
Antifungal Activity and Effect of Plant-Associated Bacteria on Phenolic Synthesis of Quercus robur L.

 , , , , ,
, , , , ,  and
and 
Abstract
:1. Introduction

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Region | Species | Ecological Niche | Authors |
|---|---|---|---|
| Central, Western, Southern, and Northern Europe | Q. petraea | endophytes of leaves, shoots and branches of different ages | Halmschlager et al., 1993, Fort et al., 2021 [17] |
| Q. robur | Griffith et Boddy, 1990, Petrini et Fisher, 1990, Kowalski et Kehr 1996, Gennaro et al., 2003, Ragazzi et al., 2003, Gonthier et al., 2006, Agostinelli et al., 2018, Matule, 2018 [54,55,56,57,58,59] | ||
| Quercus ilex | the leaves and twigs | Fisher et al., 1994, Collado et al., 1996, 1999 [60,61] | |
| Q. faginea | the leaves and twigs | Collado et al., 1996, 1999 [60] | |
| Q. cerris, Q. pubescens | the leaves and twigs | Ragazzi et al., 2001, 2003, Gennaro et al., 2003, Moricca et al., 2012 [59,62] | |
| Q. suber | young and old twigs, branches, and woody tissues | Linaldeddu et al., 2011, Costa et al., 2019 [63,64] | |
| Asia | Q. macranthera, Q. brantii | twigs, branches | Ghasemi-Esfahlan et al., 2019 [65] |
2. Results and Discussion
2.1. Determination of Pectolytic Enzymes Activity
2.2. 16S rRNA Gene Sequencing
2.3. Antifungal Activity of Plant-Associated Bacteria
2.4. Reactions of Oak Seedlings to the Inoculation of Endophytic, Epiphytic and Phytopathogenic Bacteria
3. Materials and Methods
3.1. Study Area
3.2. Isolation, Cultivation and Studying of the Biological Activity of Endophytic Bacteria
3.2.1. Isolation and Cultivation of Bacteria
3.2.2. Pectolytic Activity Tests
3.2.3. Determination of Antifungal Activity
3.3. Molecular Genetic Identification of Isolates
3.4. Biochemical Tests
3.4.1. Methods of Sample Collection
3.4.2. Determination of the Total Phenolic Content in Leaves
3.4.3. Determination of the Total Flavonoid Content
3.4.4. Determination of Antioxidant Activity
- Dk—optical density in the absence of antioxidants (control);
- Do—optical density in the presence of antioxidants;
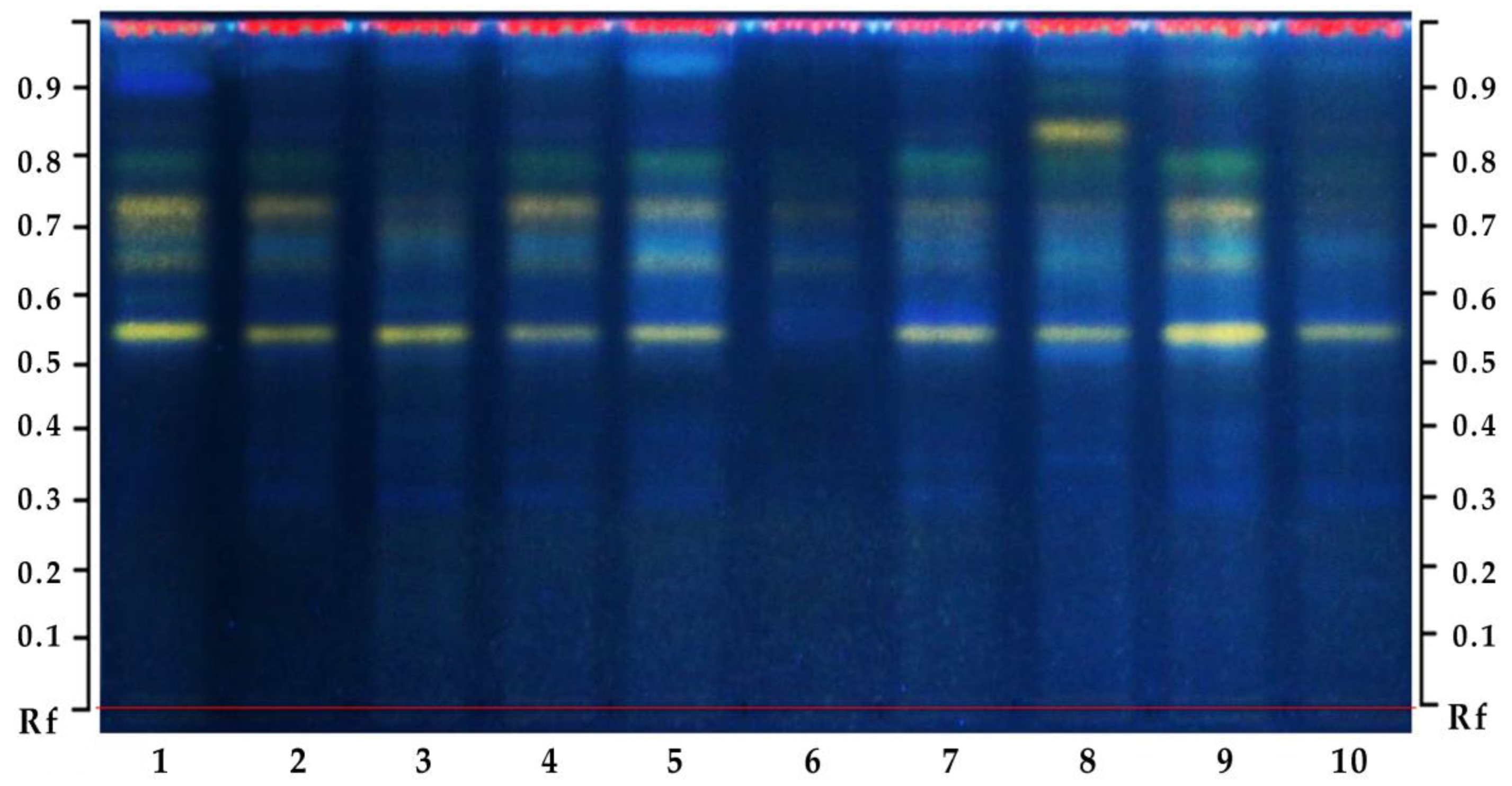
3.4.5. Biochemical Profiling of Plant Extracts with High-Performance Thin-Layer Chromatography
3.5. Field Testing of Bacteria on One-Year Seedlings of Q. robur
3.6. Photo Documentation, Digital and Statistical Data Processing
4. Conclusions
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
Appendix A


References
- Bonan, G.B. Forests and climate change: Forcings, feedbacks, and the climate benefits of forests. Science 2008, 320, 1444–1449. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shvidenko, A.; Buksha, I.; Krakovska, S.; Lakyda, P. Vulnerability of Ukrainian forests to climate change. Sustainability 2017, 9, 1152. [Google Scholar] [CrossRef] [Green Version]
- Seidl, R.; Thom, D.; Kautz, M.; Martin-Benito, D.; Peltoniemi, M.; Vacchiano, G.; Wild, J.; Ascoli, D.; Petr, M.; Honkaniemi, J.; et al. Forest disturbances under climate change. Nat. Clim. Chang. 2017, 7, 395–402. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Leach, J.E.; Triplett, L.R.; Argueso, S.T.; Trivedi, P. Communication in the phytobiome. Cell 2017, 169, 587–596. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gupta, A.; Mishra, R.; Rai, S.; Bano, A.; Pathak, N.; Fujita, M.; Kumar, M.; Hasanuzzaman, M. Mechanistic Insights of Plant Growth Promoting Bacteria Mediated Drought and Salt Stress Tolerance in Plants for Sustainable Agriculture. Int. J. Mol. Sci. 2022, 23, 3741. [Google Scholar] [CrossRef]
- Smith, S.A.; Tank, D.C.; Boulanger, L.A.; Bascom-Slack, C.A.; Eisenman, K.; Kingery, D.; Babbs, B.; Fenn, K.; Greene, J.S.; Hann, B.D.; et al. Bioactive endophytes warrant intensified exploration and conservation. PLoS ONE 2008, 3, e3052. [Google Scholar] [CrossRef] [Green Version]
- Halecker, S.; Wennrich, J.-P.; Rodrigo, S.; Andrée, N.; Rabsch, L.; Baschien, C.; Steinert, M.; Stadler, M.; Surup, F.; Schulz, B. Fungal endophytes for biocontrol of ash dieback: The antagonistic potential of Hypoxylon rubiginosum. Fungal Ecol. 2020, 45, 100918. [Google Scholar] [CrossRef]
- Khosro, M.; Yousef, S. Bacterial Biofertilizers for Sustainable Crop Production: A Review. ARPN J. Agric. Biol. Sci. 2012, 7, 307–316. [Google Scholar]
- Witzell, J.; Martín, J.A.; Blumenstein, K. Ecological Aspects of Endophyte-Based Biocontrol of Forest Diseases. In Advances in Endophytic Research; Verma, V.C., Gange, A.C., Eds.; Springer: Berlin, Germany, 2014; pp. 321–333. [Google Scholar] [CrossRef]
- Lei, Z.; Xiao, P.S.; Jian, S.W.; Hai, Y. Endophytic bacterial effects on seeds germination and mobilization of reserves ammodendron biofolium yan. Pak. J. Bot. 2017, 49, 2029–2035. [Google Scholar]
- Cregger, M.A.; Carper, D.L.; Christel, S.; Doktycz, M.J.; Labbé, J.; Michener, J.K.; Dove, N.C.; Johnston, E.R.; Moore, J.A.M.; Vélez, J.M.; et al. Plant–Microbe Interactions: From Genes to Ecosystems Using Populus as a Model System. Phytobiomes 2021, 5, 29–38. [Google Scholar] [CrossRef]
- Suhag, M. Potential of Biofertilizers to Replace Chemical Fertilizers. Int. Adv. Res. J. Sci. Eng. Technol. 2016, 3, 163–167. [Google Scholar]
- Frank, A.C.; Saldierna Guzmán, J.P.; Shay, J.E. Transmission of bacterial endophytes. Microorganisms 2017, 10, 70. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mohite, B. Isolation and characterization of indole acetic acid (IAA) producing bacteria from rhizospheric soil and its effect on plant growth. J. Soil Sci. Plant Nutr. 2013, 13, 638–649. [Google Scholar] [CrossRef]
- Karthik, M.; Pushpakanth, P.; Krishnamoorthy, R.; Senthilkumar, M. Endophytic bacteria associated with banana cultivars and their inoculation effect on plant growth. J. Hortic. Sci. Biotechnol. 2017, 92, 568–576. [Google Scholar] [CrossRef]
- Somal, M.K.; Karnwal, A. Effect of stress tolerance endophytic bacteria on the growth of Rheum emodi under abiotic stress. Plant Biol. 2022, 36000522. [Google Scholar] [CrossRef] [PubMed]
- Fort, T.; Pauvert, C.; Zanne, A.E.; Ovaskainen, O.; Caignard, T.; Barret, M.; Compant, S.; Hampe, A.; Delzon, S.; Vacher, C. Maternal effects shape the seed mycobiome in Quercus petraea. New Phytol. 2021, 230, 1594–1608. [Google Scholar] [CrossRef]
- Nguyen, M.H.; Yong, J.H.; Sung, H.J.; Lee, J.K. Screening of Endophytic Fungal Isolates Against Raffaelea quercus-mongolicae Causing Oak Wilt Disease in Korea. Mycobiology 2020, 48, 484–494. [Google Scholar] [CrossRef]
- Shestibratov, K.A.; Barano, O.Y.; Subbotina, N.M.; Lebedev, V.G.; Panteleev, S.V.; Krutovsky, K.V.; Padutov, V.E. Early Detection and Identification of the Main Fungal Pathogens for Resistance Evaluation of New Genotypes of Forest Trees. Forests 2018, 9, 732. [Google Scholar] [CrossRef] [Green Version]
- Yang, A.; Juzwik, J. Use of nested and real-time PCR for the detection of Ceratocystis fagacearum in the sapwood of diseased Oak species in Minnesota. Plant Dis. 2017, 101, 480–486. [Google Scholar] [CrossRef] [Green Version]
- Oliveira, A.L.M.; Urquiaga, S.; Döbereiner, J.; Baldani, J.I. The effect of inoculating endophytic N2-fixing bacteria on micropropagated sugarcane plants. Plant Soil 2002, 242, 205–215. [Google Scholar] [CrossRef]
- Iniguez, A.L.; Dong, Y.; Triplett, E.W. Nitrogen Fixation in Wheat Provided by Klebsiella pneumoniae 342. Mol. Plant-Microbe Interact. 2004, 17, 1078–1085. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chalk, P.M.; Lam, S.K.; Chen, D. The significance of endophytic and phyllospheric N2 fixation in forest trees: Evi-dence from stable (15N) and radioactive (13N) tracer studies. Trees 2022, 36, 1179. [Google Scholar] [CrossRef]
- Sessitsch, A.N.; Hardoim, P.A.; Döring, J.; Weilharter, A.; Krause, A.N.; Woyke, T.A.; Mitter, B.; Hauberg-Lotte, L.; Friedrich, F.; Rahalkar, M.; et al. Functional characteristics of an endophyte community colonizing rice roots as revealed by metagenomic analysis. Mol. Plant-Microbe Interact. MPMI 2012, 25, 28–36. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ambrose, C.; Varghese, C.; Subhash, J. Endophytic bacteria as a source of novel antibiotics: An overview. Pharmacogn. Rev. 2013, 7, 11–16. [Google Scholar]
- Hardoim, P.R.; van Overbeek, L.S.; Berg, G.; Pirttilä, A.M.; Compant, S.; Campisano, A.; Döring, M.; Sessitsch, A. The hidden world within plants: Ecological and evolutionary considerations for defining functioning of microbial endophytes. Microb. Mol. Biol. Rev. 2015, 79, 293–320. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wani, Z.A.; Ashraf, N.; Mohiuddin, T.; Riyaz-Ul-Hassan, S. Plant-endophyte symbiosis, an ecological perspective. Appl. Microbiol. Biotechnol. 2015, 99, 2955–2965. [Google Scholar] [CrossRef] [PubMed]
- Pandey, S.S.; Singh, S.; Pandey, H.; Srivastava, M.; Ray, T.; Soni, S.; Pandey, A.; Shanker, K.; Babu, C.S.V.; Banerjee, S.; et al. Endophytes of Withania somnifera modulate in planta content and the site of withanolide biosynthesis. Sci. Rep. 2018, 8, 5450. [Google Scholar] [CrossRef]
- García-Fraile, P.; Menéndez, E.; Rivas, R. Role of bacterial biofertilizers in agriculture and forestry. AIMS Bioeng. 2015, 2, 183–205. [Google Scholar] [CrossRef]
- Díaz Herrera, S.; Grossi, C.; Zawoznik, M.; Groppa, M.D. Wheat seeds harbour bacterial endophytes with potential as plant growth promoters and biocontrol agents of Fusarium graminearum. Microbiol. Res. 2016, 186–187, 37–43. [Google Scholar] [CrossRef]
- Borodai, V.; Kolomiiets, Y.; Likhanov, A.; Zelena, L.; Butsenko, L.; Shemetun, K.; Churilov, A.; Blume, Y. The Growth-promoting and Antipathogenic Effects of Microorganisms Isolated from Solanum nigrum L. and Inoculated in Solanum lycopersicum L. Open Agric. J. 2022, 16, e2208180. [Google Scholar] [CrossRef]
- Shahzad, R.; Waqas, M.; Khan, A.; Al-Hosni, K.; Kang, S.; Seo, C.; Lee, I. Indoleacetic acid production and plant growth promoting potential of bacterial endophytes isolated from rice (Oryza sativa L.) seeds. Acta Biol. Hung. 2017, 68, 175–186. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Orozco-Mosqueda, M.D.C.; Rocha-Granados, M.D.C.; Glick, B.R.; Santoyo, G. Microbiome engineering to improve biocontrol and plant growth-promoting mechanisms. Microbiological Res. 2018, 208, 25–31. [Google Scholar] [CrossRef]
- Rabiey, M.; Hailey, L.E.; Roy, S.R.; Grenz, K.; Al-Zadjali, M.A.; Barrett, G.A.; Jackson, R.W. Endophytes vs tree pathogens and pests: Can they be used as biological control agents to improve tree health? Eur. J. Plant Pathol. 2019, 155, 711–729. [Google Scholar] [CrossRef] [Green Version]
- Christian, N.; Herre, E.A.; Mejia, L.C.; Clay, K. Exposure to the leaf litter microbiome of healthy adults protects seedlings from pathogen damage. Proc. R. Soc. B Biol. Sci. 2017, 284, 20170641. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sarmiento, C.; Zalamea, P.; Dalling, J.; Davis, A.; Stump, S.; U’Ren, J.; Arnold, A. Soilborne fungi have host affinity and host-specific effects on seed germination and survival in a lowland tropical forest. Proc. Natl. Acad. Sci. USA 2017, 114, 11458–11463. [Google Scholar] [CrossRef] [Green Version]
- Ridout, M.E.; Schroeder, K.L.; Hunter, S.S.; Styer, J.; Newcombe, G. Priority effects of wheat seed endophytes on a rhizosphere symbiosis. Symbiosis 2019, 78, 19–31. [Google Scholar] [CrossRef]
- Uroz, S.; Buee, M.; Murat, C.; Frey-Klett, P.; Martin, F. Pyrosequencing reveals a contrasted bacterial diversity between oak rhizosphere and surrounding soil. Environ. Microbiol. 2010, 2, 281–288. [Google Scholar] [CrossRef] [PubMed]
- Kim, M.; Singh, D.; Lai-Hoe, A.; Go, R.; Abdul Rahim, R.; Ainuddin, A.N. Distinctive phyllosphere bacterial communities in tropical trees. Microb. Ecol. 2012, 63, 674–681. [Google Scholar] [CrossRef]
- Tanaka, A.; Tapper, B.A.; Popay, A.; Parker, E.J.; Scott, B. A symbiosis expressed non-ribosomal peptide synthetase from a mutualistic fungal endophyte of perennial ryegrass confers protection to the symbiotum from insect herbivory. Mol. Microbiol. 2005, 57, 1036–1050. [Google Scholar] [CrossRef]
- Vega, F.E.; Posada, F.; Aime, M.C.; Pava-Ripoll, M.; Infante, F.; Rehner, S.A. Entomopathogenic fungal endophytes. Biol. Control 2008, 46, 72–82. [Google Scholar] [CrossRef] [Green Version]
- Lugtenberg, B.J.J.; Caradus, J.R.; Johnson, L.J. Fungal endophytes for sustainable crop production. FEMS Microbiol. Ecol. 2016, 92, 194. [Google Scholar] [CrossRef] [PubMed]
- Lata, R.; Chowdhury, S.; Gond, S.K.; White, J.F., Jr. Induction of abiotic stress tolerance in plants by endophytic microbes. Lett. Appl. Microbiol. 2018, 66, 268–276. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lasa, A.V.; Maslinova, T.; Baldrian, P.; Fernandez-Lopez, M. Bacteria from the endosphere and rhizosphere of Quercus spp. use mainly cell wall-associated enzymes to decompose organic matter. PLoS ONE 2019, 14, e0214422. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Matsala, M.; Bilous, A.; Feshchenko, R.; Matiashuk, R.; Bilous, S.; Kovbasa, Y. Spatial and compositional structure of European oak urban forests in Kyiv city, Ukraine. J. For. Sci. Link Is Disabl. 2021, 67, 143–153. [Google Scholar] [CrossRef]
- Borruso, L.; Wellstein, C.; Bani, A.; Casagrande Bacchiocchi, S.; Margoni, A.; Tonin, R.; Zerbe, S.; Brusetti, L. Temporal shifts in endophyte bacterial community composition of sessile oak (Quercus petraea) are linked to foliar nitrogen, stomatal length, and herbivory. PeerJ 2018, 6, e5769. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Meaden, S.; Metcalf, C.J.E.; Koskella, B. The effects of host age and spatial location on bacterial community composition in the English oak tree (Quercus robur). Environ. Microbiol. Rep. 2016, 8, 649–658. [Google Scholar] [CrossRef] [Green Version]
- Pirttilä, A.M. Endophytic Bacteria in Tree Shoot Tissues and Their Effects on Host. In Endophytes of Forest Trees; Forestry Sciences; Pirttilä, A., Frank, A., Eds.; Springer: Berlin/Heidelberg, Germany, 2018; Volume 86. [Google Scholar]
- Rosenblueth, M.; Martínez-Romero, E. Bacterial endophytes and their interactions with hosts. Mol. Plant-Microbe Interact. MPMI 2006, 19, 827–837. [Google Scholar] [CrossRef] [Green Version]
- Nguyen, M.H.; Shin, K.C.; Lee, J.K. Fungal Community Analyses of Endophytic Fungi from Two Oak Species, Quercus mongolica and Quercus serrata, in Korea. Mycobiology 2021, 9, 385–395. [Google Scholar] [CrossRef]
- Novotný, D. Contribution to the endophytic mycobiota of aerial parts of oaks. Czech Mycol. 2022, 74, 111–121. [Google Scholar] [CrossRef]
- Sapp, M.; Lewis, E.; Moss, S.; Barrett, B.; Kirk, S.; Elphinstone, J.G.; Denman, S. Metabarcoding of Bacteria Associated with the Acute Oak Decline Syndrome in England. Forests 2016, 7, 95. [Google Scholar] [CrossRef] [Green Version]
- Brader, G.; Compant, S.; Mitter, B.; Trognitz, F.; Sessitsch, A. Metabolic potential of endophytic bacteria. Curr. Opin. Biotechnol. 2014, 27, 30–37. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Griffin, J.S.; Lu, N.; Sangwan, N.; Li, A.; Dsouza, M.; Stumpf, A.J.; Sevilla, T.; Culotti, A.; Keefer, L.L.; Kelly, J.J.; et al. Microbial diversity in an intensively managed landscape is structured by landscape connectivity. FEMS Microbiol. Ecol. 2017, 93, fix120. [Google Scholar] [CrossRef] [PubMed]
- Tashi-Oshnoei, F.; Harighi, B.; Abdollahzadeh, J. Isolation and identification of endophytic bacteria with plant growth promoting and biocontrol potential from oak trees. For. Pathol. 2017, 47, e12360. [Google Scholar] [CrossRef]
- Petrini, O.; Fisher, P.J. Occurrence of fungal endophyticin twigs of Salix fragilis and Quercus robur. Mycol. Res. 1990, 94, 1077–1080. [Google Scholar] [CrossRef]
- Kowalski, T.; Kehr, R.D. Fungal endophytes of living branch bases in several European tree species. In Endophytic Fungi in Grasses and Woody Plants, Systematics, Ecology and Evolution; Redlin, S.C., Carris, L.M., Eds.; APS: St. Paul, MN, USA, 1996; pp. 67–86. [Google Scholar]
- Gennaro, M.; Gonthier, P.; Nicolotti, G. Fungal endophytic communities in healthy and declining Quercus robur L. and Q. cerris L. trees in Northern Italy. J. Phytopathol. 2003, 151, 529–534. [Google Scholar] [CrossRef]
- Ragazzi, A.; Moricca, S.; Capretti, P.; Dellavalle, I.; Turco, E. Differences in composition of endophytic mycobiota in twigs and leaves of healthy and declining Quercus species in Italy. For. Path. 2003, 33, 31–38. [Google Scholar] [CrossRef]
- Fisher, P.J.; Petrini, O.; Petrini, L.E.; Sutton, B.C. Fungal endophytes from the leaves and twigs of Quercus ilex L. from England, Majorca and Switzerland. New Phytol. 1994, 127, 133–137. [Google Scholar] [CrossRef] [PubMed]
- Collado, J.; Platas, G.; Gonzales, I.; Peláez, F. Geographical and seasonal influences on the distribution of fungal endophytes in Quercus ilex. New Phytol. 1999, 144, 525–532. [Google Scholar] [CrossRef]
- Moricca, S.; Ragazzi, A. Species- and organ-specificity in endophytes colonizing healthy and declining Mediterranean oaks. Phytopathol. Mediterr. 2012, 51, 587–598. [Google Scholar]
- Linaldeddu, B.T.; Sirca, C.; Spano, D.; Franceschini, A. Variation of endophytic cork oak-associated fungal communities in relation to plant health and water stress. For. Pathol. 2011, 41, 193–201. [Google Scholar] [CrossRef] [Green Version]
- Costa, D.; Tavares, R.; Baptista, P.; Lino-Neto, T. Diversity of fungal endophytic community in Quercus suber L. under different climate scenarios. Rev. Cienc. Agrar. 2019, 41, 22–30. [Google Scholar]
- Ghasemi, S.; Khodaei, S.; Karimi, K.; Tavakoli, M.; Pertot, I.; Arzanlou, M. Biodiversity study of endophytic fungi associated with two Quercus species in Iran. For. Syst. 2019, 28, 12. [Google Scholar] [CrossRef]
- Moricca, S.; Ragazzi, A. Fungal endophytes in Mediterranean oak forests: A lesson from Discula quercina. Phytopathology 2008, 98, 380–386. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Terhonen, E.; Blumenstein, K.; Kovalchuk, A.; Asiegbu, F.O. Forest Tree Microbiomes and Associated Fungal Endophytes: Functional Roles and Impact on Forest Health. Forests 2019, 10, 42. [Google Scholar] [CrossRef] [Green Version]
- Gupta, G.; Parihar, S.S.; Ahirwar, N.K.; Snehi, S.K.; Singh, V. Plant Growth Promoting Rhizobacteria (PGPR): Current and Future Prospects for Development of Sustainable Agriculture. Microb. Biochem. Technol. 2015, 7, 96–102. [Google Scholar]
- Santoyo, G.; Moreno-Hagelsieb, G.; Orozco-Mosqueda Mdel, C.; Glick, B.R. Plant growth-promoting bacterial endophytes. Microbiol. Res. 2016, 183, 92–99. [Google Scholar] [CrossRef]
- Fernández-González, A.J.; Martínez-Hidalgo, P.; Cobo-Díaz, J.F.; Villadas, P.J.; Martínez-Molina, E.; Toro, N.; Tringe, S.G.; Fernández-López, M. The rhizosphere microbiome of burned holm-oak: Potential role of the genus Arthrobacter in the recovery of burned soils. Sci. Rep. 2017, 7, 6008. [Google Scholar] [CrossRef]
- Chaudhary, P.; Agri, U.; Chaudhary, A.; Kumar, A.; Kumar, G. Endophytes and their potential in biotic stress management and crop production. Front. Microbiol. 2022, 13, 933017. [Google Scholar] [CrossRef]
- Gathercole, L.A.P.; Nocchi, G.; Brown, N.; Coker, T.L.R.; Plumb, W.J.; Stocks, J.J.; Nichols, R.A.; Denman, S.; Buggs, R.J.A. Evidence for the Widespread Occurrence of Bacteria Implicated in Acute Oak Decline from Incidental Genetic Sampling. Forests 2021, 12, 1683. [Google Scholar] [CrossRef]
- Abdelfattah, A.; Wisniewski, M.; Schena, L.; Tack, A.J.M. Experimental evidence of microbial inheritance in plants and transmission routes from seed to phyllosphere and root. Environ. Microbiol. 2021, 23, 2199–2214. [Google Scholar] [CrossRef]
- Martínez-Arias, C.; Sobrino-Plata, J.; Medel, D.; Gil, L.; Martín, J.A.; Rodríguez-Calcerrada, J. Stem endophytes increase root development, photosynthesis, and survival of elm plantlets (Ulmus minor Mill.). J. Plant Physiol. 2021, 261, 153420. [Google Scholar] [CrossRef] [PubMed]
- Vujanovic, V.; Islam, M.N.; Daida, P. Transgenerational role of seed mycobiomean endosymbiotic fungal composition as a prerequisite to stress resilience and adaptive phenotypes in Triticum. Sci. Rep. 2019, 9, 18483. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ahlholm, J.U.; Helander, M.; Henriksson, J.; Metzler, M.; Saikkonen, K. Environmental conditions and host genotype direct genetic diversity of Venturia ditricha, a fungal endophyte of birch trees. Evolution 2002, 56, 1566–1573. [Google Scholar] [PubMed]
- Khare, E.; Mishra, J.; Kumar Arora, N. Multifaceted Interactions Between Endophytes and Plant: Developments and Prospects. J. Front. Microbiol. 2018, 9, 2732. [Google Scholar] [CrossRef]
- Shao, J.; Li, S.; Zhang, N.; Cui, X.; Zhou, X.; Zhang, G.; Shen, Q.; Zhang, R. Analysis and cloning of the synthetic pathway of the phytohormone indole-3-acetic acid in the plant-beneficial Bacillus amyloliquefaciens SQR9. Microb. Cell Fact. 2015, 4, 130. [Google Scholar] [CrossRef] [Green Version]
- Keswani, C.; Singh, S.P.; Cueto, L.; García-Estrada, C.; Mezaache-Aichour, S.; Glare, T.R.; Borriss, R.; Singh, S.P.; Blázquez, M.A.; Sansinenea, E. Auxins of microbial origin and their use in agriculture. Appl. Microbiol. Biotechnol. 2020, 104, 8549–8565. [Google Scholar] [CrossRef] [PubMed]
- Vaitiekunaite, D.; Kuusiene, S.; Beniušyte, E. Oak (Quercus robur) Associated Endophytic Paenibacillus sp. Promotes Poplar (Populus spp.) Root Growth In Vitro. Microorganisms 2021, 9, 1151. [Google Scholar] [CrossRef]
- Brooks, D.S.; Gonzales, C.F.; Appel, D.N.; Filer, T.H. Evaluation of endophytic bacteria as potential biological control agents for oak wilt. Biol. Control 1994, 4, 373–381. [Google Scholar] [CrossRef]
- Chandrasekaran, M.; Chun, S.C. Expression of PR-protein genes and induction of defense-related enzymes by Bacillus subtilis CBR05 in tomato (Solanum lycopersicum) plants challenged with Erwinia carotovora sub sp. carotovora. Biosci. Biotechnol. Biochem. 2016, 80, 2277–2283. [Google Scholar] [CrossRef] [Green Version]
- Heidari, M.; Golpayegani, A. Effects of water stress and inoculation with plant growth promoting rhizobacteria (PGPR) on antioxidant status and photosynthetic pigments in basil (Ocimum basilicum L.). J. Saudi Soc. Agric. Sci. 2012, 11, 57–61. [Google Scholar] [CrossRef] [Green Version]
- Gusain, Y.S.; Singh, U.S.; Sharma, A.K. Bacterial mediated amelioration of drought stress in drought tolerant and susceptible cultivars of rice (Oryza sativa L.). Afr. J. Biotechnol. 2015, 14, 764–773. [Google Scholar]
- Verma, S.K.; White, J.F. Seed Endophytes: Biology and Biotechnology; Springer: Berlin/Heidelberg, Germany, 2019. [Google Scholar]
- Vasilchenko, A.S.; Poshvina, D.V.; Sidorov, R.Y.; Iashnikov, A.V.; Rogozhin, E.A.; Vasilchenko, A.V. Oak bark (Quercus sp. cortex) protects plants through the inhibition of quorum sensing mediated virulence of Pectobacterium carotovorum. World J. Microbiol. Biotechnol. 2022, 38, 184. [Google Scholar] [CrossRef] [PubMed]
- Liu, L.; Wen, W.; Zhang, R.; Wei, Z.; Deng, Y.; Xiao, J.; Zhang, M. Complex enzyme hydrolysis releases antioxidative phenolics from rice bran. Food Chem. 2016, 214, 1–8. [Google Scholar] [CrossRef] [PubMed]
- Likhanov, A.F.; Sereda, O.V.; Gryb, V.M.; Melnyk, V.I.; Osadchuk, L.S.; Yuskevych, T. Biochemical markers of vital biodestruction in common oak (Quercus robur). Biosyst. Divers. 2019, 27, 314–321. [Google Scholar] [CrossRef]
- Deng, Y.; Lu, S. Biosynthesis and Regulation of Phenylpropanoids in Plants. Crit. Rev. Plant Sci. 2017, 36, 257–290. [Google Scholar] [CrossRef]
- Carroll, G.C. Fungal endophytes in stems and leaves: From latent pathogen to mutualistic symbiont. Ecology 1988, 969, 2. [Google Scholar] [CrossRef]
- Carroll, G.C. Fungal associates of woody plants as insect antagonists in leaves and stems. In Microbial Mediation of Plant-Herbivore Interactions; Barbosa, P., Krischik, V.A., Jones, C.G., Eds.; John Wiley & Sons, Inc.: Hoboken, NJ, USA, 1991; pp. 253–271. [Google Scholar]
- Fontana, D.C.; Paula, S.; Torres, A.G.; Souza, V.; Pascholati, S.F.; Schmidt, D.; Dourado Neto, D. Endophytic Fungi: Biological Control and Induced Resistance to Phytopathogens and Abiotic Stresses. Pathogens 2021, 10, 570. [Google Scholar] [CrossRef]
- Radhakrishnan, R.; Hashem, A.; Abd Allah, E.F. Bacillus: A Biological Tool for Crop Improvement through Bio-Molecular Changes in Adverse Environments. Front. Physiol. 2017, 8, 667. [Google Scholar] [CrossRef] [Green Version]
- Bilous, S.; Prysiazhniuk, L.; Chernii, S.; Melnyk, S.; Marchuk, Y.; Likhanov, A. Genetic characterisation of centuries-old oak and linden trees using SSR markers. Folia For. Pol. 2022, 64, 58–68. [Google Scholar] [CrossRef]
- Borkar, S.G. Laboratory Techniques in Plant Bacteriology, 1st ed.; CRC Press: Boca Raton, FL, USA, 2017. [Google Scholar]
- Patyka, V.P.; Pasichnyk, L.A.; Gvozdyak, R.I. Phytopathogenic bacteria. Res. Methods 2017, 2, 432. [Google Scholar]
- Don, J.; Brenner, N.R.; Krieg, J.T.; Staley, G.M.; Garrity, D.R.; Boone, P.V.; Goodfellow, M.; Rainey, F.A.; Karl-Heinz, S. Bergey’s Manual of Systematic Bacteriology, 2nd ed.; Biomedical and Life Sciences; Springer: New York, NY, USA, 2005; Volume 4, p. 976. [Google Scholar] [CrossRef]
- Maheshwari, R. Fungi: Experimental Methods in Biology, 2nd ed.; CRC Press: Boca Raton, FL, USA, 2011. [Google Scholar]
- Tamura, K.; Stecher, G.; Kumar, S. MEGA11: Molecular Evolutionary Genetics Analysis version 11. Mol. Biol. Evol. 2021, 38, 3022–3027. [Google Scholar] [CrossRef] [PubMed]
- Singleton, V.L.; Orthofer, R.; Lamuela-Raventos, R. Analysis of Total Phenols and Other Oxidation Substrates and Antioxidants by Means of Folin-Ciocalteu Reagent. Methods Enzymol. 1999, 299, 152–178. [Google Scholar]
- Cimpoiu, C.; Cristea, M.; Hosu, A.; Sandru, M.; Seserman, L. Antioxidant activity prediction and classification of some teas using neutral networks. Food Chem. 2011, 127, 1323–1328. [Google Scholar] [CrossRef] [PubMed]
- Brand-Williams, W.; Cuvelier, M.E.; Berset, C.L.W.T. Use of a Free Radical Method to Evaluate Antioxidant Activity. LWT-Food Sci. Technol. 1995, 28, 25–30. [Google Scholar] [CrossRef]







| Phytopathogens | EI | R1 | R2 | R3 | R4 | R1/Rav | GIZ |
|---|---|---|---|---|---|---|---|
| Botrytis | Q2 | 20.3 ± 0.58 | 17.3 ± 1.00 | 16.3 ± 0.84 a | 17.7 ± 0.41 | 1.2 ± 0.08 | 1.3 ± 0.11 |
| cinerea | Q7 | 22.2 ± 0.95 | 13.1 ± 0.19 b | 11.0 ± 0.25 b | 11.6 ± 0.32 b | 1.9 ± 0.10 | 8.0 ± 0.46 |
| BCIBL-2143 | Contr | 8.8 ± 0.60 | 9.0 ± 0.65 | 8.87 ± 0.46 | 8.8 ± 0.20 | 1.0 ± 0.04 | - |
| Sclerotinia | Q2 | 23.5 ± 1.10 | 15.8 ± 0.20 b | 13.4 ± 0.78 b | 13.2 ± 1.03 b | 1.7 ± 0.11 | 1.5 ± 0.21 |
| sclerotiorum | Q7 | 22.7 ± 1.79 | 16.6 ± 0.37 b | 14.7 ± 0.47 b | 13.8 ± 0.24 b | 1.5 ± 0.12 | 4.0 ± 0.26 |
| SIBL-2135 | Contr | 20.5 ± 1.63 | 21.3 ± 0.86 | 18.4 ± 1.93 | 18.9 ± 1.22 | 1.1 ± 0.05 | - |
| Fusarium | Q2 | 19.5 ± 0.47 | 13.6 ± 0.53 a | 12.9 ± 0.61 a | 12.7 ± 0.56 b | 1.5 ± 0.08 | 5.1 ± 0.48 |
| tricinctum | Q7 | 18.2 ± 1.23 | 7.2 ± 0.39 b | 7.4 ± 0.24 b | 7.4 ± 0.40 b | 2.5 ± 0.16 | 13.2 ± 0.36 |
| FTIBL-2151 | Contr | 20.6 ± 0.85 | 20.0 ± 0.49 | 19.7 ± 0.20 | 19.4 ± 0.71 | 1.0 ± 0.03 | - |
| Samples | Phenols, mg·g−1 | Flavonoids, mg·g−1 | Antioxidants, μM-eq | AAi/Phi * | |||
|---|---|---|---|---|---|---|---|
| Samples | Experim | Samples | Experim | Samples | Experim | Experim | |
| Control | 101.3 ± 1.51 | 125.7 ± 3.84 | 3.8 ± 0.14 | 2.0 ± 0.08 | 196.0 ± 7.39 | 187.1 ± 5.72 | 1.00 ± 0.01 |
| Q2 | 98.9 ± 3.02 | 102.0 ± 3.40 | 2.7 ± 0.10 b | 1.8 ± 0.06 b | 210.5 ± 7.94 | 242.1 ± 7.40 | 1.59 ± 0.01 b |
| Q7 | 84.3 ± 2.78 | 77.8 ± 2.59 | 1.9 ± 0.07 b | 1.7 ± 0.06 b | 174.5 ± 6.58 | 164.8 ± 5.49 | 1.44 ± 0.01 b |
| Q2/Q7 (mix 1:1) | 143.6 ± 4.74 b | 104.4 ± 3.48 | 2.3 ± 0.07 b | 1.9 ± 0.06 b | 252.2 ± 9.51 a | 232.6 ± 7.75 | 1.51 ± 0.01 b |
| Pectobacterium | 140.5 ± 4.64 b | 157.3 ± 5.24 b | 2.4 ± 0.09 b | 2.3 ± 0.08 b | 259.6 ± 7.94 b | 254.1 ± 8.47 b | 1.10 ± 0.01 a |
| Pseuodomonas | 150.2 ± 4.96 b | 175.9 ± 5.86 b | 2.3 ± 0.07 b | 1.7 ± 0.06 b | 257.7 ± 9.72 b | 256.5 ± 8.55 b | 0.99 ± 0.01 |
| Q5 | 173.0 ± 5.71 b | 194.3 ± 6.48 b | 0.4 ± 0.02 b | 0.3 ± 0.01 | 257.5 ± 7.87 b | 246.3 ± 8.21 a | 0.86 ± 0.01 b |
| Q6 | 118.8 ± 3.92 | 92.7 ± 3.09 | 3.2 ± 0.12 a | 2.2 ± 0.07 b | 201.4 ± 7.60 | 163.9 ± 5.46 | 1.20 ± 0.01 b |
| EpQ1 | 143.0 ± 4.72 b | 158.3 ± 5.28 b | 3.4 ± 0.13 b | 3.4 ± 0.11 b | 242.6 ± 9.15 | 238.6 ± 7.95 | 1.02 ± 0.01 |
| EpQ2 | 165.9 ± 5.47 b | 173.6 ± 5.79 b | 2.5 ± 0.08 b | 2.4 ± 0.08 b | 245.7 ± 7.51 | 247.5 ± 9.33 a | 0.96 ± 0.01 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Bilous, S.; Likhanov, A.; Boroday, V.; Marchuk, Y.; Zelena, L.; Subin, O.; Bilous, A. Antifungal Activity and Effect of Plant-Associated Bacteria on Phenolic Synthesis of Quercus robur L. Plants 2023, 12, 1352. https://0-doi-org.brum.beds.ac.uk/10.3390/plants12061352
Bilous S, Likhanov A, Boroday V, Marchuk Y, Zelena L, Subin O, Bilous A. Antifungal Activity and Effect of Plant-Associated Bacteria on Phenolic Synthesis of Quercus robur L. Plants. 2023; 12(6):1352. https://0-doi-org.brum.beds.ac.uk/10.3390/plants12061352
Chicago/Turabian StyleBilous, Svitlana, Artur Likhanov, Vira Boroday, Yurii Marchuk, Liubov Zelena, Oleksandr Subin, and Andrii Bilous. 2023. "Antifungal Activity and Effect of Plant-Associated Bacteria on Phenolic Synthesis of Quercus robur L." Plants 12, no. 6: 1352. https://0-doi-org.brum.beds.ac.uk/10.3390/plants12061352