
Vernalization Procedure of Tuberous Roots Affects Growth, Photosynthesis and Metabolic Profile of Ranunculus asiaticus L.

 , , , and
, , , and
Abstract
:1. Introduction
2. Results
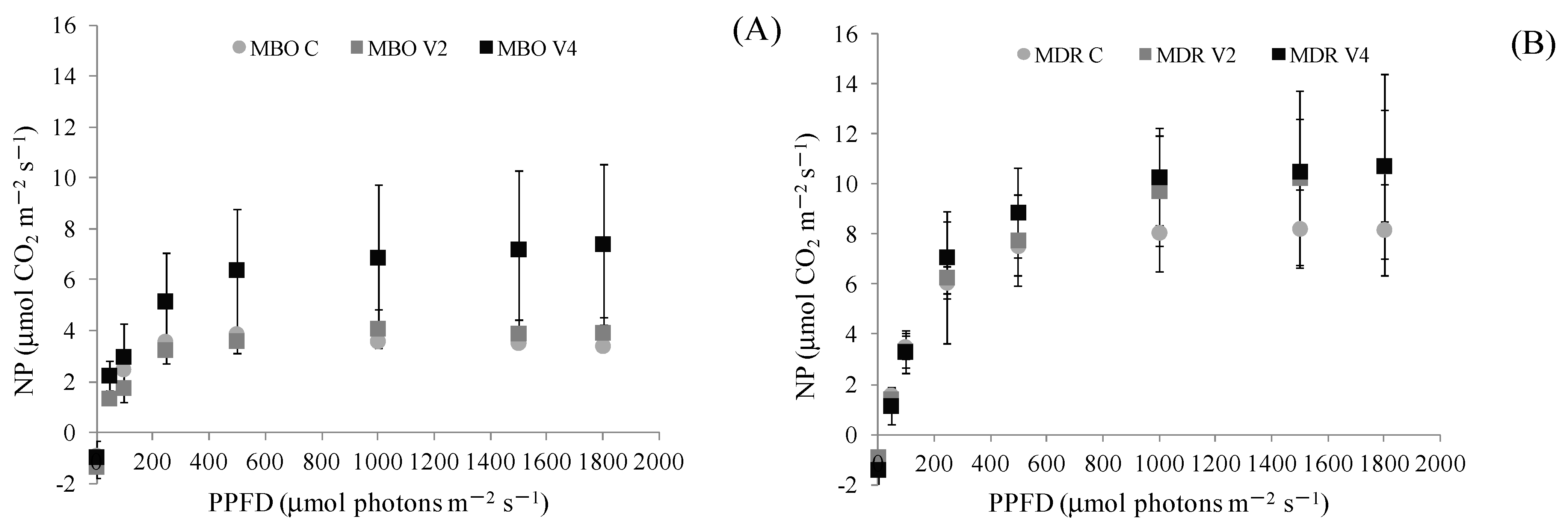
2.1. Net Photosynthesis, Chlorophyll a Fluorescence and Leaf Photosynthetic Pigments
2.2. Plant Growth and Flowering
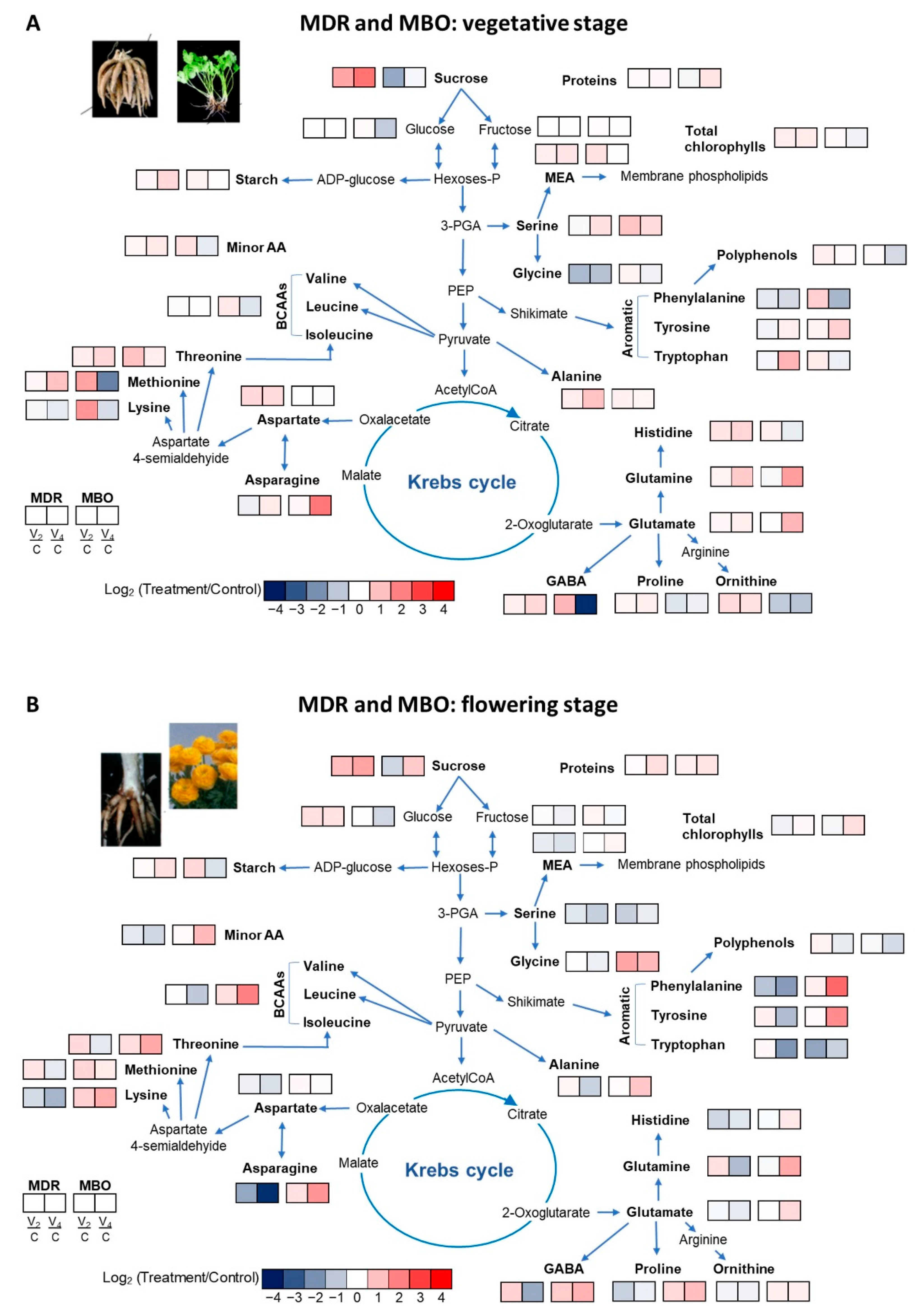
2.3. Leaves’ Metabolites Profiling
3. Discussion
4. Materials and Methods
- -
- only rehydration: exposure to 12 °C for 24 h in humid chamber (Control, C);
- -
- rehydration followed by vernalization at 3.5 °C for 2 weeks (V2);
- -
- rehydration followed by vernalization at 3.5 °C for 4 weeks (V4).
4.1. Net Photosynthesis, Chlorophyll a Fluorescence and Leaf Photosynthetic Pigments
4.2. Plant Growth and Flowering
4.3. Metabolic Profile
4.3.1. Starch and Soluble Carbohydrate Analysis
4.3.2. Soluble Proteins, Free Amino Acid Analysis
4.3.3. Polyphenols Analysis
4.3.4. Chlorophylls and Carotenoids Analysis
5. Statistical Analysis
6. Conclusions
Supplementary Materials
Author Contributions
Funding
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Abbreviations
References
- Beruto, M.; Rabaglio, M.; Viglione, S.; Van Labeke, M.-C.; Dhooghe, E. Ranunculus. In Ornamental Crops; Van Huylenbroeck, J., Ed.; Springer International Publishing: Cham, Switzerland, 2018; pp. 649–671. [Google Scholar] [CrossRef]
- Horovitz, A. Handbook of Flowering; CRC Press: Boca Raton, FL, USA, 1985; Volume 4, pp. 155–161. [Google Scholar] [CrossRef]
- Meynet, J. Ranunculus. In Physiology of Flower Bulbs: A Comprehensive Treatise on the Physiology and Utilization of Ornamental Flowering Bulbous and Tuberous Plants; De Hertogh, A.A., Le Nard, M., Eds.; Elsevier: Amsterdam, NL, USA, 1992; pp. 603–610. [Google Scholar]
- Kamenetsky, R.; Okubo, H. Production of Flower Bulbs in Regions with Warm Climates; Okubo, H., Miller, W.B., Chastagner, G.A., Eds.; International Society for Horticultural Science: Niigata, Japan, 2005; Volume 2, pp. 59–66. [Google Scholar]
- Beruto, M.; Fibiani, M.; Rinino, S.; Scalzo, R.L.; Curir, P. Plant development of Ranunculus asiaticus L. tuberous roots is affected by different temperature and oxygen conditions during storage period. Isr. J. Plant Sci. 2009, 57, 377–388. [Google Scholar] [CrossRef]
- Wang, W.; Su, X.; Tian, Z.; Liu, Y.; Zhou, Y.; He, M. Transcriptome profiling provides insights into dormancy release during cold storage of Lilium pumilum. BMC Genom. 2018, 19, 196. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liu, F.; Zhang, H.; Wu, G.; Sun, J.; Hao, L.; Ge, X.; Yu, J.; Wang, W. Sequence Variation and Expression Analysis of Seed Dormancy- and Germination-Associated ABA- and GA-Related Genes in Rice Cultivars. Front. Plant Sci. 2011, 2, 17. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ohkawa, K. Growth and Flowering of Ranunculus Asiaticus; Doss, R.P., Byther, R.S., Chastagner, G.A., Eds.; Acta Horticolturae: Seattle, WA, USA, 1986; Volume 1, pp. 165–172. [Google Scholar] [CrossRef]
- Farina, E.; Paterniani, T.; Suffia, G. Prove sperimentali sulla reattività del ranuncolo ibrido al fotoperiodo. Ann. Dell’istituto Sper. Per La Fioritura San Remo 1985, 16, 93–99. [Google Scholar]
- Karlsson, M. Producing Ravishing Ranunculus; Greenhouse Product News: Sparta, MI, USA, 2003; pp. 44–48. Available online: https://gpnmag.com/article/producing-ravishing-ranunculus/ (accessed on 1 October 2022).
- Carillo, P.; Arena, C.; Modarelli, G.C.; De Pascale, S.; Paradiso, R. Photosynthesis in Ranunculus asiaticus L.: The Influence of the Hybrid and the Preparation Procedure of Tuberous Roots. Front. Plant Sci. 2019, 10, 241. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Carillo, P.; Dell’Aversana, E.; Modarelli, G.C.; Fusco, G.M.; De Pascale, S.; Paradiso, R. Metabolic Profile and Performance Responses of Ranunculus asiaticus L. Hybrids as Affected by Light Quality of Photoperiodic Lighting. Front. Plant Sci. 2020, 11, 597823. [Google Scholar] [CrossRef] [PubMed]
- Carillo, P.; Modarelli, G.C.; Fusco, G.M.; Dell’Aversana, E.; Arena, C.; De Pascale, S.; Paradiso, R. Light spectral composition affects metabolic response and flowering in non-vernalized Ranunculus asiaticus L. Environ. Exp. Bot. 2021, 192, 104649. [Google Scholar] [CrossRef]
- Modarelli, G.C.; Arena, C.; Pesce, G.; Dell’Aversana, E.; Fusco, G.M.; Carillo, P.; De Pascale, S.; Paradiso, R. The role of light quality of photoperiodic lighting on photosynthesis, flowering and metabolic profiling in Ranunculus asiaticus L. Physiol. Plant. 2020, 170, 187–201. [Google Scholar] [CrossRef] [PubMed]
- Strand, Å.; Hurry, V.; Gustafsson, P.; Gardeström, P. Development of Arabidopsis thaliana leaves at low temperatures releases the suppression of photosynthesis and photosynthetic gene expression despite the accumulation of soluble carbohydrates. Plant J. 1997, 12, 605–614. [Google Scholar] [CrossRef]
- Björkman, O.; Demmig, B. Photon yield of O2 evolution and chlorophyll fluorescence characteristics at 77 K among vascular plants of diverse origins. Planta 1987, 170, 489–504. [Google Scholar] [CrossRef]
- Maxwell, K.; Johnson, G.N. Chlorophyll fluorescence—A practical guide. J. Exp. Bot. 2000, 51, 659–668. [Google Scholar] [CrossRef] [PubMed]
- Baker, N.R.; Rosenqvist, E. Applications of chlorophyll fluorescence can improve crop production strategies: An examination of future possibilities. J. Exp. Bot. 2004, 55, 1607–1621. [Google Scholar] [CrossRef] [Green Version]
- Muhlemann, J.K.; Klempien, A.; Dudareva, N. Floral volatiles: From biosynthesis to function. Plant Cell Environ. 2014, 37, 1936–1949. [Google Scholar] [CrossRef]
- Borghi, M.; Fernie, A.R. Floral metabolism of sugars and amino acids: Implications for pollinators’ preferences and seed and fruit set. Plant Physiol. 2017, 175, 1510–1524. [Google Scholar] [CrossRef] [Green Version]
- Rus, A.; Lee, B.-h.; Munoz-Mayor, A.; Sharkhuu, A.; Miura, K.; Zhu, J.-K.; Bressan, R.A.; Hasegawa, P.M. AtHKT1 facilitates Na+ homeostasis and K+ nutrition in planta. Plant Physiol. 2004, 136, 2500–2511. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Khan, N.; Ali, S.; Zandi, P.; Mehmood, A.; Ullah, S.; Ikram, M.; Ismail, M.A.S.; Babar, M. Role of sugars, amino acids and organic acids in improving plant abiotic stress tolerance. Pak. J. Bot. 2020, 52, 355–363. [Google Scholar] [CrossRef] [PubMed]
- Bernier, G.; Havelange, A.e.; Houssa, C.; Petitjean, A.; Lejeune, P. Physiological signals that induce flowering. Plant Cell 1993, 5, 1147. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Raven, J.A. Tansley review no. 2. Regulation of pH and generation of osmolarity in vascular plants: A cost-benefit analysis in relation to efficiency of use of energy, nitrogen and water. New Phytol. 1985, 101, 25–77. [Google Scholar] [CrossRef] [PubMed]
- Carillo, P.; Soteriou, G.A.; Kyriacou, M.C.; Giordano, M.; Raimondi, G.; Napolitano, F.; Di Stasio, E.; Mola, I.D.; Mori, M.; Rouphael, Y. Regulated salinity eustress in a floating hydroponic module of sequentially harvested lettuce modulates phytochemical constitution, plant resilience, and post-harvest nutraceutical quality. Agronomy 2021, 11, 1040. [Google Scholar] [CrossRef]
- Yanagida, M.; Mino, M.; Iwabuchi, M.; Ogawa, K.i. Reduced glutathione is a novel regulator of vernalization-induced bolting in the rosette plant Eustoma grandiflorum. Plant Cell Physiol. 2004, 45, 129–137. [Google Scholar] [CrossRef] [Green Version]
- Woodrow, P.; Ciarmiello, L.F.; Annunziata, M.G.; Pacifico, S.; Iannuzzi, F.; Mirto, A.; D’Amelia, L.; Dell’Aversana, E.; Piccolella, S.; Fuggi, A. Durum wheat seedling responses to simultaneous high light and salinity involve a fine reconfiguration of amino acids and carbohydrate metabolism. Physiol. Plant. 2017, 159, 290–312. [Google Scholar] [CrossRef] [PubMed]
- Carillo, P. GABA shunt in durum wheat. Front. Plant Sci. 2018, 9, 100. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bouche, N.; Fromm, H. GABA in plants: Just a metabolite? Trends Plant Sci. 2004, 9, 110–115. [Google Scholar] [CrossRef] [PubMed]
- Zhou, X.; Cao, K.; Zhang, Z.; Tan, Y.; Fan, H.; Xu, H. TMT-Based Proteomic Profiling Reveals the Molecular Mechanism in the Cold Tolerance of Rhododendron Aureum Georgi. Res. Sq. 2021. [Google Scholar] [CrossRef]
- von Caemmerer, S.; Farquhar, G.D. Some relationships between the biochemistry of photosynthesis and the gas exchange of leaves. Planta 1981, 153, 376–387. [Google Scholar] [CrossRef]
- Genty, B.; Briantais, J.-M.; Baker, N.R. The relationship between the quantum yield of photosynthetic electron transport and quenching of chlorophyll fluorescence. Biochim. Et Biophys. Acta (BBA)—Gen. Subj. 1989, 990, 87–92. [Google Scholar] [CrossRef]
- Bilger, W.; Björkman, O. Role of the xanthophyll cycle in photoprotection elucidated by measurements of light-induced absorbance changes, fluorescence and photosynthesis in leaves of Hedera canariensis. Photosynth. Res. 1990, 25, 173–185. [Google Scholar] [CrossRef]
- Annunziata, M.G.; Carillo, P.; Fuggi, A.; Troccoli, A.; Woodrow, P. Metabolic profiling of cauliflower under traditional and reduced tillage systems. Aust. J. Crop Sci. 2013, 7, 1317–1323. [Google Scholar]
- Dell’Aversana, E.; Hessini, K.; Ferchichi, S.; Fusco, G.M.; Woodrow, P.; Ciarmiello, L.F.; Abdelly, C.; Carillo, P. Salinity Duration Differently Modulates Physiological Parameters and Metabolites Profile in Roots of Two Contrasting Barley Genotypes. Plants 2021, 10, 307. [Google Scholar] [CrossRef]
- Salbitani, G.; Carillo, P.; Di Martino, C.; Bolinesi, F.; Mangoni, O.; Loreto, F.; Carfagna, S. Microalgae cross-fertilization: Short-term effects of Galdieria phlegrea extract on growth, photosynthesis and enzyme activity of Chlorella sorokiniana cells. J. Appl. Phycol. 2022, 34, 1957–1966. [Google Scholar] [CrossRef]
- Rouphael, Y.; Corrado, G.; Colla, G.; De Pascale, S.; Dell’Aversana, E.; D’Amelia, L.I.; Fusco, G.M.; Carillo, P. Biostimulation as a means for optimizing fruit phytochemical content and functional quality of tomato landraces of the San Marzano area. Foods 2021, 10, 926. [Google Scholar] [CrossRef] [PubMed]
- Bradford, N. A rapid and sensitive method for the quantitation microgram quantities of a protein isolated from red cell membranes. Anal. Biochem. 1976, 72, e254. [Google Scholar] [CrossRef]
- Carillo, P.; Colla, G.; Fusco, G.M.; Dell’Aversana, E.; El-Nakhel, C.; Giordano, M.; Pannico, A.; Cozzolino, E.; Mori, M.; Reynaud, H. Morphological and physiological responses induced by protein hydrolysate-based biostimulant and nitrogen rates in greenhouse spinach. Agronomy 2019, 9, 450. [Google Scholar] [CrossRef] [Green Version]
- Wellburn, A.R. The spectral determination of chlorophylls a and b, as well as total carotenoids, using various solvents with spectrophotometers of different resolution. J. Plant Physiol. 1994, 144, 307–313. [Google Scholar] [CrossRef]



{kind=link}
{kind=link}
| MDR | MBO | Significance | |||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|
| C | V2 | V4 | Mean | C | V2 | V4 | Mean | H | V | H × V | |
| NP | 6.07 ± 0.66 | 4.35 ± 1.04 | 4.71 ± 1.12 | 5.05 | 3.57 ± 0.20 | 3.24 ± 0.53 | 7.42 ± 0.74 | 4.74 | ns | ns | * |
| Fv/Fm | 0.79 ± 0.01 | 0.79 ± 0.01 | 0.79 ± 0.00 | 0.79 | 0.79 ± 0.01 | 0.81 ± 0.01 | 0.80 ± 0.01 | 0.8 | * | ns | ns |
| ΦPSII | 0.42 ± 0.08 | 0.57 ± 0.03 | 0.51 ± 0.02 | 0.5 | 0.50 ± 0.04 | 0.50 ± 0.02 | 0.48 ± 0.03 | 0.49 | ns | ns | ns |
| ETR | 72.38 ± 6.45 | 54.12 ± 5.15 | 95.85 ± 9.06 | 74.11 | 86.65 ± 13.68 | 91.14 ± 4.89 | 85.69 ± 8.52 | 87.83 | ns | ns | * |
| NPQ | 1.59 ± 0.40 | 1.13 ± 0.18 | 1.29 ± 0.23 | 1.37 | 1.57 ± 0.19 | 1.52 ± 0.25 | 1.27 ± 0.10 | 1.46 | ns | ns | ns |
| MDR | MBO | Significance | |||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|
| C | V2 | V4 | Mean | C | V2 | V4 | Mean | H | V | H × V | |
| Number of leaves (N./plant) | 24.3 ± 3.3 | 27.7 ± 5.4 | 17.5 ± 0.3 | 23.2 | 24.0 ± 3.1 | 30.7 ± 6.7 | 15.0 ± 1.2 | 23.2 | ns | * | ns |
| Total leaf area (cm2/plant) | 883.6 ± 41.5 | 578.8 ± 33.2 | 242.8 ± 49.8 | 568.4 | 646.5 ± 100.8 | 458.8 ± 84.9 | 107.7 ± 4.5 | 406.9 | ** | ** | * |
| Stem length (cm) | 57.5 ± 2.7 | 47.9 ± 1.5 | 38.8 ± 2.3 | 48.1 | 59.4 ± 2.7 | 57.1 ± 2.7 | 59.3 ± 6.0 | 58.6 | * | * | * |
| Stem diameter (mm) | 5.25 ± 0.36 | 3.45 ± 0.16 | 3.85 ± 0.21 | 4.18 | 5.26 ± 0.23 | 4.04 ± 0.30 | 2.64 ± 0.49 | 3.98 | ns | * | ns |
| Leaf D.W. (g/plant) | 3.32 ± 0.23 | 2.74 ± 0.47 | 0.92 ± 0.17 | 2.36 | 3.10 ± 0.45 | 2.18 ± 0.37 | 0.30 ± 0.04 | 1.86 | ns | * | ns |
| Root D.W. (g/plant) | 0.61 ± 0.12 | 0.32 ± 0.07 | 0.12 ± 0.03 | 0.36 | 0.48 ± 0.14 | 0.27 ± 0.10 | 0.08 ± 0.01 | 0.28 | ns | * | ns |
| Flower stem D.W. (g/plant) | 4.26 ± 0.54 | 1.70 ± 0.14 | 1.12 ± 0.12 | 2.36 | 3.25 ± 0.29 | 1.87 ± 0.17 | 1.10 ± 0.47 | 2.07 | ns | * | ns |
| MDR | MBO | Significance | |||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|
| C | V2 | V4 | Mean | C | V2 | V4 | Mean | H | V | H × V | |
| Vegetative phase | |||||||||||
| Glucose | 35.63 | 35.17 | 35.94 | 35.58 | 37.32 | 39.02 | 19.66 | 32.00 | ns | ns | ns |
| Fructose | 20.75 | 20.86 | 21.08 | 20.89 A | 22.82 | 23.59 | 23.47 | 23.29 B | ** | ns | ns |
| Sucrose | 0.05 a | 0.14 b | 0.24 c | 0.14 | 0.16 b | 0.05 a | 0.14 b | 0.12 | ns | * | * |
| Starch | 19.22 a | 21.17 a | 29.39 b | 23.26 | 24.64 c | 28.11 bc | 25.29 c | 26.01 | ns | ns | *** |
| Polyphenols | 7.81 a | 9.39 a | 8.37 a | 8.53 | 11.34 b | 11.93 b | 7.06 a | 10.12 | ns | * | *** |
| Chl a | 0.18 a | 0.21 a | 0.23 a | 0.21 A | 0.17 a | 0.18 a | 0.15 b | 0.17 B | *** | ns | * |
| Chl b | 0.05 a | 0.07 b | 0.06 b | 0.06 A | 0.05 a | 0.05 a | 0.04 a | 0.05 B | ** | ns | *** |
| Carotenoids | 0.04 a | 0.05 b | 0.05 b | 0.05 A | 0.04 a | 0.04 a | 0.04 a | 0.04 B | * | * | *** |
| Flowering phase | |||||||||||
| Glucose | 19.9 a | 28.27 b | 27.32 b | 25.16 | 35.7 b | 35.18 b | 21.71 a | 30.87 | ns | ns | * |
| Fructose | 21.50 | 20.95 | 18.07 | 20.17 A | 22.72 | 25.55 | 21.45 | 23.24 B | * | ns | ns |
| Sucrose | 0.05 a | 0.10 b | 0.14 b | 0.10 | 0.09 b | 0.05 a | 0.16 b | 0.10 | ns | *** | * |
| Starch | 19.82 a | 21.02 a | 28.2 a | 23.01 | 25.56 a | 39.31 b | 17.55 a | 27.47 | ns | ns | ** |
| Polyphenols | 12.87 a | 14.97 b | 10.05 c | 12.63 | 13.76 ab | 12.68 a | 8.82 c | 11.75 | ns | ** | *** |
| Chl a | 0.17 a | 0.15 b | 0.19 a | 0.17 | 0.15 b | 0.15 b | 0.20 a | 0.17 | ns | ** | * |
| Chl b | 0.05 a | 0.04 a | 0.06 b | 0.05 | 0.04 a | 0.04 a | 0.06 b | 0.05 | ns | ** | ns |
| Carotenoids | 0.04 a | 0.03 a | 0.04 b | 0.04 | 0.03 a | 0.03 a | 0.04 b | 0.04 | ns | * | ns |
| MDR | MBO | Significance | |||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|
| C | V2 | V4 | Mean | C | V2 | V4 | Mean | H | V | H × V | |
| Vegetative phase | |||||||||||
| Soluble proteins | 46.26 a | 48.50 a | 50.89 a | 48.55 A | 57.58 a | 51.86 a | 77.24 b | 62.23 B | * | ns | * |
| Alanine | 2.75 a | 3.41 a | 5.43 b | 3.86 | 3.52 a | 4.11 b | 3.96 a | 3.86 | ns | * | * |
| Asparagine | 49.03 a | 40.67 a | 62.11 a | 50.61 A | 6.32 b | 6.84 b | 28.29 b | 13.82 B | *** | ns | * |
| Aspartate | 7.82 | 11.85 | 11.67 | 10.45 | 10.69 | 10.74 | 10.27 | 10.56 | ns | ns | ns |
| GABA | 7.37 a | 8.72 ab | 11.36 bc | 9.15 | 6.10 a | 13.59 c | 0.47 d | 6.72 | ns | ns | *** |
| Glutamate | 14.98 a | 17.39 a | 26.71 c | 19.69 | 7.95 b | 7.33 b | 23.22 c | 12.83 | ns | ** | *** |
| Glutamine | 25.46 a | 27.84 a | 30.4 b | 27.90 | 16.84 c | 17.16 c | 37.50 b | 23.83 | ns | *** | *** |
| Glycine | 1.45 a | 0.61 b | 0.67 b | 0.91 A | 0.54 b | 0.62 b | 0.46 b | 0.54 B | * | ns | ** |
| MEA | 3.70 | 4.48 | 4.91 | 4.36 | 3.98 | 5.49 | 4.03 | 4.50 | ns | ns | ns |
| Ornithine | 0.46 a | 0.69 b | 0.63 b | 0.60 A | 0.63 b | 0.32 ac | 0.30 c | 0.42 B | * | ns | *** |
| Proline | 1.96 | 2.21 | 2.35 | 2.18 | 2.18 | 1.55 | 1.83 | 1.85 | ns | ns | ns |
| Serine | 4.12 a | 3.92 a | 5.87 b | 4.64 | 4.12 a | 7.74 c | 6.06 b | 5.98 | ns | * | *** |
| Threonine | 0.69 a | 0.84 b | 1.06 c | 0.86 | 0.64 ab | 1.24 c | 0.78 a | 0.89 | ns | * | *** |
| Total AA | 129.6 a | 133.4 a | 175.8 b | 146.3 A | 74.08 c | 91.45 c | 125.5 a | 97 B | *** | * | *** |
| Minor AA | 9.76 ad | 10.80 ac | 12.60 ab | 11.05 | 10.57 ad | 14.71 b | 8.30 cd | 11.19 | ns | ns | ** |
| BCAAs | 3.14 ab | 3.02 ab | 3.13 ab | 3.10 | 3.20 b | 4.20 c | 2.28 a | 3.23 | ns | ns | * |
| Flowering phase | |||||||||||
| Soluble Proteins | 31.58 a | 32.71 a | 45.43 b | 36.57 | 33.51 a | 40.39 a | 45.06 b | 39.65 | ns | * | ns |
| Alanine | 2.22 ab | 2.47 a | 1.27 b | 1.99 | 1.97 ab | 2.05 ab | 3.61 c | 2.54 | ns | ns | ** |
| Asparagine | 7.02 a | 2.34 b | 0.56 c | 3.31 | 4.04 b | 5.72 a | 13.03 d | 7.60 | * | ns | ** |
| Aspartate | 6.41 ac | 5.34 b | 4.29 c | 5.35 A | 7.99 a | 8.64 a | 7.77 ab | 8.14 B | *** | ns | * |
| GABA | 1.12 a | 1.84 b | 0.39 c | 1.11 | 1.08 a | 1.91 b | 2.55 d | 1.85 | ns | ns | *** |
| Glutamate | 10.98 | 15.53 | 4.81 | 10.44 | 9.74 | 8.98 | 25.04 | 14.59 | ns | ns | ns |
| Glutamine | 21.34 ab | 19.86 a | 16.49 c | 19.23 | 16.28 ac | 15.97 bc | 24.44 d | 18.89 | ns | ns | *** |
| Glycine | 0.21 ab | 0.21 b | 0.17 b | 0.20 | 0.14 b | 0.38 c | 0.31 ac | 0.28 | ns | ns | ** |
| MEA | 3.92 | 3.06 | 2.73 | 3.24 | 3.11 | 3.20 | 3.49 | 3.27 | ns | ns | ns |
| Ornithine | 0.57 | 0.50 | 0.49 | 0.52 | 0.54 | 0.64 | 0.64 | 0.60 | ns | ns | ns |
| Proline | 2.31 ab | 1.30 d | 1.90 ac | 1.83 | 1.17 d | 1.79 bcd | 2.38 a | 1.78 | ns | ns | *** |
| Serine | 3.77 ac | 2.73 bc | 2.10 c | 2.87 A | 5.96 a | 3.31 ac | 4.76 ab | 4.67 B | ** | ns | * |
| Threonine | 0.47 a | 0.70 b | 0.35 a | 0.51 | 0.30 a | 0.43 a | 0.77 b | 0.50 | ns | ns | ** |
| Total AA | 69.28 a | 62.59 a | 40.96 b | 57.61 | 58.27 ab | 59.38 ab | 101.44 c | 73.03 | ns | ns | *** |
| Minor AA | 8.94 a | 6.69 b | 5.40 b | 7.01 | 5.97 b | 6.36 b | 12.64 c | 8.32 | ns | ns | *** |
| BCAAs | 1.65 ab | 1.64 ab | 0.85 a | 1.38 | 1.07 a | 1.54 ab | 4.31 b | 2.31 | ns | ns | * |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Fusco, G.M.; Carillo, P.; Nicastro, R.; Modarelli, G.C.; Arena, C.; De Pascale, S.; Paradiso, R. Vernalization Procedure of Tuberous Roots Affects Growth, Photosynthesis and Metabolic Profile of Ranunculus asiaticus L. Plants 2023, 12, 425. https://0-doi-org.brum.beds.ac.uk/10.3390/plants12030425
Fusco GM, Carillo P, Nicastro R, Modarelli GC, Arena C, De Pascale S, Paradiso R. Vernalization Procedure of Tuberous Roots Affects Growth, Photosynthesis and Metabolic Profile of Ranunculus asiaticus L. Plants. 2023; 12(3):425. https://0-doi-org.brum.beds.ac.uk/10.3390/plants12030425
Chicago/Turabian StyleFusco, Giovanna Marta, Petronia Carillo, Rosalinda Nicastro, Giuseppe Carlo Modarelli, Carmen Arena, Stefania De Pascale, and Roberta Paradiso. 2023. "Vernalization Procedure of Tuberous Roots Affects Growth, Photosynthesis and Metabolic Profile of Ranunculus asiaticus L." Plants 12, no. 3: 425. https://0-doi-org.brum.beds.ac.uk/10.3390/plants12030425