
Functional Characterization of ShK Domain-Containing Protein in the Plant-Parasitic Nematode Bursaphelenchus xylophilus

Abstract
:1. Introduction
2. Results
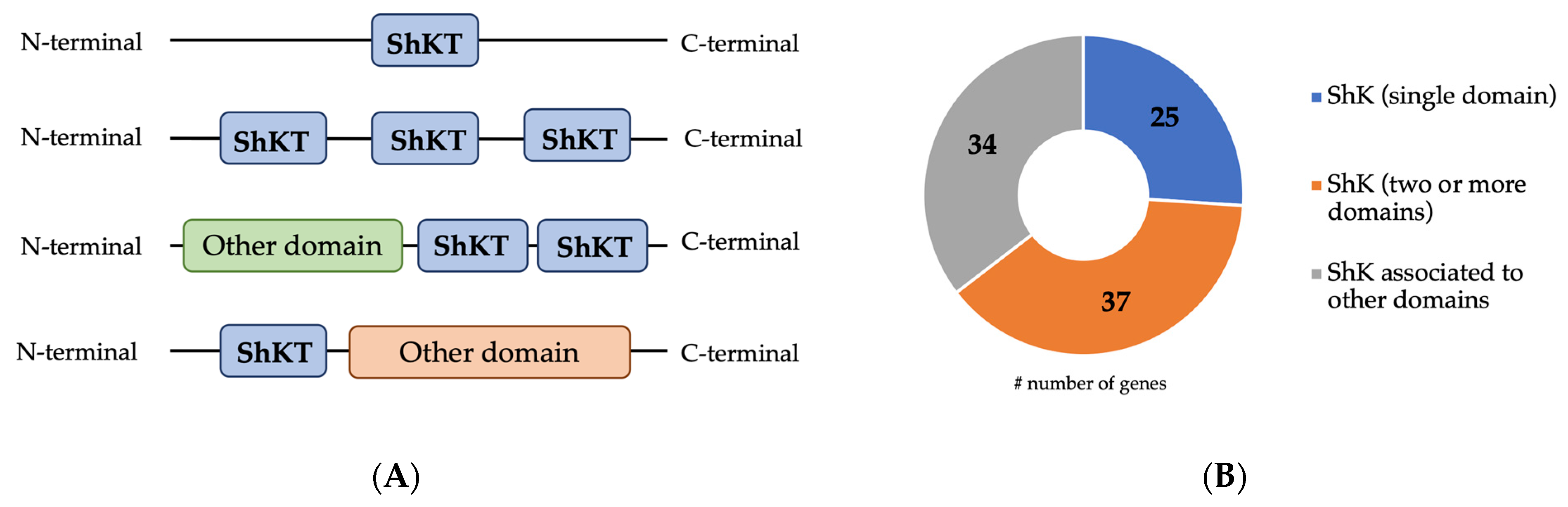
2.1. Characterization of ShK Domain-Containing Protein from B. xylophilus
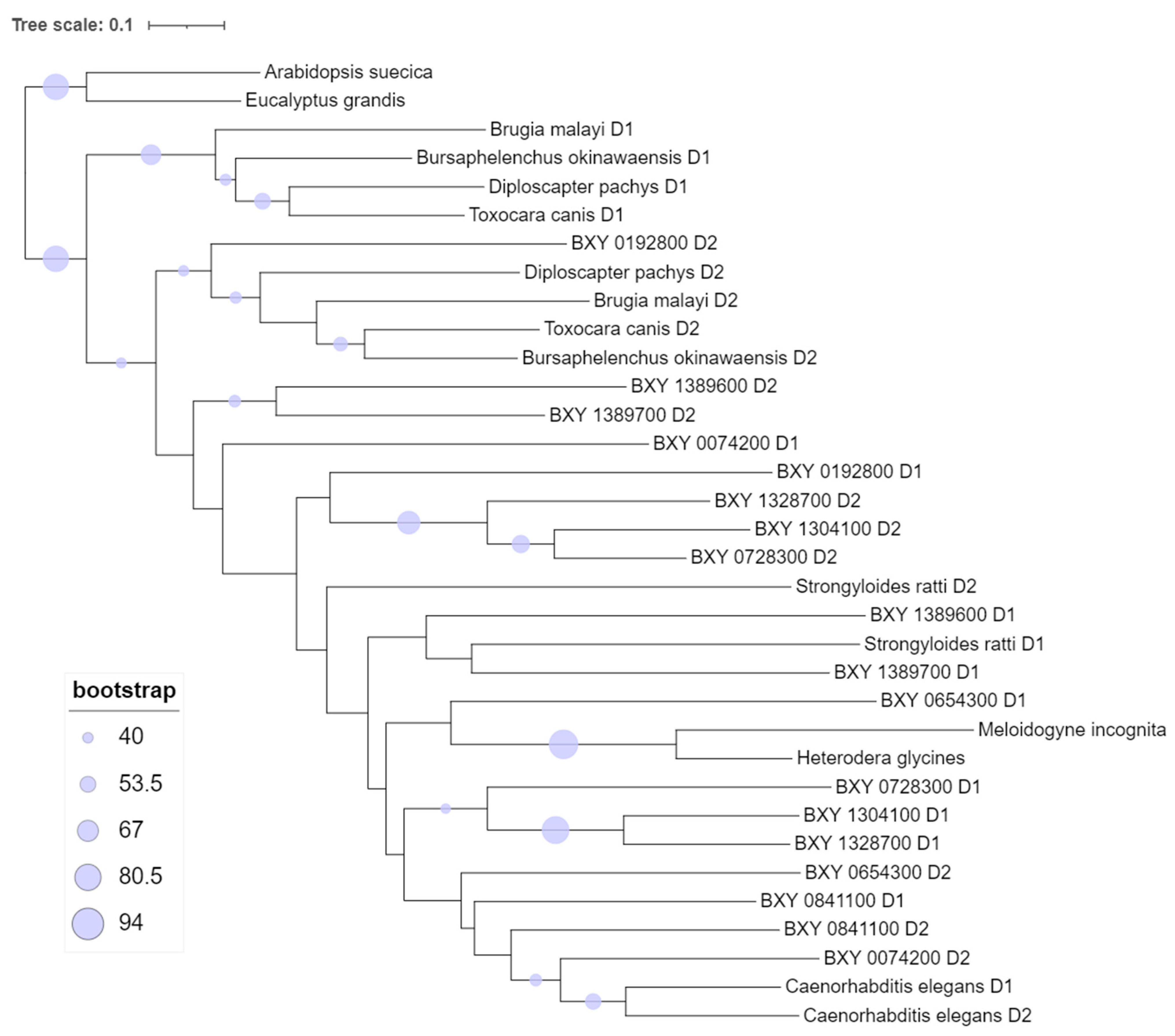
2.2. In Silico Analysis and Phylogeny of the Nine Candidate B. xylophilus Putatively Specific Transcripts
2.3. Spatial Expression of the Candidate ShK Domain-Containing Proteins
2.4. High Gene Expression of B. xylophilus ShK Domain-Containing Proteins under Oxidative Stress
3. Discussion
4. Materials and Methods
4.1. Biological Material
4.2. Phylogenetic Analysis of B. xylophilus ShK Domain-Containing Protein Sequences
4.3. Validation of the Candidate Genes
4.4. Oxidative Stress Assay
4.5. RNA Extraction and Gene Relative Expression Profile
4.6. In Situ Hybridization Assay
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
References
- Boyd, I.L.; Freer-Smith, P.H.; Gilligan, C.A.; Godfray, H.C.J. The Consequence of Tree Pests and Diseases for Ecosystem Services. Science 2013, 342, 1235773. [Google Scholar] [CrossRef]
- Jones, J.T.; Haegeman, A.; Danchin, E.G.J.; Gaur, H.S.; Helder, J.; Jones, M.G.K.; Kikuchi, T.; Manzanilla-López, R.; Palomares-Rius, E.J.; Wesemael, W.M.L.; et al. Top 10 plant-parasitic nematodes in molecular plant pathology. Mol. Plant Pathol. 2013, 14, 946–961. [Google Scholar] [CrossRef]
- PM 7/4 (4) Bursaphelenchus xylophilus. EPPO Bull. 2023, 53, 156–183. [CrossRef]
- Mamiya, Y. Pathology of the pine wilt disease caused by Bursaphelenchus xylophilus. Annu. Rev. Phytopathol. 1983, 21, 201–220. [Google Scholar] [CrossRef]
- Mota, M.M.; Braasch, H.; Bravo, M.A.; Penas, A.C.; Burgermeister, W.; Metge, K.; Sousa, E. First report of Bursaphelenchus xylophilus in Portugal and in Europe. Nematology 1999, 1, 727–734. [Google Scholar] [CrossRef]
- Robertson, L.; Cobacho Arcos, S.; Escuer, M.; Santiago Merino, R.; Esparrago, G.; Abelleira, A.; Navas, A. Incidence of the pinewood nematode Bursaphelenchus xylophlius Steiner & Buhrer, 1934 (Nickle, 1970) in Spain. Nematology 2011, 13, 755–757. [Google Scholar] [CrossRef]
- Vicente, C.S.L.; Espada, M.; Vieira, P.; Mota, M. Pine wilt disease: A threat to European forestry. Eur. J. Plant Pathol. 2012, 133, 89–99. [Google Scholar] [CrossRef]
- Vieira, P.; Gleason, C. Plant-parasitic nematode effectors—Insights into their diversity and new tools for their identification. Curr. Opin. Plant Biol. 2019, 50, 37–43. [Google Scholar] [CrossRef] [PubMed]
- Espada, M.; Silva, A.C.; Eves-van den Akker, S.; Cock, P.J.A.; Mota, M.; Jones, J.T. Identification and characterization of parasitism genes from the pinewood nematode Bursaphelenchus xylophilus reveals a multi-layered detoxification strategy. Mol. Plant Pathol. 2016, 17, 286–295. [Google Scholar] [CrossRef] [PubMed]
- Tsai, I.J.; Tanaka, R.; Kanzaki, N.; Akiba, M.; Yokoi, T.; Espada, M.; Jones, J.T.; Kikuchi, T. Transcriptional and morphological changes in the transition from mycetophagous to phytophagous phase in the plant-parasitic nematode Bursaphelenchus xylophilus. Mol. Plant Pathol. 2016, 17, 77–83. [Google Scholar] [CrossRef]
- Rangaraju, S.; Khoo, K.; Feng, Z.P.; Crossley, G.; Nugent, D.; Khaytin, I.; Chi, V.; Pham, C.; Calabresi, P.; Pennington, M.; et al. Potassium Channel Modulation by A Toxin Domain in Matrix Metalloprotease 23. J. Biol. Chem. 2009, 285, 9124–9136. [Google Scholar] [CrossRef] [PubMed]
- Castañeda, O.; Sotolongo, V.; Amor, A.M.; Stöcklin, R.; Anderson, A.J.; Harvey, A.L.; Engström, A.; Wernstedt, C.; Karlsson, E. Characterization of a potassium channel toxin from the Caribbean Sea anemone Stichodactyla helianthus. Toxicon 1995, 33, 603–613. [Google Scholar] [CrossRef] [PubMed]
- Cotton, J.; Crest, M.; Bouet, F.; Alessandri, N.; Gola, M.; Forest, E.; Karlsson, E.; Castañeda, O.; Harvey, A.L.; Vita, C.; et al. A Potassium-channel toxin from the sea anemone bunodosoma granulifera, an inhibitor for Kv1 channels—Revision of the amino acid sequence, disulfide-bridge assignment, chemical synthesis, and biological activity. Eur. J. Biochem. 1997, 244, 192–202. [Google Scholar] [CrossRef] [PubMed]
- Tudor, J.E.; Pallaghy, P.K.; Pennington, M.W.; Norton, R.S. Solution structure of ShK toxin, a novel potassium channel inhibitor from a sea anemone. Nat. Struct. Biol. 1996, 3, 317–320. [Google Scholar] [CrossRef]
- Loukas, A.; Hintz, M.; Linder, D.; Mullin, N.P.; Parkinson, J.; Tetteh, K.K.; Maizels, R.M. A family of secreted mucins from the parasitic nematode Toxocara canis bears diverse mucin domains but shares similar flanking six-cysteine repeat motifs. J. Biol. Chem. 2000, 275, 39600–39607. [Google Scholar] [CrossRef]
- Prentis, P.J.; Pavasovic, A.; Norton, R.S. Sea Anemones: Quiet Achievers in the Field of Peptide Toxins. Toxins 2018, 10, 30. [Google Scholar] [CrossRef]
- von Reumont, B.M.; Campbell, L.I.; Richter, S.; Hering, L.; Sykes, D.; Hetmank, J.; Jenner, R.A.; Bleidorn, C. A Polychaete’s powerful punch: Venom gland transcriptomics of Glycera reveals a complex cocktail of toxin homologs. Genome Biol. Evol. 2014, 6, 2406–2423. [Google Scholar] [CrossRef]
- Verdes, A.; Simpson, D.; Holford, M. Are Fireworms Venomous? Evidence for the Convergent Evolution of Toxin Homologs in Three Species of Fireworms (Annelida, Amphinomidae). Genome Biol. Evol. 2018, 10, 249–268. [Google Scholar] [CrossRef]
- Gerdol, M.; Cervelli, M.; Mariottini, P.; Oliverio, M.; Dutertre, S.; Modica, M.V. A Recurrent Motif: Diversity and Evolution of ShKT Domain Containing Proteins in the Vampire Snail Cumia reticulata. Toxins 2019, 11, 106. [Google Scholar] [CrossRef] [PubMed]
- Suh, J.; Hutter, H. A survey of putative secreted and transmembrane proteins encoded in the C. elegans genome. BMC Genom. 2012, 13, 333. [Google Scholar] [CrossRef] [PubMed]
- Lima, A.K.; Dhillon, H.; Dillman, A.R. ShK-Domain-Containing Protein from a Parasitic Nematode Modulates Drosophila melanogaster Immunity. Pathogens 2022, 11, 1094. [Google Scholar] [CrossRef]
- Frias, J.; Toubarro, D.; Bjerga, G.E.K.; Puntervoll, P.; Vicente, J.B.; Reis, R.L.; Simões, N. A ShK-like Domain from Steinernema carpocapsae with Bioinsecticidal Potential. Toxins 2022, 14, 754. [Google Scholar] [CrossRef]
- Vieira, P.; Myers, R.Y.; Pellegrin, C.; Wram, C.; Hesse, C.; Maier, T.; Shao, J.; Koutsovoulos, G.D.; Zasada, I.; Matsumoto, T.; et al. Targeted transcriptomics reveals signatures of large-scale independent origins and concerted regulation of effector genes in Radopholus similis. PLoS Pathog. 2021, 17, e1010036. [Google Scholar] [CrossRef]
- Vieira, P.; Shao, J.; Vijayapalani, P.; Maier, T.; Pellegrin, C.; Eves-van den Akker, S.; Baum, T.J.; Nemchinov, L. A new esophageal gland transcriptome reveals signatures of large scale de novo effector birth in the root lesion nematode Pratylenchus penetrans. BMC Genom. 2020, 21, 738. [Google Scholar] [CrossRef]
- Kumar, A.; Fitoussi, N.; Sanadhya, P.; Sichov, N.; Bucki, P.; Bornstein, M.; Belausuv, E.; Miyara, B.S. Two Candidate Meloidogyne javanica Effector Genes, MjShKT and MjPUT3: A Functional Investigation of Their Roles in Regulating Nematode Parasitism. Mol. Plant-Microbe. Interact. 2023, 36, 79–94. [Google Scholar] [CrossRef]
- Habash, S.S.; Radakovic, Z.S.; Vankova, R.; Siddique, S.; Dobrev, P.; Gleason, C.; Grundler, F.M.W.; Elashry, A. Heterodera schachtii Tyrosinase-like protein—A novel nematode effector modulating plant hormone homeostasis. Sci. Rep. 2017, 7, 6874. [Google Scholar] [CrossRef]
- Cantacessi, C.; Mitreva, M.; Campbell, B.E.; Hall, R.S.; Young, N.D.; Jex, A.R.; Ranganathan, S.; Gasser, R.B. First transcriptomic analysis of the economically important parasitic nematode, Trichostrongylus colubriformis, using a next-generation sequencing approach. Infect. Genet. Evol. 2010, 10, 1199–1207. [Google Scholar] [CrossRef]
- Cahalan, M.D.; Chandy, K.G. The functional network of ion channels in T lymphocytes. Immunol. Rev. 2009, 231, 59–87. [Google Scholar] [CrossRef] [PubMed]
- Beeton, C.; Pennington, M.W.; Norton, R.S. Analogs of the Sea Anemone Potassium Channel Blocker ShK for the Treatment of Autoimmune Diseases. Inflamm Allergy Drug Targets 2011, 10, 313–321. [Google Scholar] [CrossRef] [PubMed]
- Chhabra, S.; Chang, S.C.; Nguyen, H.M.; Huq, R.; Tanner, M.R.; Londono, L.M.; Estrada, R.; Dhawan, V.; Chauhan, S.; Upadhyay, S.K.; et al. Kv1.3 channel-blocking immunomodulatory peptides from parasitic worms: Implications for autoimmune diseases. FASEB J. 2014, 28, 3952–3964. [Google Scholar] [CrossRef] [PubMed]
- Dreyer, I.; Uozumi, N. Potassium channels in plant cells. FEBS J. 2011, 278, 4293–4303. [Google Scholar] [CrossRef]
- Joshi, I.; Kumar, A.; Singh, A.K.; Kohli, D.; Raman, K.V.; Sirohi, A.; Chaudhury, A.; Jain, P.K. Development of nematode resistance in Arabidopsis by HD-RNAi-mediated silencing of the effector gene Mi-msp2. Sci. Rep. 2019, 9, 17404. [Google Scholar] [CrossRef] [PubMed]
- Kikuchi, T.; Cotton, J.A.; Dalzell, J.J.; Hasegawa, K.; Kanzaki, N.; McVeigh, P.; Takanashi, T.; Tsai, I.J.; Assefa, S.A.; Cock, P.J.A.; et al. Genomic insights into the origin of parasitism in the emerging plant pathogen Bursaphelenchus xylophilus. PLoS Pathog. 2011, 7, e1002219. [Google Scholar] [CrossRef] [PubMed]
- Tanaka, S.E.; Dayi, M.; Maeda, Y.; Tsai, I.J.; Tanaka, R.; Bligh, M.; Takeuchi-Kaneko, Y.; Fukuda, K.; Kanzaki, N.; Kikuchi, T. Stage-specific transcriptome of Bursaphelenchus xylophilus reveals temporal regulation of effector genes and roles of the dauer-like stages in the lifecycle. Sci. Rep. 2019, 9, 6080. [Google Scholar] [CrossRef] [PubMed]
- Vicente, C.S.L.; Ikuyo, Y.; Shinya, R.; Mota, M.; Hasegawa, K. Catalases Induction in High Virulence Pinewood Nematode Bursaphelenchus xylophilus under Hydrogen Peroxide-Induced Stress. PLoS ONE 2015, 10, e0123839. [Google Scholar] [CrossRef] [PubMed]
- McNeilly, T.N.; Frew, D.; Burgess, S.T.G.; Wright, H.; Bartley, D.J.; Bartley, Y.; Nisbet, A.J. Niche-specifc gene expression in a parasitic nematode; increased expression of immunomodulators in Teladorsagia circumcincta larvae derived from host mucosa. Sci. Rep. 2017, 7, 7214. [Google Scholar] [CrossRef] [PubMed]
- Espada, M.; Eves-van den Akker, S.; Maier, T.; Paramasivan, V.; Baum, T.J.; Mota, M.; Jones, J.T. STATAWAARS: A promoter motif associated with spatial expression in the major effector-producing tissues of the plant-parasitic nematode Bursaphelenchus xylophilus. BMC Genom. 2018, 19, 553. [Google Scholar] [CrossRef] [PubMed]
- Columbus-Shenkar, Y.Y.; Sachkova, M.Y.; Macrander, J.; Fridrich, A.; Modepalli, V.; Reitzel, A.M.; Sunagar, K.; Moran, Y. Dynamics of Venom Composition Across a Complex Life Cycle. Elife 2018, 7, e35014. [Google Scholar] [CrossRef] [PubMed]
- Schafer, W. Nematode nervous systems. Curr. Biol. 2016, 26, 937–980. [Google Scholar] [CrossRef]
- Espada, M.; Jones, J.T.; Mota, M. Characterization of glutathione S-transferases from the pine wood nematode, Bursaphelenchus xylophilus. Nematology 2016, 18, 697–709. [Google Scholar] [CrossRef]
- Evans, A.A.F. Mass culture of mycophagous nematodes. J. Nematol. 1970, 2, 99–100. [Google Scholar]
- Southey, J.F. Laboratory Methods for Work with Plant and Soil Nematodes; Ministry of Agriculture Fisheries and Food, HMSO: London, UK, 1986.
- McGinnis, S.; Madden, T.L. BLAST: At the core of a powerful and diverse set of sequence analysis tools. Nucleic Acids Res. 2004, 32, 20–25. [Google Scholar] [CrossRef]
- Boratyn, G.M.; Camacho, C.; Cooper, P.S.; Coulouris, G.; Fong, A.; Ma, N.; Madden, T.L.; Matten, W.T.; McGinnis, S.D.; Merezhuk, Y.; et al. BLAST: A more efficient report with usability improvements. Nucleic Acids Res. 2013, 41, 29–33. [Google Scholar] [CrossRef]
- Blum, M.; Chang, H.; Chuguransky, S.; Grego, T.; Kandasaamy, S.; Mitchell, A.; Nuka, G.; Paysan-Lafosse, T.; Qureshi, M.; Raj, S.; et al. The InterPro protein families and domains database: 20 years on. Nucleic Acids Res. 2020, 49, 344–354. [Google Scholar] [CrossRef]
- Teufel, F.; Almagro Armenteros, J.J.; Johansen, A.R.; Gíslason, M.H.; Pihl, S.I.; Tsirigos, K.D.; Winther, O.; Brunak, S.; von Heijne, G.; Nielsen, H. SignalP 6.0 predicts all five types of signal peptides using protein language models. Nat. Biotechnol. 2022, 40, 1023–1025. [Google Scholar] [CrossRef]
- Katoh, K.; Standley, D. MAFFT multiple sequence alignment software version 7: Improvements in performance and usability. Mol. Biol. Evol. 2013, 30, 772–780. [Google Scholar] [CrossRef] [PubMed]
- Capella-Gutiérrez, S.; Silla-Martínez, J.M.; Gabaldón, T. trimAl: A tool for automated alignment trimming in large-scale phylogenetic analyses. Bioinformatics 2009, 25, 1972–1973. [Google Scholar] [CrossRef] [PubMed]
- Letunic, I.; Bork, P. Interactive Tree of Life (iTOL) v5: An online tool for phylogenetic tree display and annotation. Nucleic Acids Res. 2021, 49, 293–296. [Google Scholar] [CrossRef] [PubMed]
- Hall, T.A. BioEdit: A User-Friendly Biological Sequence Alignment Editor and Analysis Program for Windows 95/98/NT. Nucleic Acids Symp. Ser. 1999, 41, 95–98. [Google Scholar] [CrossRef]
- Untergasser, A.; Cutcutache, I.; Koressaar, T.; Ye, J.; Faircloth, B.C.; Remm, M.; Rozen, S.G. Primer3—New capabilities and interfaces. Nucleic Acids Res. 2012, 40, e115. [Google Scholar] [CrossRef] [PubMed]
- Livak, K.J.; Schmittgen, T.D. Analysis of relative gene expression data using real-time quantitative PCR and the 2−ΔΔCT method. Methods 2001, 25, 402–408. [Google Scholar] [CrossRef] [PubMed]
- de Boer, J.M.; Yan, Y.; Smant, G.; Davis, E.L.; Baum, T.J. In-situ hybridization to messenger RNA in Heterodera glycines. J. Nemat. 1998, 30, 309–312. [Google Scholar]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Gene ID | GL * | IN | TL * | PS ** | SP | Sequence Similarity ID (NCBI Accession Number) | BLAST e-Value |
|---|---|---|---|---|---|---|---|
| BXY_0192800 | 630 | 2 | 510 | 169 | Yes, 1−17 | [CAD5228099.1] Bursaphelenchus xylophilus, 100% | 4 × 10−123 |
| BXY_0728300 | 502 | 1 | 429 | 142 | Yes, 1−16 | [CAD5230694.1] Bursaphelenchus xylophilus, 100% | 6 × 10−94 |
| BXY_1389600 | 534 | 1 | 492 | 163 | Yes, 1−21 | [CAD5234029.1] Bursaphelenchus xylophilus, 100% | 1 × 10−111 |
| BXY_0074200 | 447 | 0 | 447 | 148 | Yes, 1−22 | [CAD5228719.1] Bursaphelenchus xylophilus, 100% | 3 × 10−105 |
| BXY_0841100 | 1170 | 2 | 669 | 229 | Yes, 1−17 | [CAD5208706.1] Bursaphelenchus xylophilus, 100% | 8 × 10−157 |
| BXY_1304100 | 922 | 1 | 423 | 140 | Yes, 1−16 | [CAD5233070.1] Bursaphelenchus xylophilus, 100% | 2 × 10−98 |
| BXY_1328700 | 673 | 1 | 444 | 147 | Yes, 1−23 | [CAD5233069.1] Bursaphelenchus xylophilus, 100% | 3 × 10−99 |
| BXY_0654300 | 3250 | 4 | 1926 | 641 | Yes, 1−20 | [CAD5235767.1] Bursaphelenchus xylophilus, 100% | <0 |
| BXY_1389700 | 3240 | 2 | 2625 | 874 | Yes, 1−21 | [CAD5234030.1] Bursaphelenchus xylophilus, 100% | <0 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Mendonça, M.; Vicente, C.S.L.; Espada, M. Functional Characterization of ShK Domain-Containing Protein in the Plant-Parasitic Nematode Bursaphelenchus xylophilus. Plants 2024, 13, 404. https://0-doi-org.brum.beds.ac.uk/10.3390/plants13030404
Mendonça M, Vicente CSL, Espada M. Functional Characterization of ShK Domain-Containing Protein in the Plant-Parasitic Nematode Bursaphelenchus xylophilus. Plants. 2024; 13(3):404. https://0-doi-org.brum.beds.ac.uk/10.3390/plants13030404
Chicago/Turabian StyleMendonça, Madalena, Cláudia S. L. Vicente, and Margarida Espada. 2024. "Functional Characterization of ShK Domain-Containing Protein in the Plant-Parasitic Nematode Bursaphelenchus xylophilus" Plants 13, no. 3: 404. https://0-doi-org.brum.beds.ac.uk/10.3390/plants13030404