
Effects of Bacterial and Fungal Inocula on Biomass, Ecophysiology, and Uptake of Metals of Alyssoides utriculata (L.) Medik.


 ,
,  and
and 
Abstract
:1. Introduction
2. Results
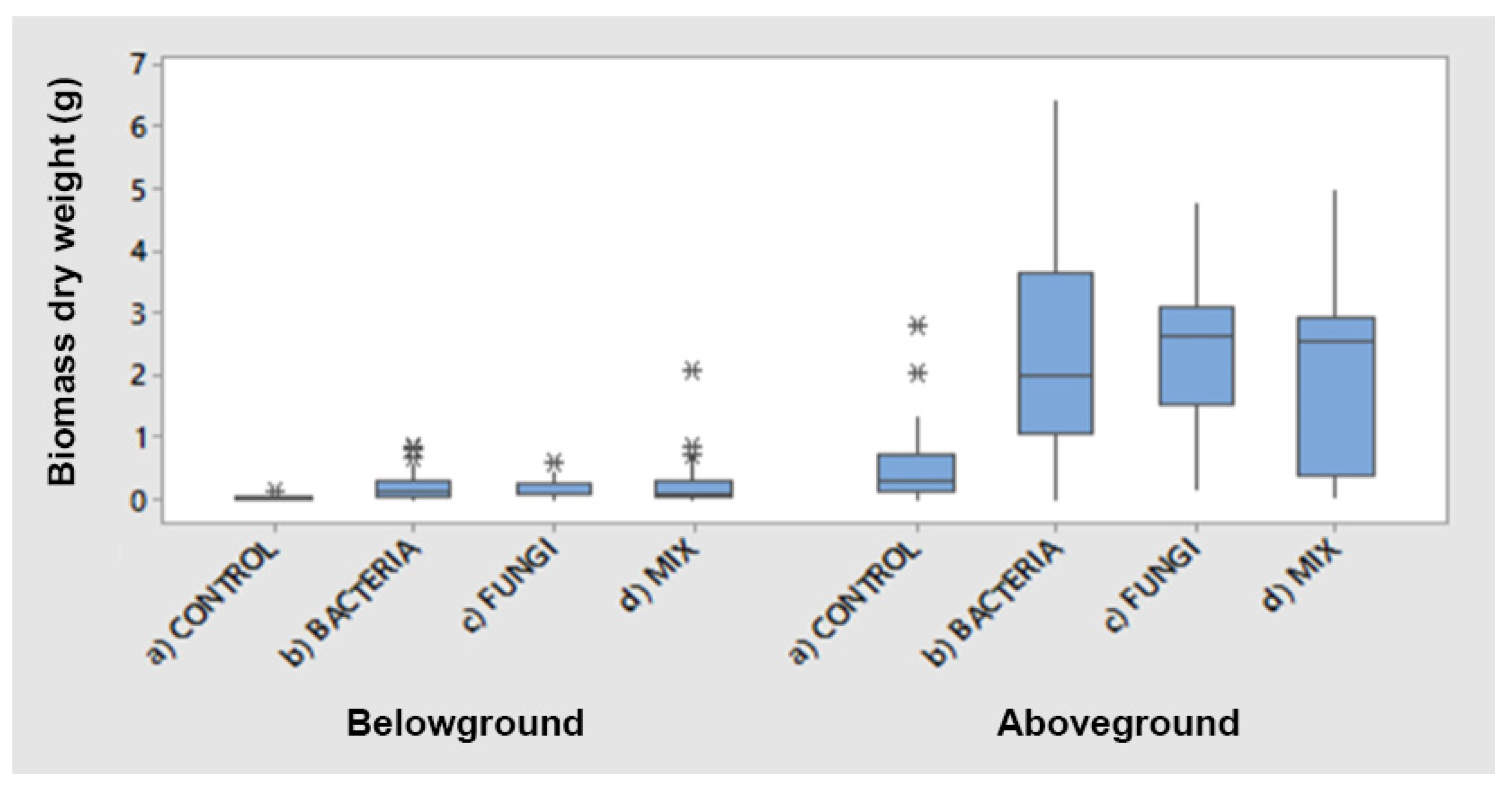
2.1. Biomass Production
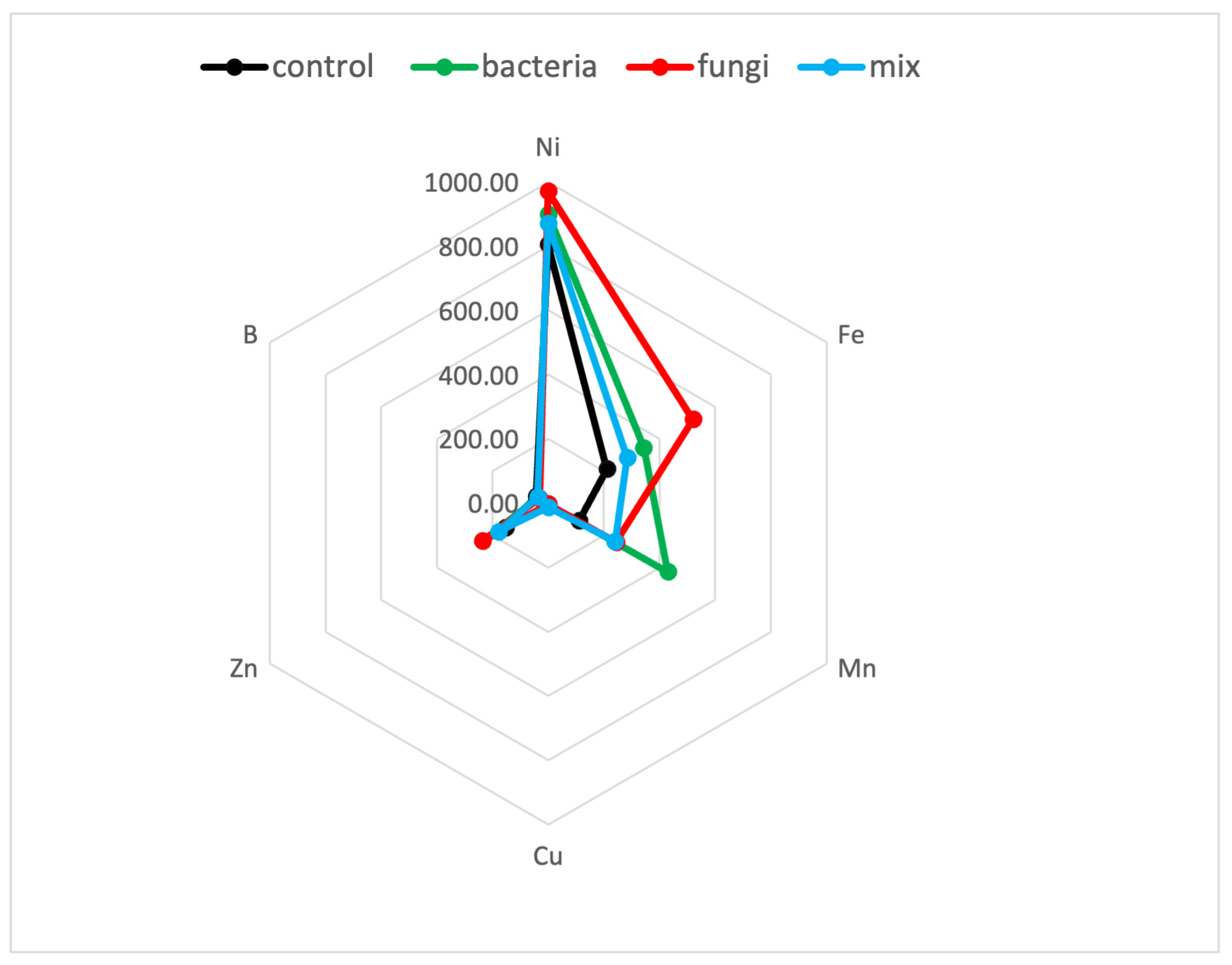
2.2. Heavy Metals and Nutrients Accumulation in the Aboveground Biomass
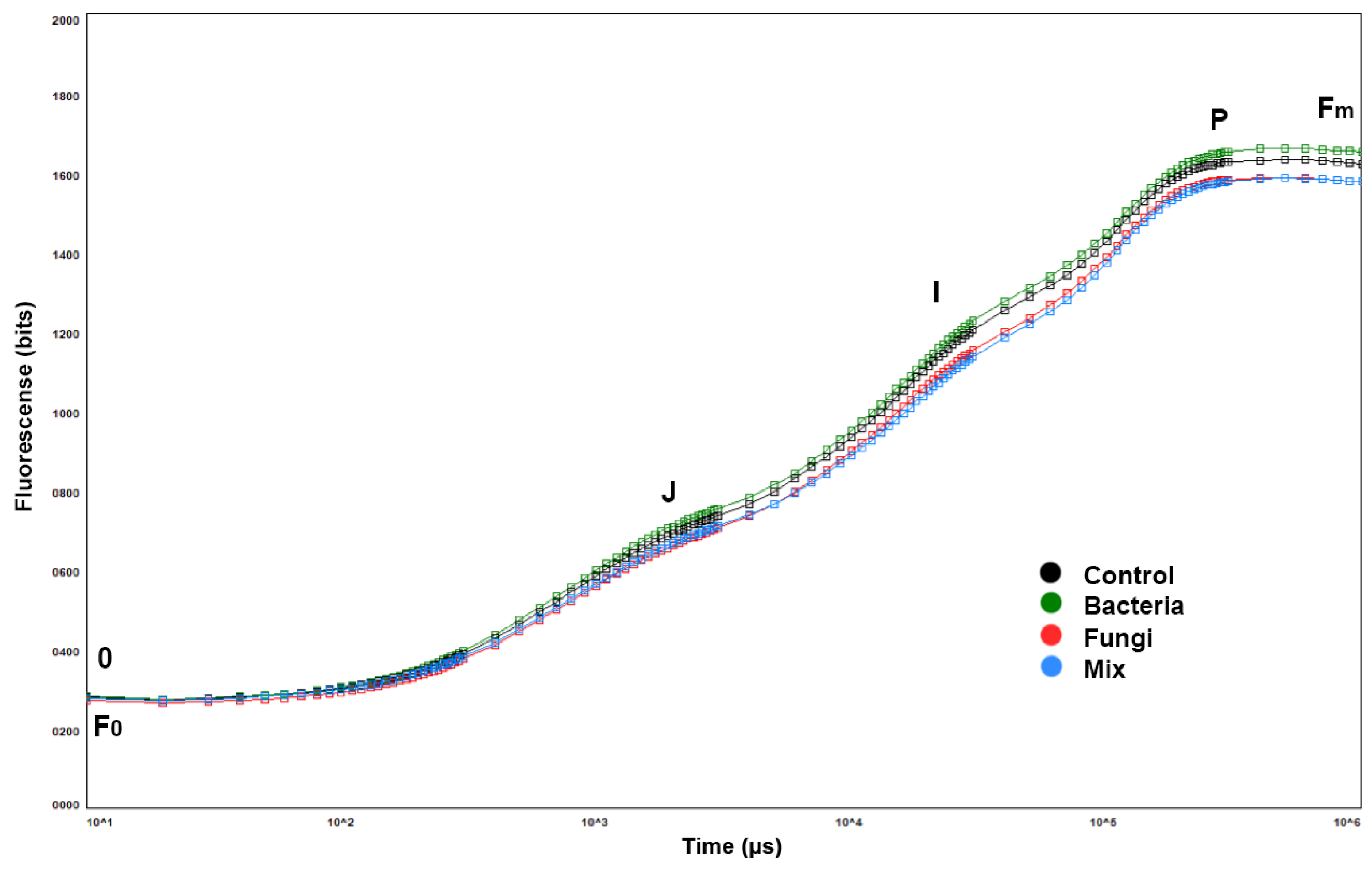
2.3. Photosynthetic Efficiency and Performance Index
3. Discussion
4. Materials and Methods
4.1. Plant Species and Soil Collection
4.2. Experimental Design
4.3. Soil Inoculation
4.4. Seeds Sowing
4.5. Transplants of A. utriculata
4.6. Ecophysiological Response
- F0 is the minimum fluorescence value, representing emissions from the excited chlorophyll molecules in the photosystem II’s antenna structure.
- Fm is the maximum fluorescence value, obtained after applying a saturation pulse to a dark-adapted leaf.
- Fv/Fm indicates the maximum quantum efficiency of the photosystem II, which indicates a plant’s photosynthetic performance.
- PI (performance index) indicates a sample’s viability.
4.7. Biomass Evaluation
4.8. Analysis of Elements
4.9. Data Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Chaoua, S.; Boussaa, S.; Gharmali, A.E.; Boumezzough, A. Impact of irrigation with wastewater on accumulation of heavy metals in soil and crops in the region of Marrakech in Morocco. J. Saudi Soc. Agric. Sci. 2018, 18, 429–436. [Google Scholar] [CrossRef]
- Ashraf, S.; Ali, Q.; Zahir, Z.A.; Ashraf, S.; Asghar, H.N. Phytoremediation: Environmentally sustainable way for reclamation of heavy metal polluted soils. Ecotoxicol. Environ. Saf. 2019, 174, 714–727. [Google Scholar] [CrossRef] [PubMed]
- United States Environmental Protection Agency (US EPA) Health Effects Assessment Summary Tables (HEAST); Annual Update; United States Environmental Protection Agency: Washington, DC, USA, 2016.
- EEA Report European Environment Agency (EEA). Environmental Pressures of Heavy Metal Releases from Europe’s Industry. Available online: https://www.eea.europa.eu/themes/industry/industrial-pollution-in-europe (accessed on 17 October 2022).
- Sitko, K.; Rusinowski, S.; Kalaji, H.M.; Szopiński, M.; Małkowski, E. Photosynthetic Efficiency as Bioindicator of Environmental Pressure in A. halleri. Plant Physiol. 2017, 175, 290–302. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yang, Y.; Zhang, L.; Huang, X.; Zhou, Y.; Quan, Q.; Li, Y.; Zhu, X. Response of Photosynthesis to Different Concentrations of Heavy Metals in Davidia Involucrata. PLoS ONE 2020, 15, e0228563. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Houri, T.; Khairallah, Y.; Zahab, A.A.; Osta, B.; Romanos, D.; Haddad, G. Heavy Metals Accumulation Effects on The Photosynthetic Performance of Geophytes in Mediterranean Reserve. J. King Saud Univ. Sci. 2020, 32, 874–880. [Google Scholar] [CrossRef]
- Chandra, R.; Kang, H. Mixed Heavy Metal Stress on Photosynthesis, Transpiration Rate, and Chlorophyll Content in Poplar Hybrids. For. Sci. Technol. 2016, 12, 55–61. [Google Scholar] [CrossRef] [Green Version]
- Goyal, D.; Yadav, A.; Prasad, M.; Singh, T.B.; Shrivastav, P.; Ali, A.; Dantu, P.K.; Mishra, S. Effects of Heavy Metals on Plant Growth: An Overview. Contam. Agric. 2020, 6, 79–101. [Google Scholar]
- Rascio, N.; Navari-Izzo, F. Heavy metal hyperaccumulating plants: How and why do they do it? And what makes them so interesting? Plant Sci. 2011, 180, 169–181. [Google Scholar] [CrossRef]
- Brooks, R.R.; Lee, J.; Reeves, R.D.; Jaffre, T. Detection of nickeliferous rocks by analysis of herbarium specimens of indicator plants. J. Geochem. Expl. 1977, 7, 49–57. [Google Scholar] [CrossRef]
- Van der Ent, A.; Baker, A.J.M.; Reeves, R.D.; Pollard, A.J.; Schat, H. Hyperaccumulators of metal and metalloid trace elements: Facts and fiction. Plant Soil 2013, 362, 319–334. [Google Scholar] [CrossRef]
- Ali, H.; Khan, E.; Sajad, M.A. Phytoremediation of heavy metals—Concepts and applications. Chemosphere 2013, 91, 869–881. [Google Scholar] [CrossRef]
- Diarra, I.; Kotra, K.K.; Prasad, S. Assessment of biodegradable chelating agents in the phytoextraction of heavy metals from multi–metal contaminated soil. Chemosphere 2021, 273, 128483. [Google Scholar] [CrossRef] [PubMed]
- Teng, Y.; Zhishuai, L.; Yu, A.; Guan, W.; Yu, H.; Zou, L. Phytoremediation of cadmium-contaminated soils by Solanum nigrum L. enhanced with biodegradable chelating agents. Environ. Sci. Pollut. Res. 2022, 37, 1–10. [Google Scholar] [CrossRef] [PubMed]
- Emamverdian, A.; Ding, Y.; Mokhberdoran, F.; Ahmad, Z. Mechanisms of Selected Plant Hormones under Heavy Metal Stress. Pol. J. Environ. Stud. 2021, 30, 1–11. [Google Scholar] [CrossRef]
- Bolan, N.; Kunhikrishnan, A.; Thangarajan, R.; Kumpiene, J.; Park, J.; Makino, T.; Kirkham, M.B.; Scheckel, K. Remediation of heavy metal(loid)s contaminated soils—To mobilize or not to immobilize? J. Haz. Mat. 2013, 266, 141–166. [Google Scholar] [CrossRef] [PubMed]
- Cassina, L.; Tassi, E.; Giorgetti, L.; Remorini, D.; Chaney, R.L.; Barbafieri, M. Exogenous Cytokinin Treatments of an NI- Hyper Accumulator, Alyssum murale, Grown in a Serpentine Soil: Implications for Phytoextraction. Int. J. Phytorem. 2011, 13, 90–101. [Google Scholar] [CrossRef]
- Cabello-Conejo, M.I.; Centofanti, T.; Kidd, P.S.; Prieto-Fernàndez, A.; Chaney, R.L. Evaluation of plant growth regulators to increase nickel phytoextraction by Alyssum species. Int. J. Phytorem. 2013, 15, 365–375. [Google Scholar] [CrossRef]
- Wang, L.; Hou, D.; Shen, Z.; Zhu, J.; Jia, X.; Ok, Y.S.; Tack, F.M.G.; Rinklebe, J. Field trials of phytomining and phytoremediation: A critical review of influencing factors and effects of additives. Crit. Rev. Environ. Sci. Technol. 2020, 50, 2724–2774. [Google Scholar] [CrossRef]
- Pérez-Àlvarez, S.; Ardisana, E.F.H.; Magallanes-Tapia, M.A.; Bonilla, C.M.E.; Garcia, C.U.; Gonzàles, M.M.; Guerrero-Morales, S. Microorganisms Used as Growth Regulators in Modern Agriculture. Benef. Microorg. Agric. 2022, 37–84. [Google Scholar]
- Muktar, S.; Bhatti, H.N.; Khalid, M.; Anwar Ul Haq, M.; Shahzad, S.M. Potential of sunflower (Helianthus annuus L.) for phytoremediation on Nickel (Ni) and lead (Pb) contaminated water. Pak. J. Bot. 2010, 42, 4017–4026. [Google Scholar]
- Shinta, Y.C.; Zaman, B.; Sumiyati, S. Citric Acid and EDTA as chelating agents in phytoremediation of heavy metal in polluted soil: A review. In IOP Conference Series: Earth and Environmental Science; IOP Publishing: Bristol, UK, 2021; p. 896. [Google Scholar]
- Hai, N.N.S.; Sanderson, P.; Qi, F.; Du, J.; Nong, N.N.; Bolan, N.; Naidu, R. Effects of chelates (EDTA, EDDS, NTA) on phytoavailability of heavy metals (As, Cd, Cu, Pb, Zn) using ryegrass (Lolium multiflorum Lam.). Environ. Sci. Pollut. Res. 2022, 29, 42102–42116. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Y.; Zhao, S.; Liu, S.; Pens, J.; Zhang, H.; Zhao, Q.; Zheng, L.; Chen, Y.; Shen, Z.; Xu, X.; et al. Enhancing Phytoremediation of Heavy Metals by Combining Hyperaccumulator and Heavy Metal-Resistant Plant Growth-Promoting Bacteria. Front. Plant Sci. 2022, 13. [Google Scholar] [CrossRef] [PubMed]
- Bulgarelli, D.; Schlaeppi, K.; Spaepen, S.; van Themaat, E.V.L.; Schulze-Lefert, P. Structure and Functions of the Bacterial Microbiota of Plants. Annu. Rev. Plant Biol. 2013, 64, 807–838. [Google Scholar] [CrossRef]
- Backer, R.; Rokem, J.S.; Ilangumaran, G.; Lamont, J.; Prasickova, D.; Ricci, E.; Subramanian, S.; Smith, D.L. Plant Growth-Promoting Rhizobacteria: Context, Mechanisms of Action, and Roadmap to Commercialization of Biostimulants for Sustainable Agriculture. Front. Plant. Sci. 2018, 9, 1473. [Google Scholar] [CrossRef] [Green Version]
- Wang, Y.; Narayanan, M.; Shi, X.; Chen, X.; Li, Z.; Natarajan, D.; Ma, Y. Plant growth-promoting bacteria in metal-contaminated soil: Current perspectives on remediation mechanisms. Front. Microb. 2022, 13, 966226. [Google Scholar] [CrossRef] [PubMed]
- Glick, B.R.; Gamalero, E. Recent developments in the Study of Plant Microbiomes. Microorganisms 2021, 9, 1533. [Google Scholar] [CrossRef]
- Mishra, J.; Singh, R.; Arora, N.K. Alleviation of Heavy Metal Stress in Plants and Remediation of Soil by Rhizosphere Microrganisms. Front. Microbiol. 2017, 8, 1706. [Google Scholar] [CrossRef] [Green Version]
- Wang, N.R.; Wiesmann, C.L.; Melnyk, R.A.; Hossain, S.S.; Chi, M.-H.; Martens, K.; Craven, K.; Haney, C.H. Commensal Pseudomonas fluorescens Strains Protect Arabidopsis from Closely Related Pseudomonas Pathogens in a Colonization-Dependent Manner. Pl. Microb. 2022, 13, e02892-21. [Google Scholar] [CrossRef]
- Rosatto, S.; Roccotiello, E.; Di Piazza, S.; Cecchi, G.; Greco, G.; Zotti, M.; Vezzulli, L.; Mariotti, M.G. Rhizosphere response to nickel in a facultative hyperaccumulator. Chemosphere 2019, 232, 243–253. [Google Scholar] [CrossRef]
- Cecchi, M.G.; Di Piazza, S.; Rosatto, S.; Mariotti, M.G.; Roccotiello, E.; Zotti, M. A Mini-Review on the Co-growth and Interactions among Microorganisms (Fungi and Bacteria) From Rhizosphere of Metal-Hyperaccumulators. Front. Fungal Biol. 2021, 2, 60. [Google Scholar] [CrossRef]
- Wu, J.; Shi, Z.; Zhu, J.; Cao, A.; Fang, W.; Yan, D.; Wang, Q.; Li, Y. Taxonomic response of bacterial and fungal populations to biofertilizers applied to soil or substrate in greenhouse-grown cucumber. Sci. Rep. 2022, 12, 18522. [Google Scholar] [CrossRef] [PubMed]
- Nishu, S.D.; No, J.H.; Lee, T.K. Transcriptional Response and Plant Growth Promoting Activity of Pseudomonas fluorescens DR397 under Drought Stress Conditions. ASM J. Microb. Spect. 2022, 10, e00979-22. [Google Scholar] [CrossRef] [PubMed]
- Tsegaye, Z.; Alemu, T.; Desta, F.A.; Assefa, F. Plant growth-promoting rhizobacterial inoculation to improve growth, yield, and grain nutrient uptake of teff varieties. Front. Microbiol. 2022, 13, 896770. [Google Scholar] [CrossRef] [PubMed]
- Hossain, M.M.; Sultana, F. Application and Mechanisms of Plant Growth Promoting Fungi (PGPF) for Phytostimulation. Org. Agric. 2020, 1–31. [Google Scholar]
- La Spada, F.; Stracquadanio, C.; riolo, M.; Pane, A.; Cacciola, S.O. Trichoderma Counteracts the Challenge of Phytophthora nicotianae Infections on Tomato by Modulating Plant Defense Mechanisms and the Expression of Crinkler, Necrosis-Inducing Phytophthora Protein 1, and Cellulose-Binding Elicitor Lectin Pathogenic Effectors. Front. Plant. Sci. 2020, 11, 583539. [Google Scholar]
- Ozimek, E.; Hanaka, A. Mortierella Specie sas the Plant-Growth-Promoting Fungi Present in the Agricultural Soils. Agriculture 2021, 11, 7. [Google Scholar] [CrossRef]
- Cecchi, G.; Roccotiello, E.; Di Piazza, S.; Riggi, A.; Mariotti, M.G.; Zotti, M. Assessment of Ni accumulation capability by fungi for a possible approach to remove metals from soils and waters. J. Environ. Sci. Health 2017, 52, 166–170. [Google Scholar] [CrossRef]
- Rosatto, S.; Cecchi, G.; Roccotiello, E.; Di Piazza, S.; Di Cesare, A.; Mariotti, M.G.; Vezzulli, L.; Zotti, M. Frenemies: Interactions between Rhizospheric Bacteria and Fungi from Metalliferous Soils. Life 2021, 202, 273. [Google Scholar] [CrossRef]
- Durand, A.; Leglize, P.; Benizri, E. Are endophytes essential partners for plants and what are the prospects for metal phytoremediation? Plant Soil 2021, 460, 1–30. [Google Scholar] [CrossRef]
- Kumar, A.; Voropaeva, O.; Maleva, M.; Panikovskaya, K.; Borisova, G.; Rajkumar, M.; Bruno, L.B. Bioaugmentation with copper tolerant endophyte Pseudomonas lurida strain EOO26 for improved plant growth and copper phytoremediation by Helianthus annuus. Chemosphere 2021, 266, 128983. [Google Scholar] [CrossRef]
- Hossain, M.M.; Sultana, F.; Islam, S. Plant Growth-Promoting Fungi (PGPF): Phytostimulation and Induced Systemic Resistance. Plant-Microbe Interact. Agro-Ecol. Perspect. 2017, 2, 135–191. [Google Scholar]
- Cabello-Conejo, M.I.; Becarra-Castro, C.; Prieto-Fernàndez, A.; Monterosso, C.; Saavedra-Ferro, A.; Kidd, P.S. Rhizobacterial inoculants can improve nickel phytoestraction by the hyperaccumulator Alyssum pintodasilvae. Plant Soil 2014, 379, 35–50. [Google Scholar] [CrossRef]
- Vrabl, P.; Fuchs, V.; Pichler, B.; Schinagl, C.W.; Burgstaller, W. Organic acid excretion in Penicillium ochrochloron increases with ambient pH. Front. Microbiol. 2012, 3, 121. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Abbaszadeh-Dahaii, P.; Atajan, F.A.; Omidvari, M.; Tahan, V.; Kariman, K. Mitigation of Copper Stress in Maize (Zea mays) and Sunflower (Helianthus annuus) Plant by Copper-resistant Pseudomonas Strains. Curr. Microb. 2021, 78, 1335–1343. [Google Scholar] [CrossRef] [PubMed]
- Babu, A.G.; Kim, S.W.; Yadav, D.R.; Hyum, U.; Adhikar, M.; Lee, Y.S. Penicillium menonorum: A Novel Fungus to Promote Growth and Nutrient Management in Cucumber Plants. Mycobiology 2015, 43, 49–56. [Google Scholar] [CrossRef] [Green Version]
- Ahemad, M. Remediation of metalliferous soil through the heavy metal resistant plant growth promoting bacteria: Paradigms and prospects. Arab. J. Chem. 2017, 12, 1365–1377. [Google Scholar] [CrossRef]
- Tarroum, M.; Romdhane, W.B.; Al-Qurainy, F.; Ali, A.A.M.; Al-Doss, A.; Fki, L.; Hassairi, A. A novel PGPF Penicillium olsonii isolated from the rhizosphere of Aeluropus littoralis promotes plant growth, enhances salt stress tolerance, and reduces chemical fertilizer inputs in hydroponic system. Front. Microbiol. 2022, 13, 996054. [Google Scholar] [CrossRef]
- Hemashenpagam, N.; Selvaraj, T. Effect of arbuscular mychorrhizal (AM) fungus and plant growth promoting rhizomicroorganisms (PGPR’s) on medicinal plant Solanum viarum seedlings. J. Environ. Biol. 2011, 32, 579–583. [Google Scholar]
- Kaur, T.; Devi, R.; Kumar, S.; Sheikh, I.; Kour, D.; Yadav, A.N. Microbial consortium with nitrogen fixing and mineral solubilizing attributes for growth of barley (Hordeum vulgare L.). Heliyon 2022, 8, e09326. [Google Scholar] [CrossRef]
- Wu, Q.-S.; Srivastava, A.K. Rhizosphere Microbial Comunities: Isolation, Characterization, and Value Addition for Substrate Development. Adv. Citrus Nutr. 2012, 169–194. [Google Scholar]
- Pidlisnyuk, V.; Mamirova, A.; Pranaw, K.; Stadnik, V.; Kuráň, P.; Trögl, J.; Shapoval, P. Miscanthus × giganteus Phytoremediation of Soil Contaminated with Trace Elements as Influenced by the Presence of Plant Growth-Promoting Bacteria. Agronomy 2022, 12, 771. [Google Scholar] [CrossRef]
- Marescotti, P.; Comodi, P.; Crispini, L.; Gigli, L.; Zucchini, A.; Fornasaro, S. Potentially Toxic Elements in Ultramafic Soils: A study from Metamorphic Ophiolites of the Voltri Massif (Western Alps, Italy). Minerals 2019, 9, 502. [Google Scholar] [CrossRef] [Green Version]
- Wei, Z.; Hillier, S.; Gadd, G.M. Biotransformation of manganese oxides by fungi: Solubilization and production of manganese oxalate biominerals Environ. Microbiology 2012, 14, 1744–1753. [Google Scholar] [CrossRef] [PubMed]
- Martino, E.; Perotto, S.; Parsons, R.; Gadd, G.M. Solubilization of insoluble inorganic zinc compounds by ericoid mycorrhizal fungi derived from heavy metal polluted sites Soil. Biol. Biochem. 2003, 35, 133–141. [Google Scholar] [CrossRef]
- Płociniczak, T.; Kukla, M.; Watroba, R.; Piotrowska-Seget, Z. The effect of soil bioaugmentation with strains of Pseudomonas on Cd, Zn and Cu uptake by Sinapis alba L. Chemosphere 2013, 91, 1332–1337. [Google Scholar] [CrossRef] [PubMed]
- Petriccione, M.; Di Patre, D.; Ferrante, P.; Papa, S.; Bartoli, G.; Fioretto, A.; Scortichini, M. Effects of Pseudomonas fluorescens Seed Bioinoculation on Heavy Metal Accumulation for Mirabilis jalapa Phytoextraction in Smelter-Contaminated Soil. Water Air Soil Pollut. 2013, 224, 1645. [Google Scholar] [CrossRef] [Green Version]
- Martínez, J.I.; Gómez-Garrido, M.; Gómez-López, M.D.; Faz, Á.; Martínez-Martínez, S.; Acosta, J.A. Pseudomonas fluorescens affects nutrient dynamics in plant-soil system for melon production. Chil. J. Agric. Res. 2019, 79, 223–233. [Google Scholar] [CrossRef] [Green Version]
- Wu, Y.; Ma, L.; Liu, Q.; Sikder, M.M.; Vestergård, M.; Zhoud, K.; Wang, Q.; Yang, X.; Feng, Y. Pseudomonas fluorescens promote photosyntesis, carbon fixation and cadmium phytoremediation of hyperaccumulator Sedum alfredii. Sci. Total Environ. 2020, 726, 138554. [Google Scholar] [CrossRef]
- Roccotiello, E.; Serrano, H.C.; Mariotti, M.G.; Branquinho, C. The impact of Ni on the physiology of a Mediterranean Ni-hyperaccumulating plant. Environ. Sci. Pollut. Res. 2016, 23, 243–253. [Google Scholar] [CrossRef]
- Domokos-Szabolcsy, É.; Fári, M.; Márton, L.; Czakó, M.; Veres, S.; Elhawat, N.; Antal, G.; El-Ramady, H.; Zsíros, O.; Garab, G.; et al. Selenate tolerance and selenium hyperaccumulation in the monocot giant reed (Arundo donax), a biomass crop plant with phytoremediation potential. Environ. Sci. Pollut. Res. Int. 2018, 25, 31368–31380. [Google Scholar] [CrossRef]
- Sytar, O.; Ghosh, S.; Malinska, H.; Zivcak, M.; Brestic, M. Physiological and molecular mechanisms of metal accumulation in hyperaccumulator plants. Phys. Plant. 2021, 173, 148–166. [Google Scholar] [CrossRef]
- Seregin, I.V.; Erlikh, N.T.; Kozhenikova, A.D. Nichel and zinc accumulation capacities and tolerance to these metals in the excluder Thlaspi arvense and the hyperaccumulator Noccaea caerulescens. Russ. J. Plant Phys. 2014, 61, 204–214. [Google Scholar] [CrossRef]
- Seregin, I.V.; Kozhevnikova, A.D. Physiological Role of Nichel and Its Toxic effects on Higher Plants. Russ. J. Plant Physiol. 2006, 56, 285–308. [Google Scholar]
- Tchounwou, P.B.; Yedjou, C.G.; Patlolla, A.K.; Sutton, D.J. Heavy Metal Toxicity and the Environment. Mol. Clin. Environ. Toxicol. 2012, 101, 133–164. [Google Scholar]
- Paunov, M.; Koleva, L.; Vassilev, A.; Vangronsveld, J.; Goltsev, V. Effects of Different Metals on Photosynthesis: Cadmium and Zinc Affect Chlorophyll Fluorescence in Durum Wheat. Int. J. Mol. Sci. 2018, 19, 787. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Marchand, L.; Lamy, P.; Bert, V.; Quintela-Sabaris, C.; Mench, M. Potential of Ranunculus acris L. for Biomonitoring Trace Element Contamination of Riverbank Soils: Photosystem II Activity and Phenotypic Responses for Two Soil Series. Environ. Sci. Pollut. Res. 2016, 23, 3104–3119. [Google Scholar] [CrossRef]
- ESCONET Seed Collecting Manual for Wild Species, 1st. ed.; Royal Botanic Gardens, Kew: Richmond, UK; Universidad Politècnica de Madrid: Madrid, Spain, 2009.






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Treatments | T-Test | |
|---|---|---|
| Belowground | Aboveground | |
| Control vs. Bacteria | 0.001 b | 0.000 b |
| Control vs. Fungi | 0.000 b | 0.000 b |
| Control vs. Mix | 0.010 b | 0.000 b |
| Bacteria vs. Mix | ns | ns |
| Fungi vs. Mix | ns | ns |
| Treatments | p Value of Elements’ Concentration in the Aboveground Biomass | |||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Ca | Mg | K | P | S | N | Ni | Fe | Mn | Cu | Zn | B | |
| Control vs. Bacteria | ns | ns | ns | 0.000 b | 0.003 a | ns | ns | ns | 0.000 b | 0.003 a | 0.001 b | 0.015 a |
| Control vs. Fungi | ns | ns | ns | 0.016 b | 0.001 a | ns | ns | ns | 0.002 b | 0.001 a | 0.003 b | 0.001 a |
| Control vs. Mix | ns | ns | ns | 0.000 b | 0.002 a | 0.024 a | ns | ns | 0.001 b | 0.001 b | ns | ns |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Priarone, S.; Romeo, S.; Di Piazza, S.; Rosatto, S.; Zotti, M.; Mariotti, M.; Roccotiello, E. Effects of Bacterial and Fungal Inocula on Biomass, Ecophysiology, and Uptake of Metals of Alyssoides utriculata (L.) Medik. Plants 2023, 12, 554. https://0-doi-org.brum.beds.ac.uk/10.3390/plants12030554
Priarone S, Romeo S, Di Piazza S, Rosatto S, Zotti M, Mariotti M, Roccotiello E. Effects of Bacterial and Fungal Inocula on Biomass, Ecophysiology, and Uptake of Metals of Alyssoides utriculata (L.) Medik. Plants. 2023; 12(3):554. https://0-doi-org.brum.beds.ac.uk/10.3390/plants12030554
Chicago/Turabian StylePriarone, Silvia, Sara Romeo, Simone Di Piazza, Stefano Rosatto, Mirca Zotti, Mauro Mariotti, and Enrica Roccotiello. 2023. "Effects of Bacterial and Fungal Inocula on Biomass, Ecophysiology, and Uptake of Metals of Alyssoides utriculata (L.) Medik." Plants 12, no. 3: 554. https://0-doi-org.brum.beds.ac.uk/10.3390/plants12030554