
Generation of Sesame Mutant Population by Mutagenesis and Identification of High Oleate Mutants by GC Analysis
1
Plant Genetic Resources Conservation Unit, USDA-ARS, 1109 Experiment Street, Griffin, GA 30223, USA
2
Wheat Health, Genetics, and Quality Research, USDA-ARS, 291 Clark Hall, Pullman, WA 99164, USA
*
Author to whom correspondence should be addressed.
Plants 2023, 12(6), 1294; https://0-doi-org.brum.beds.ac.uk/10.3390/plants12061294
Submission received: 19 November 2022
/
Revised: 31 December 2022
/
Accepted: 10 March 2023
/
Published: 13 March 2023
(This article belongs to the Special Issue Mutagenesis in the Age of Next-Generation-Sequencing and Genome Editing)
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:Sesame is one of the important oilseed crops in the world. Natural genetic variation exists in the sesame germplasm collection. Mining and utilizing the genetic allele variation from the germplasm collection is an important approach for seed quality improvement. The sesame germplasm accession, PI 263470, which has a significantly higher level of oleic acid (54.0%) than the average (39.5%), was identified by screening the entire USDA germplasm collection. The seeds from this accession were planted in a greenhouse. Leaf tissues and seeds were harvested from individual plants. DNA sequencing of the coding region of the fatty acid desaturase gene (FAD2) confirmed that this accession contained a natural mutation of G425A which may correspond to the deduced amino acid substitution of R142H leading to the high level of oleic acid, but it was a mixed accession with three genotypes (G/G, G/A, and A/A at the position). The genotype with A/A was selected and self-crossed for three generations. The purified seeds were used for EMS-induced mutagenesis to further enhance the level of oleic acid. A total of 635 M2 plants were generated from mutagenesis. Some mutant plants had significant morphological changes including leafy flat stems and others. M3 seeds were used for fatty acid composition analysis by gas chromatography (GC). Several mutant lines were identified with high oleic acid (70%). Six M3 mutant lines plus one control line were advanced to M7 or M8 generations. Their high oleate traits from M7 or M8 seeds harvested from M6 or M7 plants were further confirmed. The level of oleic acid from one mutant line (M7 915-2) was over 75%. The coding region of FAD2 was sequenced from these six mutants, but no mutation was identified. Additional loci may contribute to the high level of oleic acid. The mutants identified in this study can be used as breeding materials for sesame improvement and as genetic materials for forward genetic studies.
1. Introduction
Sesame is an important oilseed crop with a long history of cultivation (over 3000 years). Seeds, leaves, and oil from sesame have been utilized and consumed by humans as a vegetable and food ingredient for about 6000 years [1]. Sesame seeds contain 48–55% oil, 20–28% protein, 14–16% sugars, 6–8% fibers, and other nutritional and/or bioactive compounds with beneficial effects to human health such as vital minerals, vitamins, phytosterols, tocopherols and lignans [2]. The beneficial effects to human health from these bioactive compounds include the prevention of degenerative diseases such as cancer, cardiovascular diseases, atherosclerosis, and the process of aging. In the traditional Chinese and Indian systems of medicine, sesame seed and oil also played an important role in treatments such as rubbing sesame oil on the skin to sooth minor burns, aid in healing chronic skin disease, and consumption of roasted sesame seeds by new moms for producing breast milk. Significant variation for oil content, protein content, lignan, tocopherol, and fatty acid concentrations was found among eight sesame genotypes. Oil, protein, sesamin and sesamolin, tocopherol, oleic and linoleic, and the minor unsaturated fatty acids were found ranging from 29.43 to 54.69%, 13.92 to 21.76%, 0.55 to 8.98 mg/g, 0 to 239.58 µg/g, 26.6 to 54.85%, and 0.13 to 0.89%, respectively [3]. Since sesame is a highly nutritional and valuable crop, some basic genetic and genomic research work has been conducted on sesame for exploring its utilization potential and developing new cultivars.
Cultivated sesame (Sesamum indicum L.) is a diploid species (2n = 2x = 26) with a relatively small nuclear genome size (n = 354 Mb) [4,5]. Its genome has been sequenced using Illumina technology [6]. Some candidate genes involved in oil biosynthetic pathways have been identified by the analysis of comparative genomics and transcriptomics [7,8], and an integrated database (i.e., genetic information combined with comprehensive phenotypic information) for the functional genomics of sesame (SesameFG) has also been made publicly available [9]. Recently, gene-editing using the CRISPR/Cas9 (clustered regularly interspaced short palindromic repeats-associated protein 9) system with hair root transformation was also successfully applied for the targeted mutagenesis of genes in sesame for sesamin and sesamolin biosynthesis [10].
Improving seed nutritional quality (for example, elevation of oleic/linoleic acid ratio, O/L) is one of the main objectives for oilseed crop breeding programs. Seeds with the high O/L ratio (i.e., high oleic acid content and low linoleic acid content) can not only increase the seed shelf-life for preservation but also provide high quality sesame products beneficial to human health including reducing blood pressure and risk of cardiovascular diseases. High oleate (about 80%) cultivars/traits have been developed and identified in several major oilseed crops such as peanuts, soybeans, canola, and safflower [11,12,13,14] through breeding programs. However, little work has been reported in identification and development of high oleate cultivars in sesame except for some preliminary sesame mutagenesis work on other traits [15,16]. Screening the germplasm collection and EMS-mutagenesis are two efficient approaches, respectively, for identification of naturally occurring high oleate mutants and creation of newly induced high oleate mutants. Our goal was to develop high oleate germplasm lines in sesame by using these two approaches synergistically (i.e., first identification of a natural high oleate mutant and then mutagenesis of this natural high oleate mutant for further enhancing the oleic acid level to about 80%). Therefore, the objectives of this study are to (1) screen the USDA sesame germplasm collection to identify natural high oleate accessions, (2) conduct mutagenesis of the natural high oleate mutant to further enhance the oleic acid level, and (3) characterize the high oleate mutants to generate useful genetic materials for breeders and geneticists to use for development of new sesame cultivars and identification of corresponding genes for the high oleate trait.
2. Results and Discussion
2.1. Identification of Naturally High Oleate Mutants from the Germplasm Collection
2.1.1. Identification of Naturally High Oleate Mutants by GC Analysis
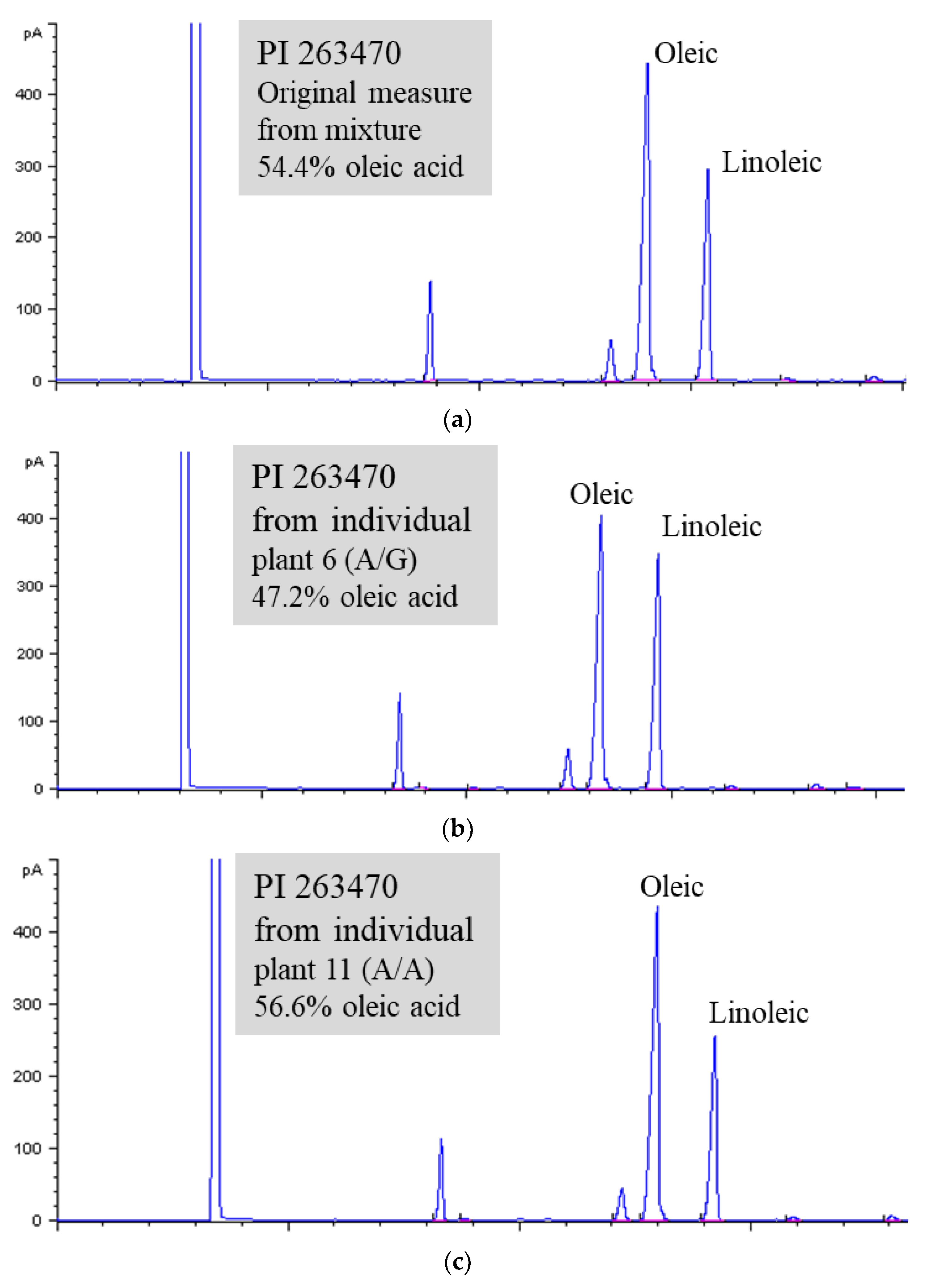
One accession (PI 263470) containing a significantly higher level of oleic acid (54.0%) than the average (39.4%) (unpublished data, Wang et al.) was identified from the USDA sesame germplasm collection by gas chromatography (GC) analysis (Figure 1A). From the germplasm passport record, this accession originated in the former Soviet Union and was donated to the collection in 1960 by Toyama University, Japan. The high oleate trait was followed up by further GC analysis and DNA sequence analysis using fresh seeds and leaf tissue collected from the plants regrown in the greenhouse.
2.1.2. Confirmation of Naturally High Oleate Mutants by DNA Sequence Analysis
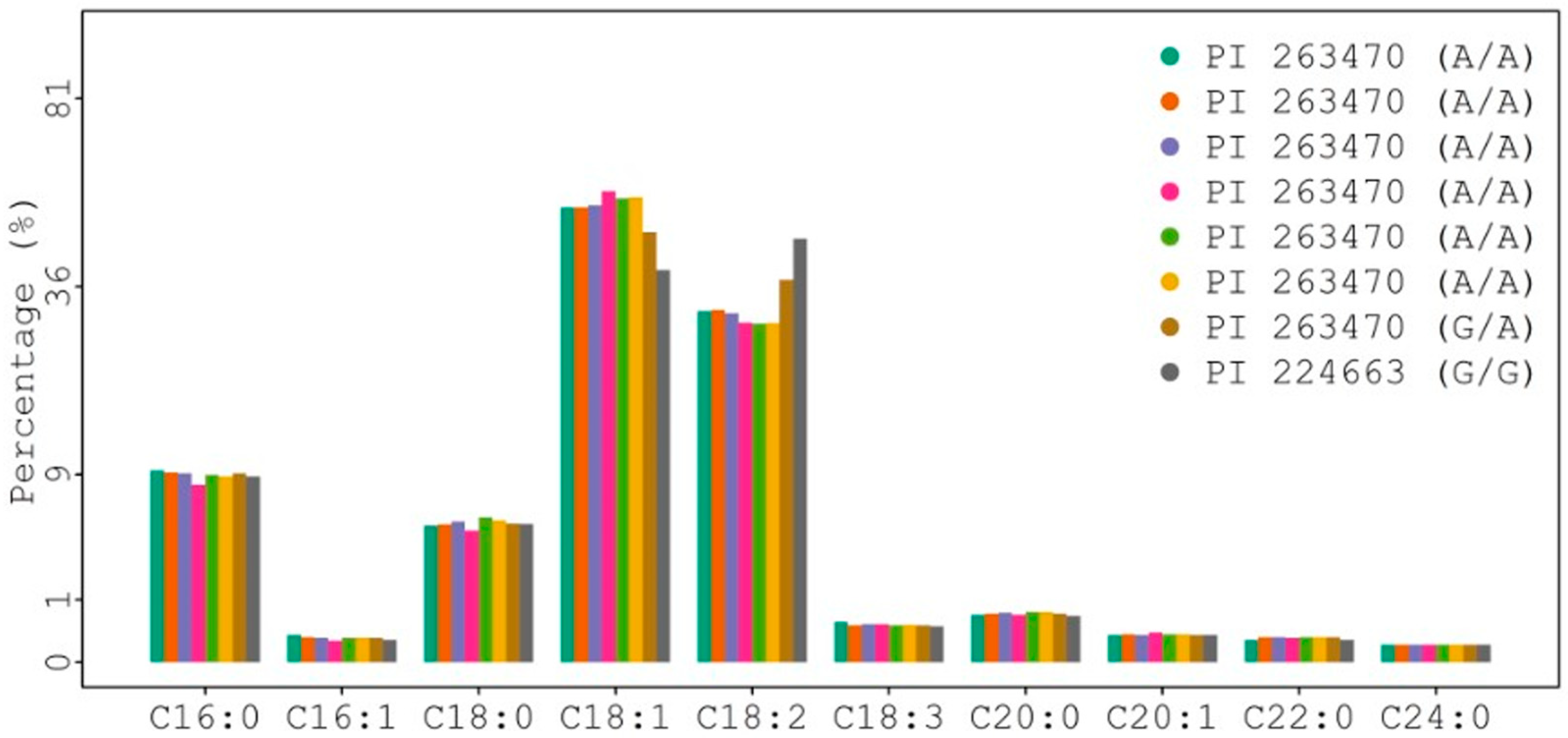
The FAD2 gene encodes a fatty acid desaturase (FAD) that is responsible for the desaturation ratio of oleic acid to linoleic acid (O/L) in oilseed crops. To check and compare the coding sequence of the FAD2 gene among individual plants of PI 263470 plus PI 224663 (as a control for the average level of oleic acid), leaf tissues were collected from individual plants. The results from DNA sequence analysis of multiple plants indicated that position 425 in the coding sequence had A/A, G/A, and G/G genotypes (Figure 2 and Table S1). Thus, DNA sequences from individual plants confirmed that PI 263470 was a mixed accession.
The results from a comparison of FAD2 gene sequences among eight sesame accessions and partially deduced amino acid sequences of FAD2 among sesame, soybean, and peanut are shown in Figure S1. Three histidine (H)-boxes (critical sites for fatty acid desaturase displaying its function) were identified, but only the flanking sequences around H-Box2 is shown in Figure S1B. The DNA mutation (G425A) in PI 263470 (Figure S1A) occurred in H-Box2, which led to an amino acid substitution of R142H (arginine substituted to histidine) within the fatty acid desaturase. This amino acid substitution likely explains the higher oleate content in seeds of PI 263470 relative to other sesame accessions. Therefore, this functional mutation can presumably reduce fatty acid desaturase activity and result in an increased level of oleic acid and the decreased level of linoleic acid in sesame seeds.
2.1.3. Reconfirmation of Naturally High Oleate Mutants by GC Analysis
Additional seeds from eight individually sequenced plants were collected and used for GC analysis. The results from GC analysis are shown in Figure 2 and Table S1. The heterozygous genotype of plant 3-6 (A/G) and homozygous genotype of plant 3-11 (A/A) from PI 263470 had 47.2 and 56.6% oleic acid, respectively. They had a higher level of oleic acid than the homozygous genotype of plant 2 (G/G) from PI 224663 with 39.22% oleic acid. All other plants (3-1, 3-3, 3-10, 3-22, and 3-23) with the A/A genotype from PI 263470 also produced seeds with >52% oleic acid. The results from both sequence analysis and GC analysis suggest that the natural mutation of G425A in the coding region of the FAD2 gene may be responsible for the enhanced level of oleic acid from 39.2 to 56.6%.
2.2. EMS Mutagenesis and Generation of Mutant Population
2.2.1. EMS Mutagenesis
In other oilseed species, the oleate level can reach to about 80%. The natural mutant sesame accession we identified was only about 54–56%. This accession would be a good starting genetic material for mutagenesis to further enhance the level of oleate.
The germination rate and seed setting can be greatly affected by EMS treatment dosage. To determine the EMS concentration, four dosages were used (phosphate buffer only, 1.0%, 1.5%, and 2.0%). We found after EMS treatment, the seed germination rate (95.31%) with 1.0% EMS treatment was very close to the control (no EMS) germination rate (95.75%). After further reducing EMS concentration, we concluded that 0.8% EMS concentration may be the most suitable treatment for sesame seed mutagenesis for our lab protocol.
2.2.2. Generation of Mutant Populations
In 2014, 1800 sesame seeds were treated with 0.8% EMS. The procedure of mutagenesis from M1 seeds to M3 seeds is shown in Figure 3. After planting in the greenhouse, 1553 M1 seeds germinated for a rate of 86.3%. Of these, some seedlings died, and some mature plants did not produce seeds. In the end, we only harvested seeds from 1412 M1 plants for a seed setting rate of 90.9%. In 2015, one or two seeds from each M2 line (depending on the seeds available for each line) were planted to clay pots. After planting, 1030 M2 seeds were germinated. The average germination rate of M2 seeds was 72.95%. After germination, some seedlings either died or the mature plants did not produce seeds despite flowering. Only 329 M2 plants produced M3 seeds. The M2 plants seed setting rate (31.94%) was probably reduced because more recessive lethal alleles were revealed or exposed in M2 from M1 selfing. M2 plant morphology was observed in the greenhouse. M3 seeds were harvested and used for chemical analysis, genetic analysis, and planted for advancement to the next generation.
2.3. Characterization and Evaluation of Mutants
2.3.1. Morphological Mutants
Although 1030 M2 seeds germinated in the greenhouse, some M2 seedlings showed albinism and some M2 seedlings did not develop into mature plants. In total, 405 M2 seedlings were dead before developing into mature plants. Among 625 M2 plants, some did not flower, while others flowered but did not produce seeds. Among M2 plants, clear variation from two agronomic traits (plant height and maturity) was observed. After GC analysis, seeds from some M3 lines were planted in one-gallon pots for morphological observation (Figure 4). Compared with the control (PI 263470), the M3 200-1-11 plant had flat stems with darker leaves and M3 447-1-1 was leafy (more leaves clustering at the top of plant). Morphological mutations with leafy and clustered capsules were also observed in a previous study [15]. Since identification of high oleate mutants was our focus in this study, only a few M3 mutant families were planted for further morphological observations in the early generations.
2.3.2. DNA Sequence Analysis of High Oleate Mutants
Using GC analysis of M3 seeds from the M2 plants, we identified five high oleate mutants (M3 965-2-36, M3 965-2-33, M3 965-2-11, M3 915-1-35, and M3 200-1-11). Their oleic acid level was approximately 70%. Leaf tissue was collected from M3 plants, and DNA was extracted from these leaves. These five high oleate mutants were sequenced in the coding region of the FAD2 gene. Surprisingly, there were no mutations identified in the coding region when compared with the control (PI 263470 with A/A). We also attempted to sequence the promoter region of this gene but encountered repetitive sequence issues and could not get high quality sequence reads to assemble the complete sequence of the promoter region. DNA sequencing from more genome regions is required.
Two possible explanations may account for the increase in oleic acid content in mutants. First, the causal mutations are in the promoter region, but we have not identified any yet. Future experiments comparing the gene expression level of FAD2 between the mutant and control may help to determine whether the regulatory region contributes to the enhanced level of oleic acid. Alternatively, other genes such as ROD1 (reduced oleate desaturation 1) gene identified in Arabidopsis [17], but which has not been identified in sesame, may partially control oleic acid levels. These genes need to be identified and sequenced for comparison with the control line.
2.3.3. High Generation of Oleate Mutant Lines
At the M3 generation, all identified lines were still segregating for fatty acid level and other plant morphological traits. We advanced these mutant lines in the greenhouse while monitoring the level of oleic acid as well as conducting some minor selection on plant morphology and maturity. In 2022, six high oleate mutant lines plus the control were planted (28 April 2022) and grown in the greenhouse. Their planting, flowering, and harvest dates are listed in Table S2. Compared with the control (PI263470 A/A), five mutants flowered at similar times (around 20 May), but the mutant M7 965_2_33_7_6_10 had small plant stature at the early stage (about one month old) and flowered on 06/28, which was over a month later than the control (Figure 5). Two mutants (M7 915_1_35_4_3_2 and M7 915_1_35_5_5_3) were harvested three and five days earlier than the control, respectively (07/18). Four mutants (M7 965_2_36_5_3_36, M7 965_2_11_6_2_29, M8 200_1_11_15_3_5_21, and M7 965_2_33_7_6_10) were harvested six days, eight days, ten days, and one month later than the control, respectively.
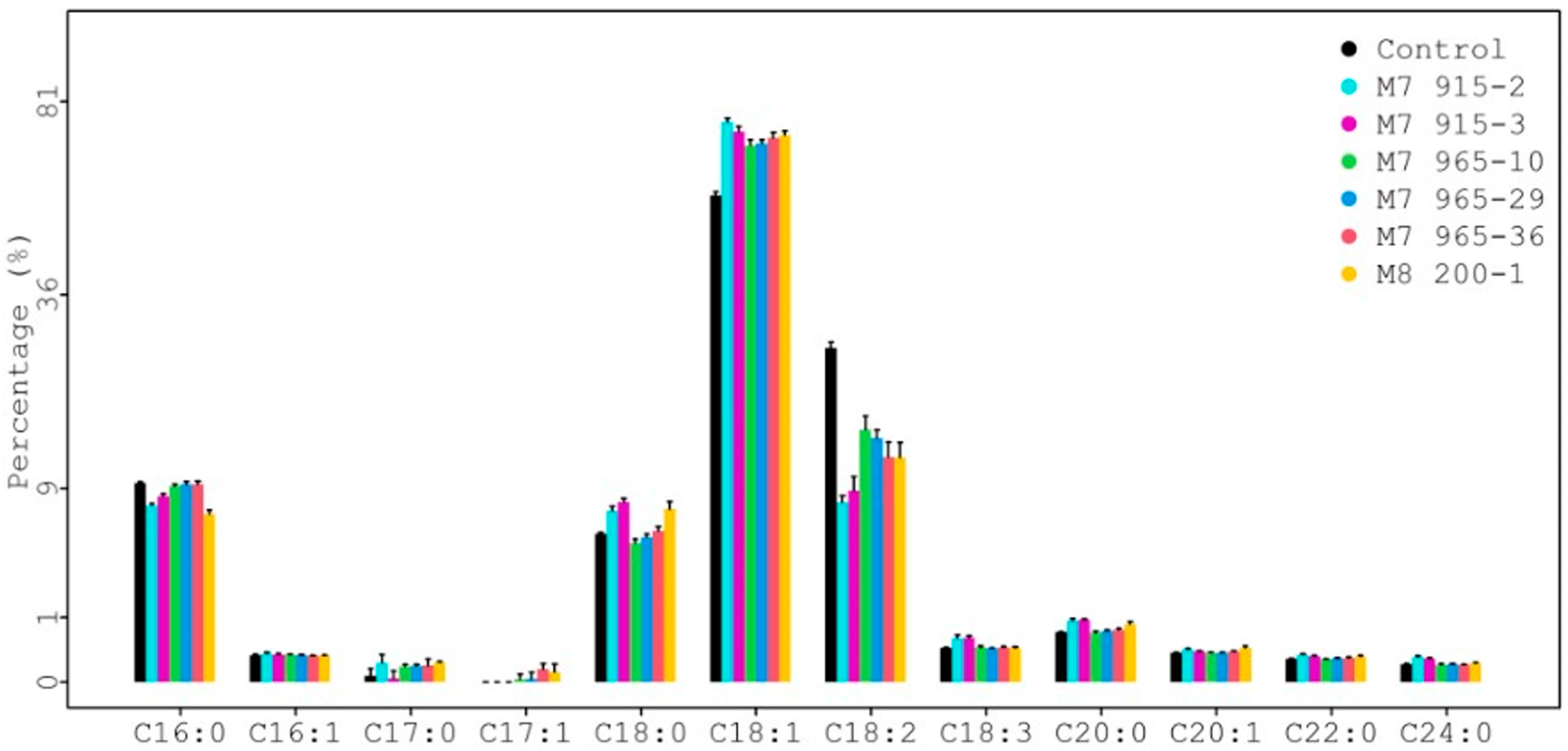
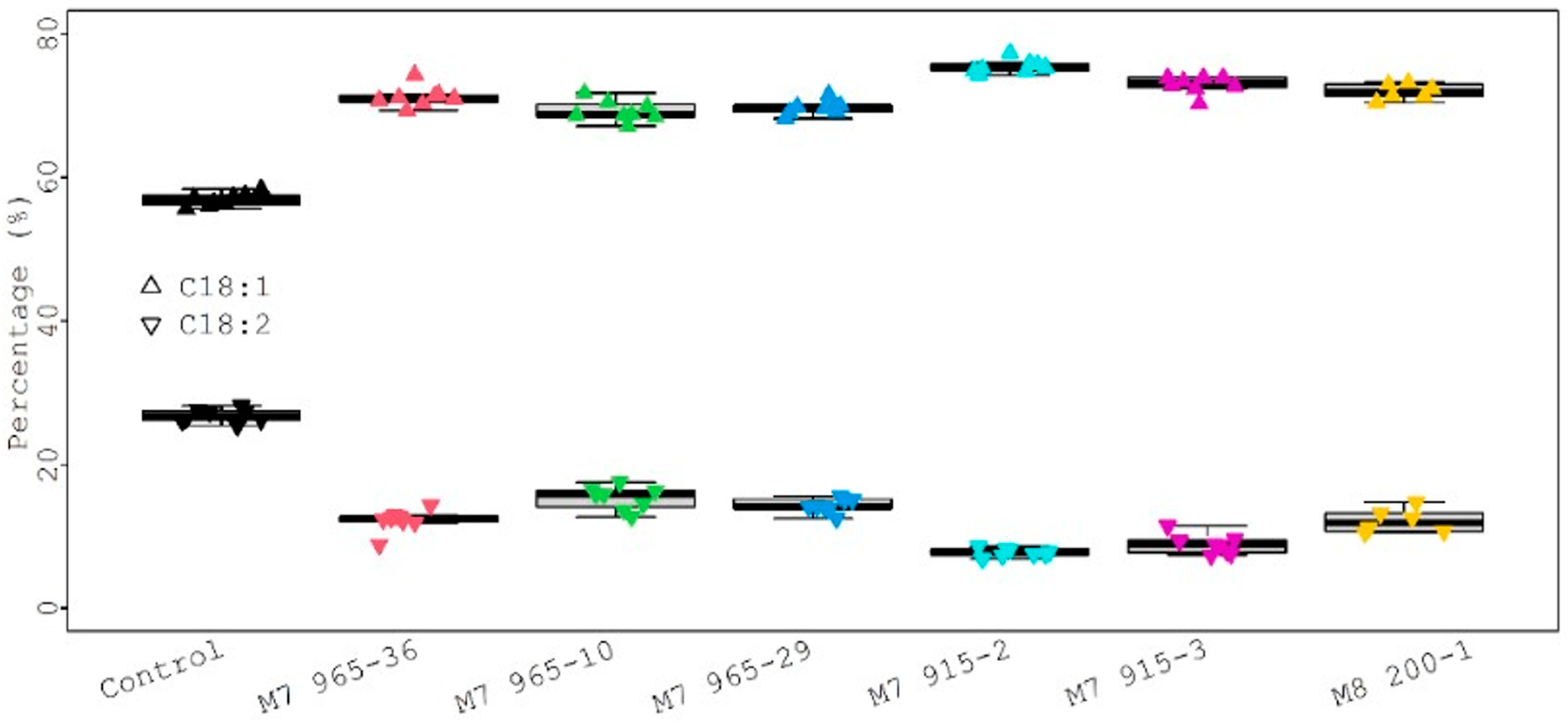
The seed fatty acid composition results from each mutant line plus the control are summarized in Table S3 and shown in Figure 6. There were 12 fatty acids identified. Among them, four major fatty acids (>1%) were palmitic (C16:0), stearic (C18:0), oleic (C18:1), and linoleic (C18:2) [18,19]. The rest were minor fatty acids (<1%). For each fatty acid, the average value from eight individual plants is in the cell, and the range is in the bracket for each genotype in Table S3. For example, the average value and range of stearic acid (C18:0) for the control (PI 263470) is 5.28% and 5.09–5.44%, respectively.
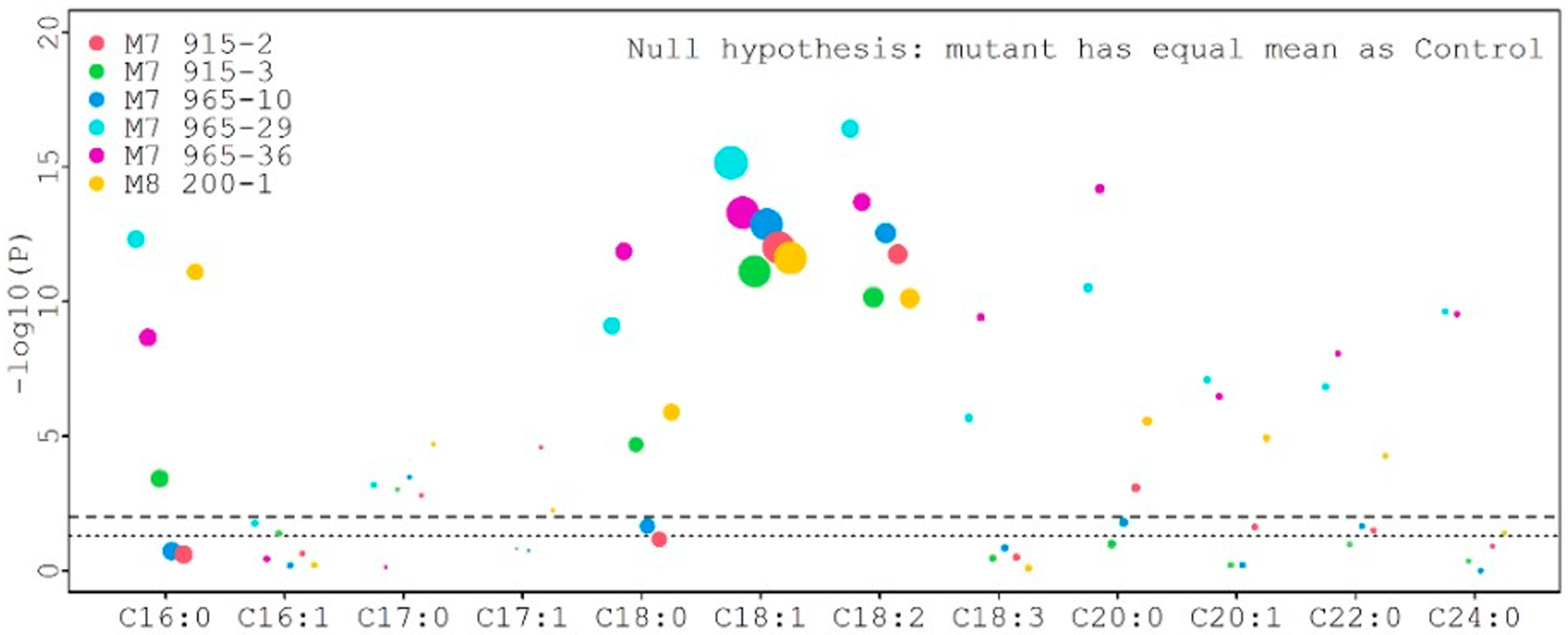
For statistical analysis, each fatty acid value from all mutants was compared with the control value, and the results are presented in Figure 7. Six different color dots represent six mutant lines. The size of different dots represents the different percentage of fatty acids. Larger dots correspond to higher fatty acid percentages. The lower and upper dotted lines represent the threshold value for p = 0.05 and p = 0.01, respectively. The palmitic acid (C16:0) levels (6.80%, 7.48%, 8.32%, and 9.24%) from four mutants (M8 200-1, M7 915-2, M7 915-3, and M7 965-10) were significantly lower than the control (9.54%) at p = 0.01, but the palmitic acid level (9.40% and 9.41%) from two mutants (M7 965-29 and M7 965-36) was not significantly different from the control at p = 0.05. The stearic acid (C18:0) levels (7.20%, 7.08%, and 7.81%) from three mutants (M8 200-1, M7 915-2, and M7 915-3) were significantly higher than the control (5.28%) at p = 0.01. The stearic acid levels (4.65% and 5.05%) from M7 965-10 and M7 965-29 were significantly lower than the control (5.28%) at p = 0.01 and p = 0.05, respectively. The stearic acid level (5.50%) from mutant M7 965-36 was not significantly different from the control. The oleic acid (C18:1) and linoleic acid (C18:2) levels from all mutants were significantly higher and lower than the control at p = 0.01, respectively (Table S3 and Figure 7). For the four major fatty acids, in general, stearic acid significantly increased and palmitic acid significantly decreased after mutagenesis. Additionally, oleic acid significantly increased and linoleic acid significantly decreased. Compared with the control, this large negative change between oleic acid and linoleic acid can be clearly observed from Figure 8. We also observed that the variation among eight individual plants within the same mutant was not significant.
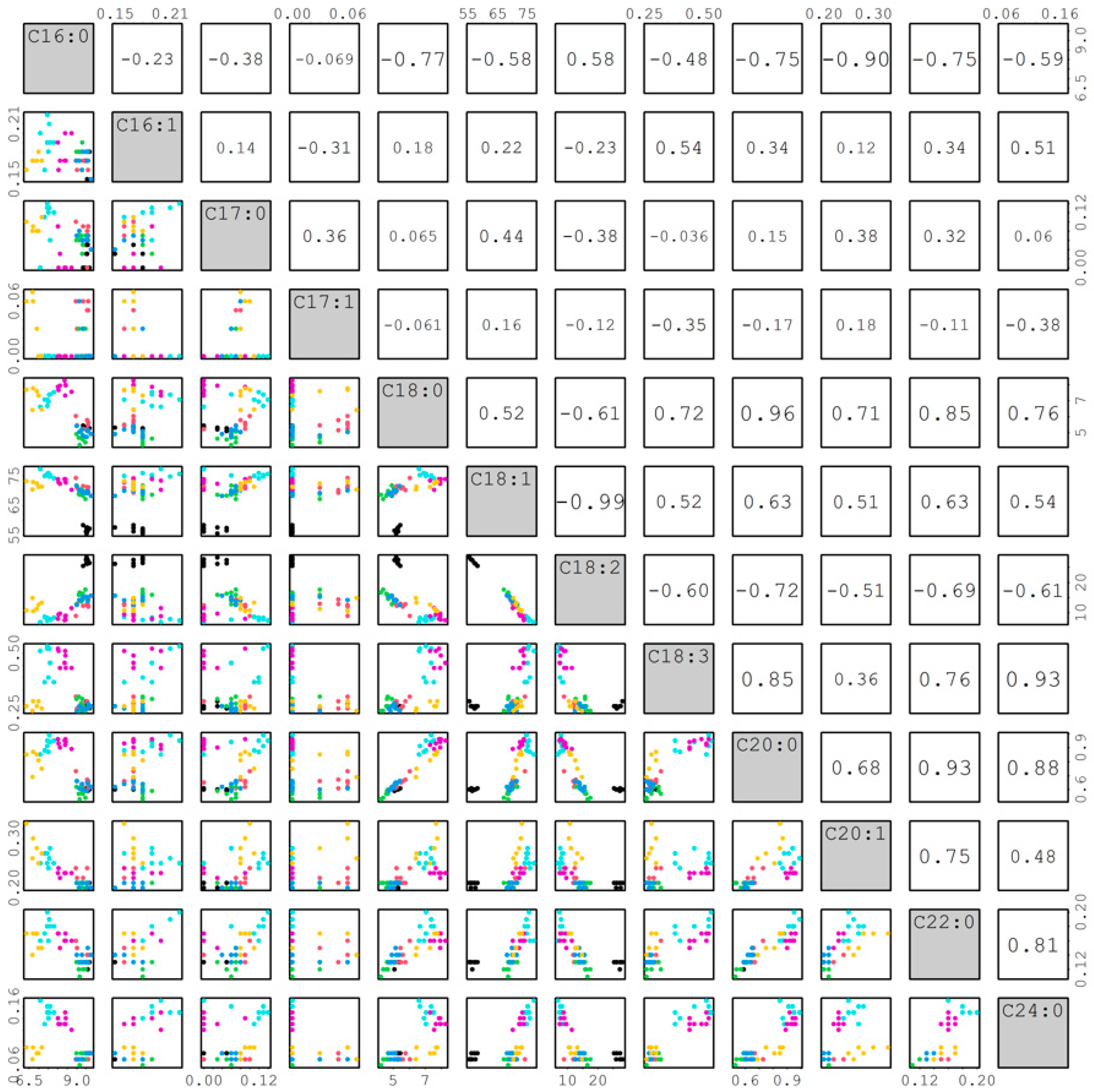
2.3.4. Correlation among Fatty Acids
The correlations among 12 fatty acids are shown in Figure 9. The r value can reflect how the traits change relative to one another. Most r values from the correlation of palmitoleic (C16:1), margaric (C17:0), and heptadecenoic (C17:1) with other fatty acids were lower than 0.5, whereas most r values among the other nine fatty acids were larger than 0.5. Therefore, we focused on discussing the correlations among these nine fatty acids from the top to bottom of Figure 9. Palmitic acid (C16:0) had significantly negative correlations (r = −0.77, −0.58, −0.48, −0.75, −0.90, −0.75, and −0.59) with stearic (C18:0), oleic (C18:1), linolenic (C18:3), arachidic (C20:0), eicosenoic (C20:1), behenic (C22:0), and lignoceric (C24:0) except for a significantly positive correlation (r = 0.58) with linoleic (C18:2) acid. Stearic acid had significantly positive correlations (r =0.52, 0.72, 0.96, 0.71, 0.85, and 0.76) with oleic, linolenic, arachidic, eicosenoic, behenic, and lignoceric except for a significantly negative correlation (r = −0.61) with linoleic. Oleic acid had significantly positive correlations (r = 0.52, 0.63, 0.51, 0.63, and 0.54) with linolenic, arachidic, eicosenoic, behenic, and lignoceric except for significantly and negatively correlated (r = −0.99) with linoleic acid. Linoleic acid had significantly negative correlations (r = −0.6, −0.72, −0.51, −0.69, and −0.61) with linolenic, arachidic, eicosenoic, behenic, and lignoceric. Arachidic acid had significantly positive correlations (r = 0.68, 0.93, and 0.88) with eicosenoic, behenic, and lignoceric. Eicosenoic acid had significantly positive correlations (r = 0.75 and 0.48) with behenic and lignoceric acids. Behenic acid also had a significantly positive correlation (r = 0.81) with lignoceric acid. The correlations among fatty acids identified above will be useful for sesame breeders to improve seed nutritional qualities in breeding programs. Furthermore, the FAD2 genes showed a high effect for oleic acid and linoleic acid (or O/L ratio) in peanuts [20]. The trait for O/L ratio in sesame may also be highly heritable. Therefore, the high oleate trait can be easily introduced from the mutant lines into elite lines by backcrossing in sesame.
3. Materials and Methods
3.1. Plant Materials and Plant Growth under Greenhouse Conditions
Sesame seeds were planted in the greenhouse under controlled conditions (26.7 °C/8 h for day with natural light and 32.2 °C/16 h with natural dark). Due to the space limitation, we adjusted water content and fertilizer amount from planting to harvesting for controlling the plant height and size. The purpose was to harvest enough seeds from each plant for one generation. To prevent plants from lodging, each plant was supported by a bamboo pole tied with a piece of white plastic tape. The seeds were harvested from each capsule and put into small paper bags which were transferred to the seed storage room for drying (21 °C, 25% RH for two weeks). The dried seeds were used for next generation planting or fatty acid analysis.
3.2. EMS Mutagenesis
The accession (PI 263470) with high oleate (54%) was used for mutagenesis. On 3 March 2014, about 600 purified seeds harvested from the plant number 6-3-6 (with genotype A/A) were put into a 50 mL tube containing 40 mL of 100 mM phosphate buffer (pH 7.5) and were incubated at 4 °C for 24 h to allow seed imbibition. In total, we treated the seeds in four 50 mL tubes (4 tubes × 600 seeds = 2400 seeds). After imbibition, the buffer was removed. Freshly made 40 mL of 0.8% EMS in 100 mM phosphate buffer was added to the tube. The tube was covered with aluminum foil for preventing light followed by agitation on a shaker at 80 rpm at room temperature for 8 h. After EMS treatment, the EMS solution was removed into a waste container. The treated seeds were washed with tap water (with 3–4 changes during 30 min).
The treated (M1) seeds were planted into a tray filled with soil (Metro-Mix360, Product of Canada, Sun Gro Horticulture, Vassar, MB, Canada) and placed in the greenhouse. Each tray had 32 small pots, and two seeds were planted into each pot. After planting, the tray was covered with a transparent plastic cover to maintain moisture levels. After seed germination, the transparent plastic cover was removed immediately. Once the seedlings were well established (with two true leaves), they were thinned to one seedling per pot. This thinning process prevented us from calculating the mutation rate because we preferred to remain only one healthy plant per pot. When the plants grew taller, a thin bamboo pole was inserted into each pot and tied to the plant using a piece of white plastic tape to prevent plants from lodging (see M1 young seedlings and plants in Figure 3). On 21 April 2014, most plants flowered. To prevent plants from snapping, we watered the plants from both sides only. On 30 May 2014, some capsules turned brown and matured. At this time, we started harvesting mature capsules. We finished harvesting all the mature capsules by 7 July 2014. After drying, we found some capsules containing mature seeds and some capsules did not contain good seeds (immature or seeds with poor quality). After seed cleaning, 1412 lines of M2 seeds were harvested from the M1 plants.
On 11 August 2014, all M2 seeds plus the control (10 seeds from PI 263470 that were not treated with EMS) were planted into small clay pots (1–2 seeds planted in each pot) in the greenhouse. The M2 plant morphology was observed and recorded after 48 days of growth. Not all plants produced M3 seeds. M3 seeds were harvested from 329 M2 plants, and they were used for fatty acid composition analysis by GC.
Fatty acid analysis of M3 seeds showed five mutant lines with high oleate mutants (among them, one mutant line separated into two mutant lines due to significant segregation), and they were planted in the greenhouse and advanced to later generations. On 18 April 2022, six mutant lines (plus one control) were planted in the greenhouse. Plant morphology was observed and recorded. Seeds were harvested from eight plants for each mutant line and these seeds from individual plant were used for GC analysis.
3.3. Fatty Acid Composition Analysis with GC
Since M3 seeds were still segregating, individual M3 seeds were selected and analyzed for fatty acid composition using gas chromatography (GC). Two seeds were measured for two replicates. The average from two measurements represented the fatty acid composition value of each M3 line. In total, 635 M3 seeds were analyzed. A single seed was put into a 1.5 mL glass injection vial. A small glass rod was used for manually crushing the seed into a fine powder within the vial. Fatty acid methyl esters (FAMEs) were prepared from sesame seed powder by alkaline transmethylation. Fatty acid composition was determined on an Agilent 7890A gas chromatograph (Santa Clara, CA, USA) equipped with a flame ionization detector (FID) following a previously described lab method [21].
3.4. DNA Sequence and Deduced Amino Acid Sequence Analysis
Fresh leaf tissue (75–100 mg) was collected from individual sesame plants. DNA was extracted from these fresh leaf tissues using an Omega Bio-Tek E.Z.N.A. (Plant DNA kit, Norcross, GA, USA). DNA quality and quantity were determined on a Nanodrop 2000C spectrophotometer (Themo Scientific, Wilmington, DE, USA). DNA quality of samples was also checked by using 1% agarose gel electrophoresis. DNA concentration for all samples was diluted to 10 ng/µL. Diluted DNA was used as the template for amplification of the coding region of the FAD2 gene. The primers (5′ TGGGAGGTTTTGATTCAGACA 3′ and 5′ CACTAACAAAGGACGCATCTTA 3′) from the flanking coding region of sesame FAD2 were designed (modified from Chen et al., 2014) and synthesized by Operon (Qiagen, Germantown, MD, USA, http://www.operon.com, accessed on 18 November 2022). The PCR mix contained 2 mM of MgCl2, 0.5 mM of dNTPs, 0.25 µM each of primer, and 15 ng of DNA template for a total volume of 20 µL. The PCR cycling conditions consisted of 1 cycle of 94 °C for 5 min, 35 cycles of 94 °C for 30 s, 72 °C for 100 s, and a final extension of 72 °C for 7 min. Amplicons were sequenced using PCR primers and two internal primers (5′ GAGGCCAACCGAGAGTAAGA 3′ and 5′ CTGCTCGTGCCCTATTTCTC 3′) to cover the coding region. After trimming each sequence, consensus sequences were assembled using Sequencher, ver. 5.3. SNPs were identified from the natural mutant but not from EMS-induced mutants. Substitutions were identified in polypeptides by comparison of the deduced amino acid sequences.
3.5. High Generation of Mutant Lines
Five M3 mutant lines (M200-1-11, M915-1-35, M965-2-11, M965-2-33, and M965-2-36 plus the control were purified PI 263470 with A/A), further selected, and followed up to the M7 or M8 generations in 2022. Their M6 or M7 plant morphology was observed in the green house, and the oleate level from M7 or M8 seeds was measured by GC using 7–10 seeds in a bulk analysis.
3.6. Data Analysis
For the six mutant lines and control, the mean value from two replications (when available) of each fatty acid composition was analyzed and visualized with R v4.1.1. For each mutant, varTest in R was used to test whether the mutant had significantly different variance than the control. Then a student’s t-test was conducted to test whether the composition was significantly different between the mutant line and the control. Pairwise correlations among fatty acid composition were plotted by the pair R function.
4. Conclusions
We identified a natural high oleate accession (PI 263470) that harbors a nonsynonymous mutation (R142H) in the FAD2 gene by screening the entire USDA sesame germplasm collection. This accession has 54% oleic acid compared with the average of 39.4% of the entire collection. Functional SNP markers were developed from this natural mutant for the marker-assisted selection (MAS). Because this high level of oleic acid in sesame is still lower than 80% achieved by high oleic varieties in other oilseed crops, we employed EMS-induced mutagenesis of this natural accession to further increase the oleic acid level. Several mutants that had oleic acid over 70% were isolated. However, no additional mutation was identified in the coding region of the FAD2 gene. More genome regions (including regulatory regions and other possible copies of FAD2 genes) should be sequenced. Crosses among these mutants can be made to combine different mutant loci if they are not linked to further enhance the oleic acid level. After reaching 80%, the new high oleate genetic lines can be crossed with the elite breeding lines to develop high oleate sesame cultivars.
Supplementary Materials
The following supporting information can be downloaded at: https://0-www-mdpi-com.brum.beds.ac.uk/article/10.3390/plants12061294/s1, Figure S1A: Alignment of FAD2 gene DAN sequences from eight sesame accessions; Figure S1B: Identification of an amino acid substitution of R142H in the H-Box2 by comparison of the partial deduced amino acids sequences from FAD2 genes among species from three different genera. Table S1: Comparison of fatty acid profiles among different FAD2 genotypes; Table S2: Planting time, early plant stature, flowering time, and harvest time in 2022 in greenhouse; Table S3: Average value and range each fatty acid of sesame mutant lines.
Author Contributions
M.L.W. conceived this project and wrote the first draft of this manuscript. B.T. performed chemical analysis, analyzed data, and helped revise this manuscript. X.L. performed statistical analysis and revised this manuscript. J.B.M. provided germplasm accessions, guided planting, harvesting sesame in the greenhouse, and revised this manuscript. All authors have read and agreed to the published version of the manuscript.
Funding
This research received no external funding.
Data Availability Statement
The data and plant materials that support the findings of this study are available from the corresponding author upon request.
Acknowledgments
We are grateful to David Pinnow for helping manage the sesame plants in the greenhouse and cleaning sesame seeds during the project.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Namiki, M. Nutraceutical Functions of Sesame: A Review. Crit. Rev. Food Sci. Nutr. 2007, 47, 651–673. [Google Scholar] [CrossRef]
- Pathak, N.; Rai, A.K.; Kumari, R.; Bhat, K.V. Value addition in sesame: A perspective on bioactive components for enhancing utility and profitability. Phacog. Rev. 2014, 8, 147–155. [Google Scholar] [CrossRef] [Green Version]
- Morris, J.B.; Wang, M.L.; Tonnis, B.D. Variability for oil, protein, lignan, tocopherol, and fatty acid concentrations in eight sesame (Sesamum indicum L.) genotypes. Ind. Crop. Prod. 2021, 164, 1–12. [Google Scholar] [CrossRef]
- Nayar, N.M.; Mehra, K.L. Sesame: Its use, botany, cytogenetics, and origin. Econ. Bot. 1970, 24, 20–31. [Google Scholar]
- Morinaga, T.; Fukushima, E.; Kano, T.; Yamasaki, Y. Chromosome Numbers of cultivated Plants II. J. Plant Res. 1929, 43, 589–594. [Google Scholar] [CrossRef] [Green Version]
- Zhang, H.; Miao, H.; Wang, L.; Qu, L.; Liu, H.; Wang, Q.; Yue, M. Genome sequencing of the important oilseed crop Sesamum indicum L. Genome Biol. 2013, 14, 401. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, L.; Yu, S.; Tong, C.; Zhao, Y.; Liu, Y.; Song, C.; Zhang, Y.; Zhang, X.; Wang, Y.; Hua, W.; et al. Genome sequencing of the high oil crop sesame provides insight into oil biosynthesis. Genome Biol. 2014, 15, R39. [Google Scholar] [CrossRef] [Green Version]
- Wei, X.; Zhu, X.; Yu, J.; Wang, L.; Zhang, Y.; Li, D.; Zhou, R.; Zhang, X. Identification of Sesame Genomic Variations from Genome Comparison of Landrace and Variety. Front. Plant Sci. 2016, 7, 1169. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wei, X.; Gong, H.; Yu, J.; Liu, P.; Wang, L.; Zhang, Y.; Zhang, X. SesameFG: An integrated database for the functional genomics of sesame. Sci. Rep. 2017, 7, 1–10. [Google Scholar] [CrossRef] [Green Version]
- You, J.; Li, D.; Dossou, S.S.K.; Zhou, R.; Zhang, Y.; Wang, L. CRISPR/Cas9-Mediated Efficient Targeted Mutagenesis in Sesame (Sesamum indicum L.). Front. Plant Sci. 2022, 13, 935825. [Google Scholar] [CrossRef]
- Norden, A.J.; Borget, D.W.; Knauft, D.A.; Young, C.T. Variability in Oil Quality Among Peanut Genotypes in the Florida Breeding Program1. Peanut Sci. 1987, 4, 7–11. [Google Scholar] [CrossRef]
- Hu, X.; Sullivan-Gilbert, M.; Gupta, M.; Thompson, S.A. Mapping of the loci controlling oleic and linolenic acid contents and development of fad2 and fad3 allele-specific markers in canola (Brassica napus L.). Theor. Appl. Genet. 2006, 113, 497–507. [Google Scholar] [CrossRef] [PubMed]
- Pham, A.; Lee, J.; Shannon, J.G.; Bilyeu, K. Mutant alleles of FAD2-1A and FAD2-1B combine to produce soybeans with the high oleic acid seed oil trait. BMC Plant Biol. 2010, 10, 195. [Google Scholar]
- Anjani, K.; Yadav, P. High yielding-high oleic non-genetically modified Indian safflower cultivars. Ind. Crop. Prod. 2017, 104, 7–12. [Google Scholar] [CrossRef]
- Ҫağirgan, M.İ. Selection and morphological characterization of induced determinate mutants in sesame. Field Crops Res. 2006, 96, 19–24. [Google Scholar]
- Uzun, B.; Ҫağirgan, M.İ. Comparison of determinate and indeterminate lines of sesame for agronomic traits. Field Crops Res. 2006, 96, 13–18. [Google Scholar]
- Lu, C.; Xin, Z.; Ren, Z.; Miquel, M.; Browse, J. An enzyme regulating triacylglycerol composition is encoded by the ROD1 gene of Arabidopsis. Proc. Natl. Acad. Sci. USA 2009, 106, 18837–18842. [Google Scholar] [CrossRef] [Green Version]
- Kamal-Eldin, A.; Yousif, G.; Iskander, G.M.; Appelqvist, L.A. Seed lipid of Sesame indicum L. and related wild species in Sudan I: Fatty acids and triacylglycerols. Fette Seifen Anstrichm. 1992, 94, 254–259. [Google Scholar] [CrossRef]
- Chen, C.; Tonnis, B.; Morris, B.; Wang, B.R.; Zhang, A.L.; Pinnow, D.; Wang, M.L. Variation in Seed Fatty Acid Composition and Sequence Divergence in the FAD2 Gene Coding Region between Wild and Cultivated Sesame. J. Agric. Food Chem. 2014, 62, 11706–11710. [Google Scholar] [CrossRef]
- Pandey, M.K.; Wang, M.L.; Qiao, L.; Feng, S.; Khera, R.; Wang, H.; Tonnis, B.; Barkley, N.A.; Wang, J.; Holbrook, C.C.; et al. Identification of QTLs associated with oil content and mapping FAD2 genes and their relative contribution to oil quality in peanut (Arachis hypogaea L.). BMC Genet. 2014, 15, 1–14. [Google Scholar] [CrossRef] [Green Version]
- Wang, M.L.; Chen, C.Y.; Tonnis, B.; Barkley, N.A.; Pinnow, D.L.; Pittman, R.N.; Davis, J.; Holbrook, C.C.; Stalker, H.T.; Ped-erson, G.A. Oil, fatty acid, flavonoid, and resveratrol content variability and FAD2A functional SNP genotypes in the U.S. peanut mini-core collection. J. Agric. Food Chem. 2013, 61, 2875–2882. [Google Scholar] [PubMed]
Figure 1.
The levels of oleic acid and linoleic acid shown on chromatograms from three genotypes. (A). Mixed genotypes (A/A and A/G). (B). Genotype A/G. (C). Genotype A/A.
Figure 1.
The levels of oleic acid and linoleic acid shown on chromatograms from three genotypes. (A). Mixed genotypes (A/A and A/G). (B). Genotype A/G. (C). Genotype A/A.

Figure 2.
Comparison of fatty acid profiles among eight plants with FAD2 genotypes.

Figure 3.
The procedure of mutagenesis from M1 seeds to M3 seeds.

Figure 4.
Comparison of plant morphology between mutant line and control.

Figure 5.
Comparison of plant morphology between mutant line and control at a high generations.

Figure 6.
Comparison of fatty acid profiles among mutants at higher generations.

Figure 7.
Comparison of each fatty acid value from all mutants with the control.

Figure 8.
Comparison of oleic and linoleic between control and mutants.

Figure 9.
Correlations among different fatty acids in sesame.

Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Wang, M.L.; Tonnis, B.; Li, X.; Morris, J.B. Generation of Sesame Mutant Population by Mutagenesis and Identification of High Oleate Mutants by GC Analysis. Plants 2023, 12, 1294. https://0-doi-org.brum.beds.ac.uk/10.3390/plants12061294
AMA Style
Wang ML, Tonnis B, Li X, Morris JB. Generation of Sesame Mutant Population by Mutagenesis and Identification of High Oleate Mutants by GC Analysis. Plants. 2023; 12(6):1294. https://0-doi-org.brum.beds.ac.uk/10.3390/plants12061294
Chicago/Turabian StyleWang, Ming Li, Brandon Tonnis, Xianran Li, and John Bradly Morris. 2023. "Generation of Sesame Mutant Population by Mutagenesis and Identification of High Oleate Mutants by GC Analysis" Plants 12, no. 6: 1294. https://0-doi-org.brum.beds.ac.uk/10.3390/plants12061294
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.