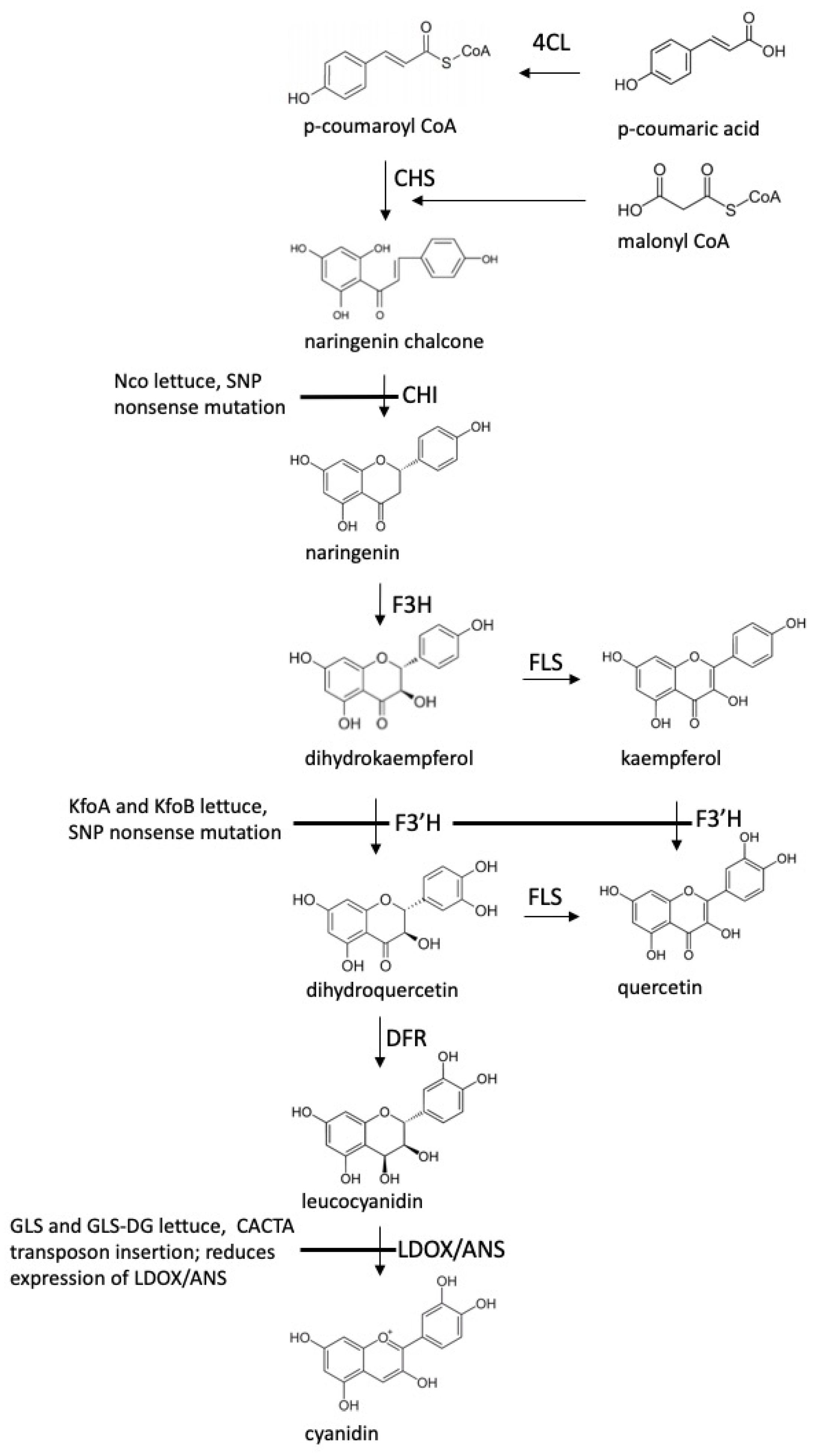
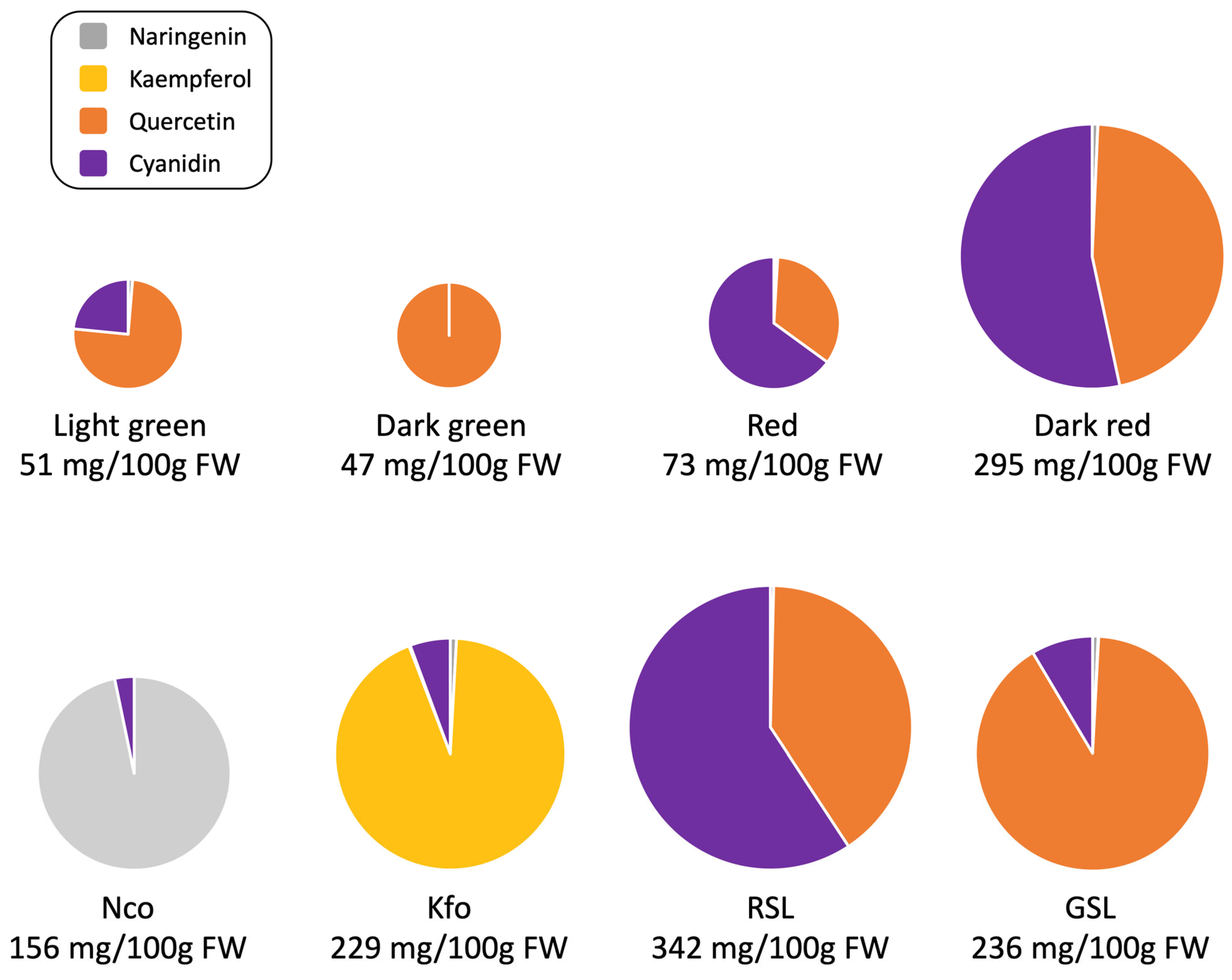
2.6. Flavonoid-Overproducing Lines Accumulate Higher Amounts of Flavonoids Compared to Parental Lines and Controls
3. Discussion
4. Materials and Methods
4.1. Plant Material
4.2. Field Cultivation
4.3. Disease Resistance Evaluation
4.3.1. Resistance to Downy Mildew
4.3.2. Resistance to Impatiens Necrotic Spot Virus
4.3.3. Resistance to Lettuce Drop
4.4. Quantification of Total Anthocyanins and Chlorophyll in the Field
4.5. Evaluations of Postharvest Quality
4.5.1. Tissue Deterioration
4.5.2. Enzymatic Discoloration
4.6. Determination of Leaf Elemental Composition
4.7. Flavonoid Quantification
4.7.1. Extraction and Acid Hydrolysis of Flavonoid Aglycones
4.7.2. UPLC-MS/MS Analysis
4.7.3. Flavonoid Aglycone Quantification
4.8. Statistical Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- World Health Organization. Cardiovascular Diseases (CVDs). Available online: https://www.who.int/news-room/fact-sheets/detail/cardiovascular-diseases-(cvds) (accessed on 7 June 2023).
- Kim, Y.; Je, Y. Flavonoid intake and mortality from cardiovascular disease and all causes: A meta-analysis of prospective cohort studies. Clin. Nutr. ESPEN 2017, 20, 68–77. [Google Scholar] [CrossRef] [PubMed]
- Zhan, J.; Liu, Y.-J.; Cai, L.-B.; Xu, F.-R.; Xie, T.; He, Q.-Q. Fruit and vegetable consumption and risk of cardiovascular disease: A meta-analysis of prospective cohort studies. Crit. Rev. Food Sci. Nutr. 2017, 57, 1650–1663. [Google Scholar] [CrossRef] [PubMed]
- Reygaert, W.C. An update on the health benefits of green tea. Beverages 2017, 3, 6. [Google Scholar] [CrossRef]
- Basu, A.; Betts, N.M.; Mulugeta, A.; Tong, C.; Newman, E.; Lyons, T.J. Green tea supplementation increases glutathione and plasma antioxidant capacity in adults with the metabolic syndrome. Nutr. Res. 2013, 33, 180–187. [Google Scholar] [CrossRef]
- Jan, A.T.; Kamli, M.R.; Murtaza, I.; Singh, J.B.; Ali, A.; Haq, Q.M.R. Dietary Flavonoid Quercetin and Associated Health Benefits—An Overview. Food Rev. Int. 2010, 26, 302–317. [Google Scholar] [CrossRef]
- Hertog, M.G.L.; Feskens, E.J.M.; Kromhout, D.; Hollman, P.C.H.; Katan, M.B. Dietary antioxidant flavonoids and risk of coronary heart disease: The Zutphen Elderly Study. Lancet 1993, 342, 1007–1011. [Google Scholar] [CrossRef]
- Weber, C.; Davis, W.V.; Lucier, G.; Wechsler, S. Lettuce Trends: Conventional, Organic Growth, and Production. In Vegetables and Pulses Outlook; VGS-370; Davis, W.V., Weber, C., Wechsler, S., Lucier, G., Soria Rodriguez, M., Yeh, A., Fan, X., Eds.; United States Department of Agriculture: Washington, DC, USA, 2023. [Google Scholar]
- Simko, I.; Hayes, R.J.; Mou, B.; McCreight, J.D. Lettuce and Spinach. In Yield Gains in Major U.S. Field Crops; Smith, S., Diers, B., Specht, J., Carver, B., Eds.; American Society of Agronomy, Inc., Crop Science Society of America, Inc., and Soil Science Society of America, Inc.: Madison, WI, USA, 2014; Chapter 4; pp. 53–85. [Google Scholar] [CrossRef]
- Hayes, R.J.; Simko, I. Breeding lettuce for improved fresh-cut processing. Acta Hortic. 2016, 1141, 65–76. [Google Scholar] [CrossRef]
- Hayes, R.J.; Sandoya, G.; Mou, B.; Simko, I.; Subbarao, K.V. Release of three iceberg lettuce populations with combined resistance to two soilborne diseases. HortScience 2018, 53, 247–250. [Google Scholar] [CrossRef]
- Damerum, A.; Chapman, M.A.; Taylor, G. Innovative breeding technologies in lettuce for improved post-harvest quality. Postharvest Biol. Technol. 2020, 168, 111266. [Google Scholar] [CrossRef]
- Armas Gutierrez, I. Nutritional Enhancement of Lettuce Using Mutational Breeding. Master’s Thesis, Rutgers University, New Brunswick, NJ, USA, 2015. [Google Scholar]
- Damerum, A.; Selmes, S.L.; Biggi, G.F.; Clarkson, G.J.; Rothwell, S.D.; Truco, M.J.; Michelmore, R.W.; Hancock, R.D.; Shellcock, C.; Chapman, M.A.; et al. Elucidating the genetic basis of antioxidant status in lettuce (Lactuca sativa). Hortic. Res. 2015, 2, 15055. [Google Scholar] [CrossRef]
- Cheng, D.M.; Pogrebnyak, N.; Kuhn, P.; Krueger, C.G.; Johnson, W.D.; Raskin, I. Development and phytochemical characterization of high polyphenol red lettuce with anti-diabetic properties. PLoS ONE 2014, 9, e91571. [Google Scholar] [CrossRef]
- Gurdon, C.; Poulev, A.; Armas, I.; Satorov, S.; Tsai, M.; Raskin, I. Genetic and Phytochemical Characterization of Lettuce Flavonoid Biosynthesis Mutants. Sci. Rep. 2019, 9, 3305. [Google Scholar] [CrossRef]
- Gurdon, C.; Kozik, A.; Tao, R.; Poulev, A.; Armas, I.; Michelmore, R.W.; Raskin, I. Isolating an active and inactive CACTA transposon from lettuce color mutants and characterizing their family. Plant Physiol. 2021, 186, 929–944. [Google Scholar] [CrossRef] [PubMed]
- Kim, M.J.; Moon, Y.; Tou, J.C.; Mou, B.; Waterland, N.L. Nutritional value, bioactive compounds and health benefits of lettuce (Lactuca sativa L.). J. Food Compos. Anal. 2016, 49, 19–34. [Google Scholar] [CrossRef]
- Simko, I. Genetic variation and relationship among content of vitamins, pigments, and sugars in baby leaf lettuce. Food Sci. Nutr. 2019, 7, 3317–3326. [Google Scholar] [CrossRef] [PubMed]
- Ferreres, F.; Gil, M.I.; Castañer, M.; Tomás-Barberán, F.A. Phenolic Metabolites in Red Pigmented Lettuce (Lactuca sativa). Changes with Minimal Processing and Cold Storage. J. Agric. Food Chem. 1997, 45, 4249–4254. [Google Scholar] [CrossRef]
- Llorach, R.; Martínez-Sánchez, A.; Tomás-Barberán, F.A.; Gil, M.I.; Ferreres, F. Characterisation of polyphenols and antioxidant properties of five lettuce varieties and escarole. Food Chem. 2008, 108, 1028–1038. [Google Scholar] [CrossRef] [PubMed]
- Medina-Lozano, I.; Bertolín, J.R.; Díaz, A. Nutritional value of commercial and traditional lettuce (Lactuca sativa L.) and wild relatives: Vitamin C and anthocyanin content. Food Chem. 2021, 359, 129864. [Google Scholar] [CrossRef]
- Cheng, D.M.; Pogrebnyak, N.; Kuhn, P.; Poulev, A.; Waterman, C.; Rojas-Silva, P.; Johnson, W.D.; Raskin, I. Polyphenol-rich Rutgers Scarlet Lettuce improves glucose metabolism and liver lipid accumulation in diet-induced obese C57BL/6 mice. Nutrition 2014, 30, S52–S58. [Google Scholar] [CrossRef] [PubMed]
- Cheng, D.M.; Roopchand, D.E.; Poulev, A.; Kuhn, P.; Armas, I.; Johnson, W.D.; Oren, A.; Ribnicky, D.; Zelzion, E.; Bhattacharya, D.; et al. High phenolics Rutgers Scarlet Lettuce improves glucose metabolism in high fat diet-induced obese mice. Mol. Nutr. Food Res. 2016, 60, 2367–2378. [Google Scholar] [CrossRef]
- Calderón-Montaño, J.M.; Burgos-Morón, E.; Pérez-Guerrero, C.; López-Lázaro, M. A review on the dietary flavonoid kaempferol. Mini. Rev. Med. Chem. 2011, 11, 298–344. [Google Scholar] [CrossRef]
- Hertog, M.G.L.; Hollman, P.C.H.; Katan, M.B. Content of potentially anticarcinogenic flavonoids of 28 vegetables and 9 fruits commonly consumed in the Netherlands. J. Agric. Food Chem. 1992, 40, 2379–2383. [Google Scholar] [CrossRef]
- Justesen, U.; Knuthsen, P.; Leth, T. Quantitative analysis of flavonols, flavones, and flavanones in fruits, vegetables and beverages by high-performance liquid chromatography with photo-diode array and mass spectrometric detection. J. Chromatogr. A 1998, 799, 101–110. [Google Scholar] [CrossRef]
- Inocencio, C.; Rivera, D.; Alcaraz, F.; Tomás-Barberán, F.A. Flavonoid content of commercial capers (Capparis spinosa, C. sicula and C. orientalis) produced in mediterranean countries. Eur. Food Res. Technol. 2000, 212, 70–74. [Google Scholar] [CrossRef]
- Franke, A.A.; Custer, L.J.; Arakaki, C.; Murphy, S.P. Vitamin C and flavonoid levels of fruits and vegetables consumed in Hawaii. J. Food Compos. Anal. 2004, 17, 1–35. [Google Scholar] [CrossRef]
- Yang, R.Y.; Lin, S.; Kuo, G. Content and distribution of flavonoids among 91 edible plant species. Asia Pac. J. Clin. Nutr. 2008, 17 (Suppl. S1), 275–279. [Google Scholar]
- Horiba, T.; Nishimura, I.; Nakai, Y.; Abe, K.; Sato, R. Naringenin chalcone improves adipocyte functions by enhancing adiponectin production. Mol. Cell Endocrinol. 2010, 323, 208–214. [Google Scholar] [CrossRef] [PubMed]
- Iwamura, C.; Shinoda, K.; Yoshimura, M.; Watanabe, Y.; Obata, A.; Nakayama, T. Naringenin Chalcone Suppresses Allergic Asthma by Inhibiting the Type-2 Function of CD4 T Cells. Allergol. Int. 2010, 59, 67–73. [Google Scholar] [CrossRef] [PubMed]
- Yamamoto, T.; Yoshimura, M.; Yamaguchi, F.; Kouchi, T.; Tsuji, R.; Saito, M.; Obata, A.; Kikuchi, M. Anti-allergic Activity of Naringenin Chalcone from a Tomato Skin Extract. Biosci. Biotechnol. Biochem. 2004, 68, 1706–1711. [Google Scholar] [CrossRef] [PubMed]
- Muir, S.R.; Collins, G.J.; Robinson, S.; Hughes, S.; Bovy, A.; Ric De Vos, C.H.; van Tunen, A.J.; Verhoeyen, M.E. Overexpression of petunia chalcone isomerase in tomato results in fruit containing increased levels of flavonols. Nat. Biotechnol. 2001, 19, 470–474. [Google Scholar] [CrossRef] [PubMed]
- Saito, K.; Yonekura-Sakakibara, K.; Nakabayashi, R.; Higashi, Y.; Yamazaki, M.; Tohge, T.; Fernie, A.R. The flavonoid biosynthetic pathway in Arabidopsis: Structural and genetic diversity. Plant Physiol. Biochem. 2013, 72, 21–34. [Google Scholar] [CrossRef]
- Li, Q.; Kubota, C. Effects of supplemental light quality on growth and phytochemicals of baby leaf lettuce. Environ. Exp. Bot. 2009, 67, 59–64. [Google Scholar] [CrossRef]
- Goto, E.; Hayashi, K.; Furuyama, S.; Hikosaka, S.; Ishigami, Y. Effect of UV light on phytochemical accumulation and expression of anthocyanin biosynthesis genes in red leaf lettuce. Acta Hortic. 2016, 1134, 179–186. [Google Scholar] [CrossRef]
- Kitazaki, K.; Fukushima, A.; Nakabayashi, R.; Okazaki, Y.; Kobayashi, M.; Mori, T.; Nishizawa, T.; Reyes-Chin-Wo, S.; Michelmore, R.W.; Saito, K.; et al. Metabolic Reprogramming in Leaf Lettuce Grown Under Different Light Quality and Intensity Conditions Using Narrow-Band LEDs. Sci. Rep. 2018, 8, 7914. [Google Scholar] [CrossRef] [PubMed]
- Meng, Q.; Boldt, J.; Runkle, E.S. Blue Radiation Interacts with Green Radiation to Influence Growth and Predominantly Controls Quality Attributes of Lettuce. J. Amer. Soc. Hort. Sci. 2020, 145, 75–87. [Google Scholar] [CrossRef]
- Meng, Q.; Runkle, E.S. Growth Responses of Red-Leaf Lettuce to Temporal Spectral Changes. Front. Plant Sci. 2020, 11, 571788. [Google Scholar] [CrossRef] [PubMed]
- Sytar, O.; Zivcak, M.; Bruckova, K.; Brestic, M.; Hemmerich, I.; Rauh, C.; Simko, I. Shift in accumulation of flavonoids and phenolic acids in lettuce attributable to changes in ultraviolet radiation and temperature. Sci. Hortic. 2018, 239, 193–204. [Google Scholar] [CrossRef]
- Ryan, K.G.; Swinny, E.E.; Winefield, C.; Markham, K.R. Flavonoids and UV Photoprotection in Arabidopsis Mutants. Z. aturforsch C. J. Biosci. 2001, 56, 745–754. [Google Scholar] [CrossRef]
- Havaux, M.; Kloppstech, K. The protective functions of carotenoid and flavonoid pigments against excess visible radiation at chilling temperature investigated in Arabidopsis npq and tt mutants. Planta 2001, 213, 953–966. [Google Scholar] [CrossRef]
- Landry, L.G.; Chapple, C.C.; Last, R.L. Arabidopsis mutants lacking phenolic sunscreens exhibit enhanced ultraviolet-B injury and oxidative damage. Plant Physiol. 1995, 109, 1159–1166. [Google Scholar] [CrossRef] [PubMed]
- Gazula, A.; Kleinhenz, M.D.; Scheerens, J.C.; Ling, P.P. Anthocyanin levels in nine lettuce (Lactuca sativa) cultivars: Influence of planting date and relations among analytic, instrumented, and visual assessments of color. HortScience 2007, 42, 232–238. [Google Scholar] [CrossRef]
- Marin, A.; Ferreres, F.; Barberá, G.G.; Gil, M.I. Weather variability influences color and phenolic content of pigmented baby leaf lettuces throughout the season. J. Agric. Food Chem. 2015, 63, 1673–1681. [Google Scholar] [CrossRef] [PubMed]
- Cao, J.; Chen, W.; Zhang, Y.; Zhang, Y.; Zhao, X. Content of Selected Flavonoids in 100 Edible Vegetables and Fruits. Food Sci. Technol. Res. 2010, 16, 395–402. [Google Scholar] [CrossRef]
- Arabbi, P.R.; Genovese, M.I.; Lajolo, F.M. Flavonoids in Vegetable Foods Commonly Consumed in Brazil and Estimated Ingestion by the Brazilian Population. J. Agric. Food Chem. 2004, 52, 1124–1131. [Google Scholar] [CrossRef]
- Jaakola, L.; Hohtola, A. Effect of latitude on flavonoid biosynthesis in plants. Plant Cell Environ. 2010, 33, 1239–1247. [Google Scholar] [CrossRef] [PubMed]
- Zahra, N.; Hafeez, M.B.; Shaukat, K.; Wahid, A.; Hasanuzzaman, M. Fe toxicity in plants: Impacts and remediation. Physiol. Plant. 2021, 173, 201–222. [Google Scholar] [CrossRef]
- Sthapit Kandel, J.; Peng, H.; Hayes, R.J.; Mou, B.; Simko, I. Genome-wide association mapping reveals loci for shelf life and developmental rate of lettuce. Theor. Appl. Genet. 2020, 133, 1947–1966. [Google Scholar] [CrossRef]
- Rosental, L.; Still, D.W.; You, Y.; Hayes, R.J.; Simko, I. Mapping and identification of genetic loci affecting earliness of bolting and flowering in lettuce. Theor. Appl. Genet. 2021, 134, 3319–3337. [Google Scholar] [CrossRef]
- Padmavati, M.; Reddy, A.R. Flavonoid Biosynthetic Pathway and Cereal Defence Response: An Emerging Trend in Crop Biotechnology. J. Plant Biochem. Biotechnol. 1999, 8, 15–20. [Google Scholar] [CrossRef]
- Ramaroson, M.-L.; Koutouan, C.; Helesbeux, J.-J.; Le Clerc, V.; Hamama, L.; Geoffriau, E.; Briard, M. Role of Phenylpropanoids and Flavonoids in Plant Resistance to Pests and Diseases. Molecules 2022, 27, 8371. [Google Scholar] [CrossRef]
- Simko, I.; Hayes, R.J.; Kramer, M. Computing Integrated Ratings from Heterogeneous Phenotypic Assessments: A Case Study of Lettuce Postharvest Quality and Downy Mildew Resistance. Crop Sci. 2012, 52, 2131–2142. [Google Scholar] [CrossRef]
- Simko, I.; Peng, H.; Sthapit Kandel, J.; Zhao, R. Genome-wide association mapping reveals genomic regions frequently associated with lettuce field resistance to downy mildew. Theor. Appl. Genet. 2022, 135, 2009–2024. [Google Scholar] [CrossRef] [PubMed]
- Simko, I.; Hayes, R.J.; Bull, C.T.; Mou, B.; Luo, Y.; Trent, M.A.; Atallah, A.J.; Ryder, E.J.; Sideman, R.G. Characterization and Performance of 16 New Inbred Lines of Lettuce. HortScience 2014, 49, 679–687. [Google Scholar] [CrossRef]
- Hayes, R.J.; Wu, B.; Pryor, B.; Chitrampalam, P.; Subbarao, K. Assessment of Resistance in Lettuce (Lactuca sativa L.) to Mycelial and Ascospore Infection by Sclerotinia minor Jagger and S. sclerotiorum (Lib.) de Bary. HortScience 2010, 45, 333–341. [Google Scholar] [CrossRef]
- Simko, I.; Sthapit Kandel, J.; Peng, H.; Zhao, R.; Subbarao, K.V. Genetic determinants of lettuce resistance to drop caused by Sclerotinia minor identified through genome-wide association mapping frequently co-locate with loci regulating anthocyanin content. Theor. Appl. Genet. 2023, 136, 180. [Google Scholar] [CrossRef] [PubMed]
- Mamo, B.E.; Hayes, R.J.; Truco, M.J.; Puri, K.D.; Michelmore, R.W.; Subbarao, K.V.; Simko, I. The genetics of resistance to lettuce drop (Sclerotinia spp.) in lettuce in a recombinant inbred line population from Reine des Glaces × Eruption. Theor. Appl. Genet. 2019, 132, 2439–2460. [Google Scholar] [CrossRef]
- Hasegawa, D.K.; Del Pozo-Valdivia, A.I. Epidemiology and Economic Impact of Impatiens Necrotic Spot Virus: A Resurging Pathogen Affecting Lettuce in the Salinas Valley of California. Plant Dis. 2022, 107, 1192–1201. [Google Scholar] [CrossRef] [PubMed]
- Simko, I.; Richardson, C.E.; Wintermantel, W.M. Variation within Lactuca spp. for Resistance to Impatiens necrotic spot virus. Plant Dis. 2017, 102, 341–348. [Google Scholar] [CrossRef] [PubMed]
- Simko, I.; Hasegawa, D.K.; Peng, H.; Zhao, R. Genetic and physiological determinants of lettuce partial resistance to Impatiens necrotic spot virus. Front. Plant Sci. 2023, 14, 1163683. [Google Scholar] [CrossRef]
- Peng, H.; Simko, I. Extending lettuce shelf life through integrated technologies. Curr. Opin. Biotechnol. 2023, 81, 102951. [Google Scholar] [CrossRef] [PubMed]
- Van Dun, C.M.P. Screening Method for Selecting Plants that Show Reduced Wound-Induced Surface Discolouration and Plant and Plant Parts thus Obtained. Patent US 8809631B2, 19 August 2014. [Google Scholar]
- Simko, I.; Atallah, A.J.; Ochoa, O.E.; Antonise, R.; Galeano, C.H.; Truco, M.J.; Michelmore, R.W. Identification of QTLs conferring resistance to downy mildew in legacy cultivars of lettuce. Sci. Rep. 2013, 3, 2875. [Google Scholar] [CrossRef] [PubMed]
- Subbarao, K.V. Progress toward integrated management of lettuce drop. Plant Dis. 1998, 82, 1068–1078. [Google Scholar] [CrossRef] [PubMed]
- Parry, C.; Blonquist, J.M., Jr.; Bugbee, B. In situ measurement of leaf chlorophyll concentration: Analysis of the optical/absolute relationship. Plan Cell Environ. 2014, 37, 2508–2520. [Google Scholar] [CrossRef]
- van den Berg, A.K.; Perkins, T.D. Nondestructive estimation of anthocyanin content in autumn sugar maple leaves. HortScience 2005, 40, 685–686. [Google Scholar] [CrossRef]
- Simko, I. Predictive modeling of a leaf conceptual midpoint quasi-color (CMQ) using an artificial neural network. Sensors 2020, 20, 3938. [Google Scholar] [CrossRef]
- Hayes, R.J.; Galeano, C.H.; Luo, Y.; Antonise, R.; Simko, I. Inheritance of decay of fresh-cut lettuce in a recombinant inbred line population from ‘Salinas 88’ × ‘La Brillante’. J. Am. Soc. Hortic. Sci. 2014, 139, 388–398. [Google Scholar] [CrossRef]
- Simko, I.; Hayes, R.J. Accuracy, reliability, and timing of visual evaluations of decay in fresh-cut lettuce. PLoS ONE 2018, 13, e0194635. [Google Scholar] [CrossRef]
- Rueden, C.T.; Schindelin, J.; Hiner, M.C.; DeZonia, B.E.; Walter, A.E.; Arena, E.T.; Eliceiri, K.W. ImageJ2: ImageJ for the next generation of scientific image data. BMC Bioinform. 2017, 18, 529. [Google Scholar] [CrossRef]
- Rossini Oliva, S.; Raitio, H.; Mingorance, M.D. Comparison of two wet digestion procedures for multi-element analysis of plant samples. Commun. Soil Sci. Plant Anal. 2003, 34, 2913–2923. [Google Scholar] [CrossRef]
- United States Environmental Protection Agency (EPA). Method 6010B. Inductively Coupled Plasma-Atomic Emission Spectrometry. Available online: https://www.epa.gov/sites/default/files/documents/6010b.pdf (accessed on 14 November 2022).
- Soltanpour, P.N.; Johnson, G.W.; Workman, S.M.; Jones, J.B., Jr.; Miller, R.O. Inductively coupled plasma emission spectrometry and inductively coupled plasma-mass spectrometry. In Methods of Soil Analysis: Part 3 Chemical Methods; Sparks, D.L., Page, A.L., Helmke, P.A., Loeppert, R.H., Soltanpour, P.N., Tabatabai, M.A., Johnston, C.T., Sumner, M.E., Eds.; Soil Science Society of America, Inc.: Madison, WI, USA, 1996; Chapter 5; pp. 91–139. [Google Scholar] [CrossRef]
- Bremner, J.M. Nitrogen-total. In Methods of Soil Analysis: Part 3 Chemical Methods; Sparks, D.L., Page, A.L., Helmke, P.A., Loeppert, R.H., Soltanpour, P.N., Tabatabai, M.A., Johnston, C.T., Sumner, M.E., Eds.; Soil Science Society of America, Inc.: Madison, WI, USA, 1996; Chapter 37; pp. 1085–1121. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Accession | Category | Origin | Color | Horticultural Type |
|---|---|---|---|---|
| RSL-NAR | Mutant line | Tissue culture derivative of Annapolis | Dark red | Romaine like |
| RSL-NBR | Mutant line | Tissue culture derivative of Grand Rapids Blackhawk | Dark red | Loose-leaf |
| RSL-NFR | Mutant line | Tissue culture derivative of Firecracker | Dark red | Loose-leaf |
| GSL | Mutant line | Natural mutant of RSL-NAR | Green with red patches | Romaine like |
| GSL-DG | Mutant line | Natural mutant of GSL | Dark green | Romaine like |
| KfoA | Mutant line | EMS induced mutant of Firecracker | Green with red tingle | Loose-leaf |
| KfoB | Mutant line | EMS induced mutant of Firecracker | Green with red tingle | Loose-leaf |
| Nco | Mutant line | EMS induced mutant of Firecracker | Yellow-green | Loose-leaf |
| Annapolis | Parental line | Released cultivar | Dark red | Romaine |
| Firecracker | Parental line | Released cultivar | Dark red | Loose-leaf |
| Darkland | Control | Released cultivar | Dark green | Romaine |
| Eruption | Control | Released cultivar | Red (mostly) | Latin |
| Grand Rapids | Control | Released cultivar | Light green | Loose-leaf |
| Merlot | Control | Released cultivar | Dark red | Loose-leaf |
| SM13-L2 | Control | Grand Rapids × Iceberg | Light green | Loose-leaf |
| Accession | Plant Weight (g) | Plant Height (cm) | Plant Width (cm) | Chlorophyll (SPAD-sqrt) | Anthocyanins (ACI-lb) | Downy Mildew (Rating) | Lettuce drop (Incidence %) | INSV (Incidence %) | Tissue Deterioration (AUDPS) | Enzymatic Discoloration (Area mm2) |
|---|---|---|---|---|---|---|---|---|---|---|
| RSL-NAR | 353 c | 29 bc | 23 bc | 6.5 a | 8.0 a | 0.04 | 52 a–c | 10 ab | 29 ab | - |
| RSL-NBR | 433 c | 55 a | 25 b | 6.4 a | 7.5 a | 0.36 | 55 a–c | 3 ab | 25 ab | 19 de |
| RSL-NFR | 434 c | 31 b | 24 bc | 6.4 a | 7.8 a | 1.18 | 61 ab | 11 ab | 32 ab | 61 a–c |
| GSL | 571 bc | 26 bc | 23 bc | 7.0 a | 3.4 d–f | 0.35 | 64 ab | 13 ab | 26 ab | 22 c–e |
| GSL-DG | 510 bc | 29 bc | 20 c | 6.7 a | 4.0 de | 1.01 | 55 a–c | 11 ab | 30 ab | 63 ab |
| KfoA | 411 c | 27 bc | 27 b | 6.1 ab | 3.3 d–f | 0.03 | 71 a | 24 a | 35 a | 94 a |
| KfoB | 500 c | 25 cd | 24 bc | 6.1 ab | 4.7 cd | 0.73 | 72 a | 10 ab | 32 a | 93 a |
| Nco | 10 d | 10 e | 6 d | 5.2 ab | 1.3 e–g | - | 47 a–c | 3 ab | - | - |
| Annapolis | 433 c | 30 b | 23 bc | 6.7 a | 7.9 a | 0.64 | 33 bc | 5 ab | 34 a | 67 ab |
| Darkland | 965 a | 29 bc | 21 bc | 7.2 a | 3.4 d–f | 0.79 | 65 ab | 21 ab | 17 b | 49 b–d |
| Eruption | 495 c | 20 d | 23 bc | 6.4 a | 5.7 bc | 0.85 | 19 c | 4 b | 22 ab | 95 a |
| Firecracker | 389 c | 31 b | 23 bc | 6.4 a | 7.1 ab | 0.40 | 68 ab | 11 ab | 25 ab | 75 ab |
| Grand Rapids | 447 bc | 29 bc | 31 ab | 5.9 ab | 1.9 fg | 0.03 | 72 a–c | 12 ab | 25 ab | 46 b–e |
| Merlot | 435 c | 30 bc | 26 b | 6.2 ab | 7.7 a | 0.10 | 62 a–c | 15 ab | 28 ab | 92 a |
| SM13-L2 | 764 ab | 28 bc | 36 a | 5.0 b | 1.8 g | 0.10 | 54 ab | 8 ab | 17 b | 7 e |
| Accession | N | P | K | Ca | S | Mg | Fe | B | Cl- | Mn | Zn | Cu |
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| RSL-NAR | 3.40 b–d | 6155 a–c | 45,984 b–d | 11,236 ab | 2296 bc | 4975 bc | 225 b | 30.3 a | 30,629 a–d | 157 a | 31 ab | 5.5 b |
| RSL-NBR | 4.04 ab | 3533 e | 37,882 cd | 11,158 ab | 2505 ab | 7680 a | 300 b | 26.6 ab | 38,655 a | 107 a–c | 32 ab | 7.2 b |
| RSL-NFR | 4.13 ab | 5658 a–d | 49,574 a–c | 8960 ab | 2294 bc | 4737 bc | 210 b | 28.9 ab | 38,032 ab | 107 a–c | 32 ab | 7.6 b |
| GSL | 3.72 ab | 5079 b–e | 31,848 d | 8547 ab | 2246 bc | 3595 bc | 242 b | 28.0 ab | 14,483 g | 103 a–c | 31 ab | 4.9 b |
| GSL-DG | 3.62 a–c | 5389 a–e | 35,000 cd | 9933 ab | 2252 bc | 4432 bc | 251 b | 24.8 ab | 22,591 c–g | 96 a–c | 29 ab | 6.6 b |
| KfoA | 3.75ab | 4871 b–e | 48,708 a–c | 11,284 ab | 2461 a–c | 5568 ab | 444 b | 29.7 a | 29,705 a–e | 146 a | 32 ab | 6.9 b |
| KfoB | 3.41 a–d | 5138 b–e | 43,713 b–d | 10,310 ab | 2306 bc | 5351 a–c | 247 b | 28.7 ab | 29,143 a–e | 127 a–c | 33 ab | 6.5 b |
| Nco | 4.22 ab | 6683 ab | 62,969 a | 13,970 a | 2809 a | 4912 bc | 1523 a | 27.5 ab | 32,065 a–c | 143 ab | 40 ab | 5.3 b |
| Annapolis | 4.05 ab | 5432 a–e | 44,883 b–d | 10,575 ab | 2305 bc | 4080 bc | 357 b | 27.7 ab | 18,070 fg | 87 a–c | 31 ab | 8.0 ab |
| Darkland | 2.79 cd | 4107 de | 31,240 d | 7139 b | 1764 d | 2887 c | 217 b | 20.7 b | 18,428 fg | 88 a–c | 23 b | 3.8 b |
| Eruption | 3.89 ab | 4724 c–e | 55,544 ab | 9693 ab | 2112 b–d | 3778 bc | 402 b | 27.4 ab | 16,679 g | 101 a–c | 43 a | 12.4 a |
| Firecracker | 4.43 a | 7266 a | 50,426 a–c | 7849 b | 2320 bc | 3493 bc | 216 b | 29.9 a | 20,199 d–g | 70 bc | 41 a | 6.9 b |
| Grand Rapids | 3.76 ab | 5139 b–e | 45,783 b–d | 9212 ab | 2318 bc | 3377 bc | 253 b | 24.9 ab | 25,025 c–g | 95 a–c | 34 ab | 6.0 b |
| Merlot | 3.72 ab | 6375 a–c | 43,438 b–d | 7627 b | 2063 cd | 3253 bc | 267 b | 26.7 ab | 19,831 e–g | 67 c | 38 ab | 7.7 b |
| SM13-L2 | 2.52 d | 4121 de | 33,019 d | 9119 ab | 2075 b–d | 3740 bc | 181 b | 24.5 ab | 27,671 b–f | 97 a–c | 28 ab | 5.5 b |
| Accession | Mo | Ni | Na | Si | Al | Ba | Cd | Cr | Li | Sr | Ti | |
| RSL-NAR | 0.48 b | 1.13 b–d | 6072 bc | 451 bc | 175 b | 6.4 bc | 0.75 a–c | 0.48 b | 1.40 b | 58 ab | 11 b | |
| RSL-NBR | 0.70 b | 0.85 cd | 12,216 a | 438 bc | 271 b | 5.2 bc | 0.77 a–c | 0.65 b | 1.41 b | 53 b | 17 b | |
| RSL-NFR | 0.49 b | 1.06 b–d | 6212 bc | 475 bc | 161 b | 6.6 bc | 0.57 bc | 0.48 b | 1.22 b | 48 b | 9 b | |
| GSL | 0.45 b | 0.84 cd | 4089 bc | 418 bc | 225 b | 4.4 c | 0.57 bc | 0.46 b | 0.94 b | 43 b | 13 b | |
| GSL-DG | 0.63 b | 0.89 b–d | 6046 bc | 472 bc | 226 b | 5.9 bc | 0.65 a–c | 0.51 b | 0.96 b | 52 b | 13 b | |
| KfoA | 0.98 b | 0.93 b–d | 6168 bc | 727 b | 396 b | 5.9 bc | 0.96 a–c | 0.76 b | 1.67 b | 55 ab | 27 b | |
| KfoB | 0.43 b | 0.81 cd | 6403 bc | 573 bc | 210 b | 4.9 bc | 1.11 ab | 0.53 b | 1.26 b | 50 b | 13 b | |
| Nco | 4.04 a | 2.18 a | 6124 bc | 1320 a | 1450 a | 16.9 a | 1.15 a | 2.27 a | 5.43 a | 84 a | 99 a | |
| Annapolis | 0.99 b | 1.07 b–d | 5293 bc | 588 bc | 328 b | 8.2 b | 0.88 a–c | 0.68 b | 1.29 b | 57 ab | 20 b | |
| Darkland | 0.78 b | 0.67 d | 2768 c | 438 bc | 228 b | 4.5 c | 0.44 c | 0.47 b | 0.71 b | 30 b | 13 b | |
| Eruption | 1.06 b | 0.93 b–d | 6072 bc | 543 bc | 390 b | 6.8 bc | 1.17 a | 0.75 b | 1.36 b | 52 b | 24 b | |
| Firecracker | 0.54 b | 1.65 ab | 3966 bc | 450 bc | 180 b | 5.1 bc | 0.51 c | 0.51 b | 0.83 b | 39 b | 9 b | |
| Grand Rapids | 0.51 b | 0.86 b–d | 3537 c | 318 c | 249 b | 4.8 bc | 0.57 bc | 0.53 b | 0.86 b | 44 b | 14 b | |
| Merlot | 0.60 b | 1.48 a–c | 4959 bc | 392 bc | 246 b | 5.2 bc | 0.51 c | 0.57 b | 0.78 b | 37 b | 14 b | |
| SM13-L2 | 0.24 b | 0.55 d | 7904 b | 289 c | 160 b | 4.6 c | 0.70 a–c | 0.37 b | 1.01 b | 45 b | 9 b | |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Cho, E.; Gurdon, C.; Zhao, R.; Peng, H.; Poulev, A.; Raskin, I.; Simko, I. Phytochemical and Agronomic Characterization of High-Flavonoid Lettuce Lines Grown under Field Conditions. Plants 2023, 12, 3467. https://0-doi-org.brum.beds.ac.uk/10.3390/plants12193467
Cho E, Gurdon C, Zhao R, Peng H, Poulev A, Raskin I, Simko I. Phytochemical and Agronomic Characterization of High-Flavonoid Lettuce Lines Grown under Field Conditions. Plants. 2023; 12(19):3467. https://0-doi-org.brum.beds.ac.uk/10.3390/plants12193467
Chicago/Turabian StyleCho, Eunjin, Csanad Gurdon, Rebecca Zhao, Hui Peng, Alexander Poulev, Ilya Raskin, and Ivan Simko. 2023. "Phytochemical and Agronomic Characterization of High-Flavonoid Lettuce Lines Grown under Field Conditions" Plants 12, no. 19: 3467. https://0-doi-org.brum.beds.ac.uk/10.3390/plants12193467