
Signals and Machinery for Mycorrhizae and Cereal and Oilseed Interactions towards Improved Tolerance to Environmental Stresses


 , , , and
, , , and 
Abstract
:1. Introduction
2. Building Resilience: Harnessing Mycorrhizal Symbiosis for Enhanced (a)biotic Stress Tolerance in Cereal and Oilseed Crops
2.1. Cereal Mycorrhizal Responses to Stressed Environments
2.1.1. Cereal Mycorrhizal Responses to Abiotic Stresses
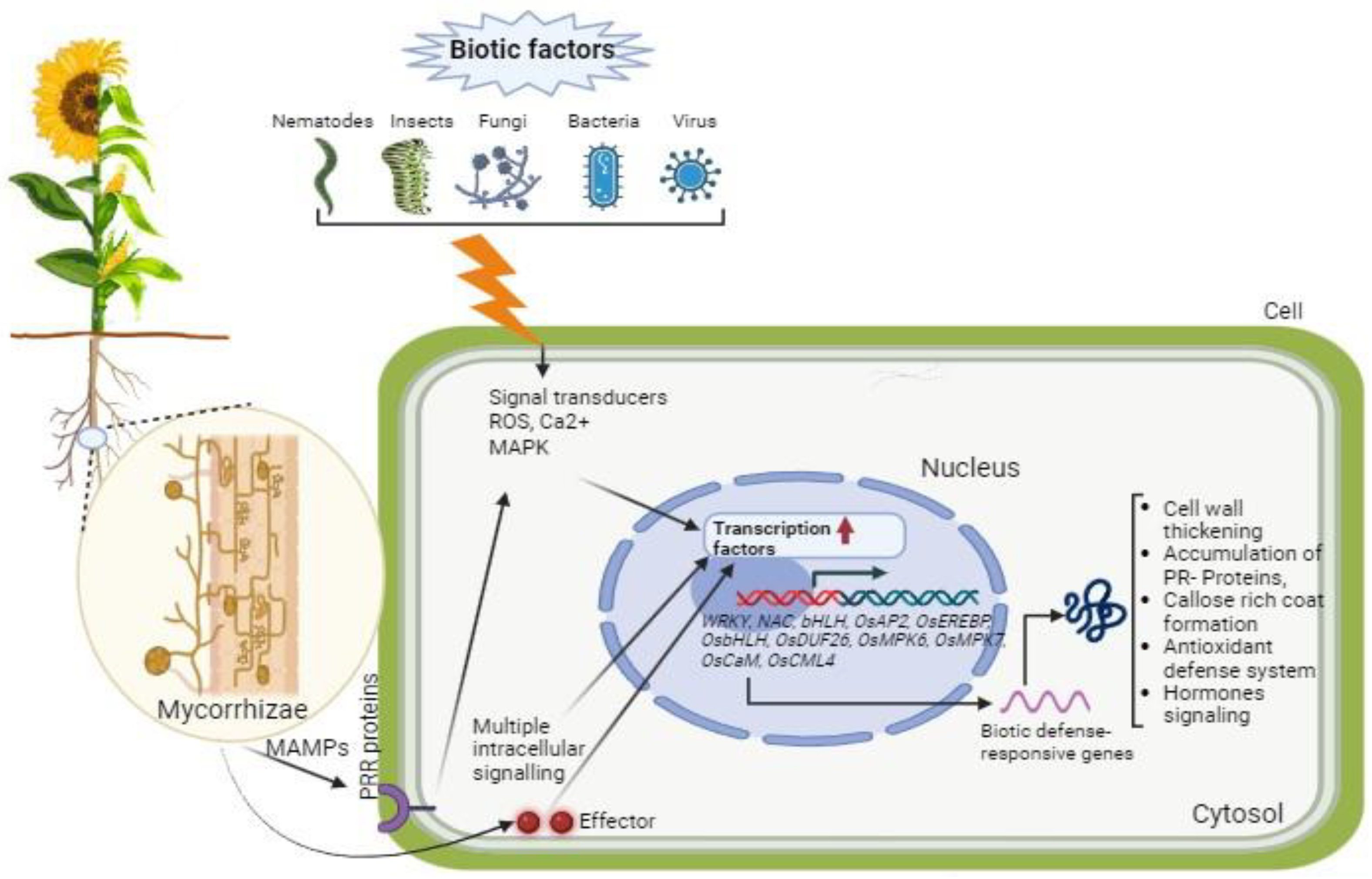
2.1.2. Cereal Mycorrhizal Responses to Biotic Stresses
2.2. Oilseed Mycorrhizal Responses to Stressed Environments
2.2.1. Oilseed Mycorrhizal Responses to Abiotic Stresses

{kind=link}
{kind=link}
| Stress | Plant | AMF Species | AMF Colonization Effects | Ref. |
|---|---|---|---|---|
| Drought | ||||
| Soybean (Glycine max) | R. irregularis | - Enhanced TSS, proline content, and MAPK transcripts. | [94] | |
| Flax (Linum usitatissimum) | F. mosseae, R. intraradices | - Enhanced leaf P content and oil yield. | [100] | |
| Sesame (Sesamum indicium) | F. mosseae, R. intraradices | - Improved Chl index, and N, P, K, Zn, Fe, and Cu content. | [98] | |
| Soybean (Glycine max) | R. intraradices, R. clarus, R. aggregatum, S. deserticola, F. mosseae, O. etunicatum | - Improved biomass, Chl content, gs, leaf water relations, and N, P, K, S, Mn, and Cu content. | [97] | |
| Flax (Linum usitatissimum) | F. mosseae, R. intraradices | - Improved vesicle diameter, yield, and SOD, APX, and POX activity. | [101] | |
| Sesame (Sesamum indicum) | F. mosseae, R. intraradices | - Improved TSP, P, Chl, flavonoid contents, and seed/oil yield. | [99] | |
| Soybean (Glycine max) | P. occulum, G. gigantea, F. mosseae, C. etunicatum, R. clarus | - Higher growth traits (pod number, seed number, and seed DM) and oilseed proline. | [95] | |
| Soybean (Glycine max) | G. clarum, G. mosseae, Gigaspora margarita | - Enhanced yield, and seed CAT and POX activity. - Decreased MDA and proline contents. - Up-regulated CAT and POX expression and down-regulated proline metabolism genes (P5CS, P5CR, PDH, and P5CDH). | [112] | |
| Sesame (Sesamum indicum) | R. intraradices, F. mosseae | - Improved grain yield, oil content, and N and P content. | [103] | |
| Soybean (Glycine max) | G. mosseae | - Improved glucose exudation, and β-glucosidase and acid phosphomonoesterase. | [113] | |
| Soybean (Glycine max) | R. clarus | - Improved plant height, water potential, WUE, Fv/Fm, and N and K content. | [102] | |
| Salinity | ||||
| Sunflower (Helianthus annuus) | R. irregularis | - Lower Na+ and MDA content. - Improved biomass and nutritional profile (K+, Mg2+, Ca2+, N, P), soil enzyme activities (CAT, dehydrogenase, phosphatase, fluorescein diacetate hydrolysis). | [104] | |
| Safflower (Carthamus tinctorius) | R. intraradices, F. moseae | - Improved shoot and root DM, stem and root heights, proline, pigment, P, N, Mg contents, and antioxidant enzyme activity. | [105] | |
| Soybean (Glycine max) | F. mosseae, R. intraradices, C. etunicatum | - Improved nitrogenase and IAA synthesis, and lower H2O2 and MDA content. | [108] | |
| Iberian dragon’head (Lallemantia iberica) | F. mosseae | - Ameliorated seeds’ oil and mucilage composition. | [106] | |
| Groundnut (Arachis hypogaea) | R. irregularis, F. mosseae | - Reduced MDA content. - Improved An, RWC, plant height, osmolyte production, SOD, POX, CAT, APX, protein, and pod yielding. | [107] | |
| Heavy Metals | ||||
| Sunflower (Helianthus annuus) | G. intraradices | - Enhanced P, Chl, carotenoid, SOD, and PPO. - Reduced MDA, As, Cr, and Ni translocation. | [109] | |
| Soybean (Glycine max) | R. intraradices | - Lowered Cd accumulation in roots. - Promoted P and Fe abundance in roots. | [110] | |
| Soybean (Glycine max) | F. mosseae | - Boosted growth, yield, and P assimilation. - Decreased Cu, Pb, and Zn translocation. | [111] | |
| Soybean (Glycine Max) | R. Intraradices | - Improved growth, P acquisition, and grain yield under Cu, Pb, and Zn soil pollution. - Reduced translocation of the HMs. | [111] | |
2.2.2. Oilseed Mycorrhizal Responses to Biotic Stresses
3. Molecular Strategies Contributing to Cereal and Oilseed Tolerance to Environmental Stresses
3.1. Molecular Mechanisms behind Cereal Mycorrhiza Responses
3.1.1. Molecular Mechanisms behind Cereal Mycorrhiza Responses to Abiotic Stresses
3.1.2. Molecular Mechanisms behind Cereal Mycorrhiza Responses to Biotic Stresses
3.2. Molecular Mechanisms behind Oilseed Mycorrhiza Responses to Stressed Environments
3.2.1. Molecular Mechanisms behind Oilseed Mycorrhiza Responses to Abiotic Stresses
3.2.2. Molecular Mechanisms behind Oilseed Mycorrhiza Responses to Biotic Stresses
4. Concluding Remarks
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Khoury, C.K.; Jarvis, A.; Jones, A.D. Trade and Its Trade-Offs in the Food System. Nat. Food 2020, 1, 665–666. [Google Scholar] [CrossRef]
- Fones, H.N.; Bebber, D.P.; Chaloner, T.; Kay, W.T.; Steinberg, G.; Gurr, S.J. Threats to Global Food Security from Emerging Fungal and Oomycete Crop Pathogens. Nat. Food 2020, 1, 332–342. [Google Scholar] [CrossRef]
- Cogato, A.; Meggio, F.; De Antoni Migliorati, M.; Marinello, F. Extreme Weather Events in Agriculture: A Systematic Review. Sustainability 2019, 11, 2547. [Google Scholar] [CrossRef]
- Wang, J.-W.; Luo, Z.-H.; Xu, W.; Ding, J.-F.; Zheng, T.-L. Transformation of Dimethyl Phthalate Esters (DMPEs) by a Marine Red Yeast Rhodotorula Mucilaginosa Isolated from Deep Sea Sediments of the Atlantic Ocean. Int. Biodeterior. Biodegrad. 2016, 109, 223–228. [Google Scholar] [CrossRef]
- Zhu, J.-K. Abiotic Stress Signaling and Responses in Plants. Cell 2016, 167, 313–324. [Google Scholar] [CrossRef] [PubMed]
- Zhang, W.; Sun, K.; Shi, R.-H.; Yuan, J.; Wang, X.-J.; Dai, C.-C. Auxin Signalling of Arachis hypogaea Activated by Colonization of Mutualistic Fungus Phomopsis liquidambari Enhances Nodulation and N2-Fixation: Auxin in Fungus-Legume-Rhizobium Interactions. Plant Cell Environ. 2018, 41, 2093–2108. [Google Scholar] [CrossRef] [PubMed]
- Grassini, P.; Eskridge, K.M.; Cassman, K.G. Distinguishing between Yield Advances and Yield Plateaus in Historical Crop Production Trends. Nat. Commun. 2013, 4, 2918. [Google Scholar] [CrossRef] [PubMed]
- Smith, S.E.; Read, D. Mineral Nutrition, Toxic Element Accumulation and Water Relations of Arbuscular Mycorrhizal Plants. In Mycorrhizal Symbiosis; Elsevier: Amsterdam, The Netherlands, 2008; pp. 145–187. ISBN 978-0-12-370526-6. [Google Scholar]
- Genre, A.; Lanfranco, L.; Perotto, S.; Bonfante, P. Unique and Common Traits in Mycorrhizal Symbioses. Nat. Rev. Microbiol. 2020, 18, 649–660. [Google Scholar] [CrossRef]
- Shi, J.; Wang, X.; Wang, E. Mycorrhizal Symbiosis in Plant Growth and Stress Adaptation: From Genes to Ecosystems. Annu. Rev. Plant Biol. 2023, 74, 569–607. [Google Scholar] [CrossRef]
- Huang, D.; Ma, M.; Wang, Q.; Zhang, M.; Jing, G.; Li, C.; Ma, F. Arbuscular Mycorrhizal Fungi Enhanced Drought Resistance in Apple by Regulating Genes in the MAPK Pathway. Plant Physiol. Biochem. 2020, 149, 245–255. [Google Scholar] [CrossRef]
- Ait-El-Mokhtar, M.; Baslam, M.; Ben-Laouane, R.; Anli, M.; Boutasknit, A.; Mitsui, T.; Wahbi, S.; Meddich, A. Alleviation of Detrimental Effects of Salt Stress on Date Palm (Phoenix dactylifera L.) by the Application of Arbuscular Mycorrhizal Fungi and/or Compost. Front. Sustain. Food Syst. 2020, 4, 131. [Google Scholar] [CrossRef]
- Anli, M.; Baslam, M.; Tahiri, A.; Raklami, A.; Symanczik, S.; Boutasknit, A.; Ait-El-Mokhtar, M.; Ben-Laouane, R.; Toubali, S.; Ait Rahou, Y.; et al. Biofertilizers as Strategies to Improve Photosynthetic Apparatus, Growth, and Drought Stress Tolerance in the Date Palm. Front. Plant Sci. 2020, 11, 516818. [Google Scholar] [CrossRef]
- Begum, N.; Qin, C.; Ahanger, M.A.; Raza, S.; Khan, M.I.; Ashraf, M.; Ahmed, N.; Zhang, L. Role of Arbuscular Mycorrhizal Fungi in Plant Growth Regulation: Implications in Abiotic Stress Tolerance. Front. Plant Sci. 2019, 10, 1068. [Google Scholar] [CrossRef]
- Wahab, A.; Muhammad, M.; Munir, A.; Abdi, G.; Zaman, W.; Ayaz, A.; Khizar, C.; Reddy, S.P.P. Role of Arbuscular Mycorrhizal Fungi in Regulating Growth, Enhancing Productivity, and Potentially Influencing Ecosystems under Abiotic and Biotic Stresses. Plants 2023, 12, 3102. [Google Scholar] [CrossRef]
- Ben-Laouane, R.; Baslam, M.; Ait-El-Mokhtar, M.; Anli, M.; Boutasknit, A.; Ait-Rahou, Y.; Toubali, S.; Mitsui, T.; Oufdou, K.; Wahbi, S.; et al. Potential of Native Arbuscular Mycorrhizal Fungi, Rhizobia, and/or Green Compost as Alfalfa (Medicago sativa) Enhancers under Salinity. Microorganisms 2020, 8, 1695. [Google Scholar] [CrossRef]
- Frew, A. Contrasting Effects of Commercial and Native Arbuscular Mycorrhizal Fungal Inoculants on Plant Biomass Allocation, Nutrients, and Phenolics. Plants People Planet 2021, 3, 536–540. [Google Scholar] [CrossRef]
- Smith, G.R.; Finlay, R.D.; Stenlid, J.; Vasaitis, R.; Menkis, A. Growing Evidence for Facultative Biotrophy in Saprotrophic Fungi: Data from Microcosm Tests with 201 Species of Wood-decay Basidiomycetes. New Phytol. 2017, 215, 747–755. [Google Scholar] [CrossRef]
- Dowarah, B.; Gill, S.S.; Agarwala, N. Arbuscular Mycorrhizal Fungi in Conferring Tolerance to Biotic Stresses in Plants. J. Plant Growth Regul. 2022, 41, 1429–1444. [Google Scholar] [CrossRef]
- Bahadur, A.; Batool, A.; Nasir, F.; Jiang, S.; Mingsen, Q.; Zhang, Q.; Pan, J.; Liu, Y.; Feng, H. Mechanistic Insights into Arbuscular Mycorrhizal Fungi-Mediated Drought Stress Tolerance in Plants. Int. J. Mol. Sci. 2019, 20, 4199. [Google Scholar] [CrossRef] [PubMed]
- Bhatta, M.; Morgounov, A.; Belamkar, V.; Wegulo, S.N.; Dababat, A.A.; Erginbas-Orakci, G.; El Bouhssini, M.; Gautam, P.; Poland, J.; Akci, N.; et al. Genome-Wide Association Study for Multiple Biotic Stress Resistance in Synthetic Hexaploid Wheat. Int. J. Mol. Sci. 2019, 20, 3667. [Google Scholar] [CrossRef] [PubMed]
- Zheng, F.L.; Liang, S.M.; Chu, X.N.; Yang, Y.L.; Wu, Q.S. Mycorrhizal Fungi Enhance Flooding Tolerance of Peach through Inducing Proline Accumulation and Improving Root Architecture. Plant Soil Environ. 2020, 66, 624–631. [Google Scholar] [CrossRef]
- Meddich, A. Biostimulants for Resilient Agriculture—Improving Plant Tolerance to Abiotic Stress: A Concise Review. Gesunde Pflanz. 2022, 75, 709–727. [Google Scholar] [CrossRef]
- Diagne, N.; Ngom, M.; Djighaly, P.I.; Fall, D.; Hocher, V.; Svistoonoff, S. Roles of Arbuscular Mycorrhizal Fungi on Plant Growth and Performance: Importance in Biotic and Abiotic Stressed Regulation. Diversity 2020, 12, 370. [Google Scholar] [CrossRef]
- Halford, N.G.; Curtis, T.Y.; Chen, Z.; Huang, J. Effects of Abiotic Stress and Crop Management on Cereal Grain Composition: Implications for Food Quality and Safety. J. Exp. Bot. 2015, 66, 1145–1156. [Google Scholar] [CrossRef]
- Chikkaputtaiah, C.; Debbarma, J.; Baruah, I.; Havlickova, L.; Deka Boruah, H.P.; Curn, V. Molecular Genetics and Functional Genomics of Abiotic Stress-Responsive Genes in Oilseed Rape (Brassica napus L.): A Review of Recent Advances and Future. Plant Biotechnol. Rep. 2017, 11, 365–384. [Google Scholar] [CrossRef]
- Slimani, A.; Ait-El-Mokhtar, M.; Ben-Laouane, R.; Boutasknit, A.; Anli, M.; Abouraicha, E.F.; Oufdou, K.; Meddich, A.; Baslam, M. Molecular and Systems Biology Approaches for Harnessing the Symbiotic Interaction in Mycorrhizal Symbiosis for Grain and Oil Crop Cultivation. Int. J. Mol. Sci. 2024, 25, 912. [Google Scholar] [CrossRef]
- Lenoir, I.; Fontaine, J.; Lounès-Hadj Sahraoui, A. Arbuscular Mycorrhizal Fungal Responses to Abiotic Stresses: A Review. Phytochemistry 2016, 123, 4–15. [Google Scholar] [CrossRef] [PubMed]
- Sharma, N.; Yadav, K.; Cheema, J.; Badda, N.; Aggarwal, A. Arbuscular mycorrhizal symbiosis and water stress: A critical review. Pertanika J. Trop. Agric. Sci. 2015, 38, 427–453. [Google Scholar]
- Zhao, R.; Guo, W.; Bi, N.; Guo, J.; Wang, L.; Zhao, J.; Zhang, J. Arbuscular Mycorrhizal Fungi Affect the Growth, Nutrient Uptake and Water Status of Maize (Zea mays L.) Grown in Two Types of Coal Mine Spoils under Drought Stress. Appl. Soil Ecol. 2015, 88, 41–49. [Google Scholar] [CrossRef]
- Li, H.; Luo, N.; Zhang, L.J.; Zhao, H.M.; Li, Y.W.; Cai, Q.Y.; Wong, M.H.; Mo, C.H. Do Arbuscular Mycorrhizal Fungi Affect Cadmium Uptake Kinetics, Subcellular Distribution and Chemical Forms in Rice? Sci. Total. Environ. 2016, 571, 1183–1190. [Google Scholar] [CrossRef]
- Zhang, Y.C.; Wang, P.; Wu, Q.H.; Zou, Y.N.; Bao, Q.; Wu, Q.S. Arbuscular Mycorrhizas Improve Plant Growth and Soil Structure in Trifoliate Orange under Salt Stress. Arch. Agron. Soil Sci. 2017, 63, 491–500. [Google Scholar] [CrossRef]
- Zhang, F.; Zou, Y.N.; Wu, Q.S. Quantitative Estimation of Water Uptake by Mycorrhizal Extraradical Hyphae in Citrus under Drought Stress. Sci. Hortic. 2018, 229, 132–136. [Google Scholar] [CrossRef]
- Quiroga, G.; Erice, G.; Aroca, R.; Chaumont, F.; Ruiz-Lozano, J.M. Enhanced Drought Stress Tolerance by the Arbuscular Mycorrhizal Symbiosis in a Drought-Sensitive Maize Cultivar Is Related to a Broader and Differential Regulation of Host Plant Aquaporins than in a Drought-Tolerant Cultivar. Front. Plant Sci. 2017, 8, 1056. [Google Scholar] [CrossRef]
- Bernardo, L.; Carletti, P.; Badeck, F.W.; Rizza, F.; Morcia, C.; Ghizzoni, R.; Rouphael, Y.; Colla, G.; Terzi, V.; Lucini, L. Metabolomic Responses Triggered by Arbuscular Mycorrhiza Enhance Tolerance to Water Stress in Wheat Cultivars. Plant Physiol. Biochem. 2019, 137, 203–212. [Google Scholar] [CrossRef] [PubMed]
- Kamali, S.; Mehraban, A. Effects of Nitroxin and Arbuscular Mycorrhizal Fungi on the Agro-Physiological Traits and Grain Yield of Sorghum (Sorghum bicolor L.) under Drought Stress Conditions. PLoS ONE 2021, 15, e0243824. [Google Scholar] [CrossRef] [PubMed]
- Moradi Tarnabi, Z.; Iranbakhsh, A.; Mehregan, I.; Ahmadvand, R. Impact of Arbuscular Mycorrhizal Fungi (AMF) on Gene Expression of Some Cell Wall and Membrane Elements of Wheat (Triticum aestivum L.) under Water Deficit Using Transcriptome Analysis. Physiol. Mol. Biol. Plants 2020, 26, 143–162. [Google Scholar] [CrossRef] [PubMed]
- Gutjahr, C. Phytohormone Signaling in Arbuscular Mycorhiza Development. Curr. Opin. Plant Biol. 2014, 20, 26–34. [Google Scholar] [CrossRef] [PubMed]
- Etemadi, M.; Gutjahr, C.; Couzigou, J.M.; Zouine, M.; Lauressergues, D.; Timmers, A.; Audran, C.; Bouzayen, M.; Bécard, G.; Combier, J.P. Auxin Perception Is Required for Arbuscule Development in Arbuscular Mycorrhizal Symbiosis. Plant Physiol. 2014, 166, 281–292. [Google Scholar] [CrossRef]
- Martín-Rodríguez, J.Á.; León-Morcillo, R.; Vierheilig, H.; Ocampo, J.A.; Ludwig-Müller, J.; García-Garrido, J.M. Ethylene-Dependent/Ethylene-Independent ABA Regulation of Tomato Plants Colonized by Arbuscular Mycorrhiza Fungi. New Phytol. 2011, 190, 193–205. [Google Scholar] [CrossRef] [PubMed]
- Foo, E.; Ross, J.J.; Jones, W.T.; Reid, J.B. Plant Hormones in Arbuscular Mycorrhizal Symbioses: An Emerging Role for Gibberellins. Ann. Bot. 2013, 111, 769–779. [Google Scholar] [CrossRef] [PubMed]
- Chareesri, A.; De Deyn, G.B.; Sergeeva, L.; Polthanee, A.; Kuyper, T.W. Increased Arbuscular Mycorrhizal Fungal Colonization Reduces Yield Loss of Rice (Oryza sativa L.) under Drought. Mycorrhiza 2020, 30, 315–328. [Google Scholar] [CrossRef] [PubMed]
- Mathur, S.; Tomar, R.S.; Jajoo, A. Arbuscular Mycorrhizal Fungi (AMF) Protects Photosynthetic Apparatus of Wheat under Drought Stress. Photosynth. Res. 2019, 139, 227–238. [Google Scholar] [CrossRef] [PubMed]
- Symanczik, S.; Lehmann, M.F.; Wiemken, A.; Boller, T.; Courty, P.-E. Effects of Two Contrasted Arbuscular Mycorrhizal Fungal Isolates on Nutrient Uptake by Sorghum bicolor under Drought. Mycorrhiza 2018, 28, 779–785. [Google Scholar] [CrossRef] [PubMed]
- Le Pioufle, O.; Ganoudi, M.; Calonne-salmon, M.; Dhaou, F.B.; Declerck, S. Rhizophagus irregularis MUCL 41833 Improves Phosphorus Uptake and Water Use Efficiency in Maize Plants During Recovery From Drought Stress. Front. Plant Sci. 2019, 10, 897. [Google Scholar] [CrossRef] [PubMed]
- Sun, J.; Yang, L.; Yang, X.; Wei, J.; Li, L.; Guo, E.; Kong, Y. Using Spectral Reflectance to Estimate the Leaf Chlorophyll Content of Maize Inoculated With Arbuscular Mycorrhizal Fungi Under Water Stress. Front. Plant Sci. 2021, 12, 646173. [Google Scholar] [CrossRef]
- Sun, X.; Shi, J.; Ding, G. Combined Effects of Arbuscular Mycorrhiza and Drought Stress on Plant Growth and Mortality of Forage Sorghum. Appl. Soil Ecol. 2017, 119, 384–391. [Google Scholar] [CrossRef]
- Pan, J.; Huang, C.; Peng, F.; Zhang, W.; Luo, J.; Ma, S.; Xue, X. Effect of Arbuscular Mycorrhizal Aungi (AMF) and Plant Growth-Promoting Bacteria (PGPR) Inoculations on Elaeagnus angustifolia L. in Saline Soil. Appl. Sci. 2020, 10, 945. [Google Scholar] [CrossRef]
- Bitterlich, M.; Franken, P.; Graefe, J. Atmospheric Drought and Low Light Impede Mycorrhizal Effects on Leaf Photosynthesis —A Glasshouse Study on Tomato under Naturally Fluctuating Environmental Conditions. Mycorrhiza 2019, 29, 13–28. [Google Scholar] [CrossRef]
- Tyagi, J.; Varma, A.; Pudake, R.N. Evaluation of Comparative Effects of Arbuscular Mycorrhiza (Rhizophagus intraradices) and Endophyte (Piriformospora indica) Association with Finger Millet (Eleusine coracana) under Drought Stress. Eur. J. Soil Biol. 2017, 81, 1–10. [Google Scholar] [CrossRef]
- Begum, N.; Ahanger, M.A.; Su, Y.; Lei, Y.; Mustafa, N.S.A.; Ahmad, P.; Zhang, L. Improved Drought Tolerance by AMF Inoculation in Maize (Zea mays) Involves Physiological and Biochemical Implications. Plants 2019, 8, 579. [Google Scholar] [CrossRef]
- Krishnamoorthy, R.; Kim, K.; Subramanian, P.; Senthilkumar, M.; Anandham, R.; Sa, T. Arbuscular Mycorrhizal Fungi and Associated Bacteria Isolated from Salt-Affected Soil Enhances the Tolerance of Maize to Salinity in Coastal Reclamation Soil. Agric. Ecosyst. Environ. 2016, 231, 233–239. [Google Scholar] [CrossRef]
- Eroğlu, Ç.G.; Cabral, C.; Ravnskov, S.; Bak Topbjerg, H.; Wollenweber, B. Arbuscular Mycorrhiza Influences Carbon-use Efficiency and Grain Yield of Wheat Grown under Pre- and Post-anthesis Salinity Stress. Plant Biol. 2020, 22, 863–871. [Google Scholar] [CrossRef]
- Norouzinia, F.; Ansari, M.H.; Aminpanah, H.; Firozi, S. Alleviation of Soil Salinity on Physiological and Agronomic Traits of Rice Cultivars Using Arbuscular Mycorrhizal Fungi and Pseudomonas Strains Under Field Conditions. Rev. Agric. Neotrop. 2020, 7, 25–42. [Google Scholar] [CrossRef]
- Wang, G.; Kong, Y.; Liu, Y.; Li, D.; Zhang, X.; Yuan, J.; Li, G. Evolution of Phytotoxicity during the Active Phase of Co-Composting of Chicken Manure, Tobacco Powder and Mushroom Substrate. Waste Manag. 2020, 114, 25–32. [Google Scholar] [CrossRef]
- Ndiate, N.I.; Saeed, Q.; Haider, F.U.; Liqun, C.; Nkoh, J.N.; Mustafa, A. Co-Application of Biochar and Arbuscular Mycorrhizal Fungi Improves Salinity Tolerance, Growth and Lipid Metabolism of Maize (Zea mays L.) in an Alkaline Soil. Plants 2021, 10, 2490. [Google Scholar] [CrossRef]
- Chen, Q.; Deng, X.; Elzenga, J.T.M.; van Elsas, J.D. Effect of Soil Bacteriomes on Mycorrhizal Colonization by Rhizophagus irregularis—Interactive Effects on Maize (Zea mays L.) Growth under Salt Stress. Biol. Fertil. Soils 2022, 58, 515–525. [Google Scholar] [CrossRef]
- Aboul-Nasr, A.; Al-Fayoumy, M.; Hussein, M.; Elhabbab, A. Potential Application of Glomus Intraradices (AMF) and Different Isolates of PGPR (Biotol) to Enhance the Yield and Quality of Wheat Grown in The Field in Calcareous Soil Under Different Salinity Levels. J. Adv. Agric. Res. 2016, 21, 150–181. [Google Scholar] [CrossRef]
- Elgharably, A.; Nafady, N.A. Inoculation with Arbuscular Mycorrhizae, Penicillium funiculosum and Fusarium oxysporum Enhanced Wheat Growth and Nutrient Uptake in the Saline Soil. Rhizosphere 2021, 18, 100345. [Google Scholar] [CrossRef]
- Lee, Y.; Krishnamoorthy, R.; Selvakumar, G.; Kim, K.; Sa, T. Alleviation of Salt Stress in Maize Plant by Co-Inoculation of Arbuscular Mycorrhizal Fungi and Methylobacterium oryzae CBMB20. J. Korean Soc. Appl. Biol. Chem. 2015, 58, 533–540. [Google Scholar] [CrossRef]
- Selvakumar, G.; Shagol, C.C.; Kim, K.; Han, S.; Sa, T. Spore Associated Bacteria Regulates Maize Root K+/Na+ Ion Homeostasis to Promote Salinity Tolerance during Arbuscular Mycorrhizal Symbiosis. BMC Plant Biol. 2018, 18, 109. [Google Scholar] [CrossRef]
- Moreira, H.; Pereira, S.I.A.; Vega, A.; Castro, P.M.L.; Marques, A.P.G.C. Synergistic Effects of Arbuscular Mycorrhizal Fungi and Plant Growth-Promoting Bacteria Benefit Maize Growth under Increasing Soil Salinity. J. Environ. Manag. 2020, 257, 109982. [Google Scholar] [CrossRef]
- Porcel, R.; Aroca, R.; Azcon, R.; Ruiz-Lozano, J.M. Regulation of Cation Transporter Genes by the Arbuscular Mycorrhizal Symbiosis in Rice Plants Subjected to Salinity Suggests Improved Salt Tolerance Due to Reduced Na+ Root-to-Shoot Distribution. Mycorrhiza 2016, 26, 673–684. [Google Scholar] [CrossRef]
- Parvin, S.; Van Geel, M.; Yeasmin, T.; Verbruggen, E.; Honnay, O. Effects of Single and Multiple Species Inocula of Arbuscular Mycorrhizal Fungi on the Salinity Tolerance of a Bangladeshi Rice (Oryza sativa L.) Cultivar. Mycorrhiza 2020, 30, 431–444. [Google Scholar] [CrossRef]
- Tisarum, R.; Theerawitaya, C.; Samphumphuang, T.; Polispitak, K.; Thongpoem, P.; Singh, H.P.; Cha-um, S. Alleviation of Salt Stress in Upland Rice (Oryza sativa L. Ssp. Indica Cv. Leum Pua) Using Arbuscular Mycorrhizal Fungi Inoculation. Front. Plant Sci. 2020, 11, 348. [Google Scholar] [CrossRef]
- Chandra, P.; Singh, A.; Prajapat, K.; Rai, A.K.; Yadav, R.K. Native Arbuscular Mycorrhizal Fungi Improve Growth, Biomass Yield, and Phosphorus Nutrition of Sorghum in Saline and Sodic Soils of the Semi–Arid Region. Environ. Exp. Bot. 2022, 201, 104982. [Google Scholar] [CrossRef]
- Wang, F.; Sun, Y.; Shi, Z. Arbuscular Mycorrhiza Enhances Biomass Production and Salt Tolerance of Sweet Sorghum. Microorganisms 2019, 7, 289. [Google Scholar] [CrossRef]
- Lehmann, J.; Hansel, C.M.; Kaiser, C.; Kleber, M.; Maher, K.; Manzoni, S.; Nunan, N.; Reichstein, M.; Schimel, J.P.; Torn, M.S.; et al. Persistence of Soil Organic Carbon Caused by Functional Complexity. Nat. Geosci. 2020, 13, 529–534. [Google Scholar] [CrossRef]
- Hao, L.; Zhang, Z.; Hao, B.; Diao, F.; Zhang, J.; Bao, Z.; Guo, W. Arbuscular Mycorrhizal Fungi Alter Microbiome Structure of Rhizosphere Soil to Enhance Maize Tolerance to La. Ecotoxicol. Environ. Saf. 2021, 212, 111996. [Google Scholar] [CrossRef]
- Chen, X.W.; Wu, L.; Luo, N.; Mo, C.H.; Wong, M.H.; Li, H. Arbuscular Mycorrhizal Fungi and the Associated Bacterial Community Influence the Uptake of Cadmium in Rice. Geoderma 2019, 337, 749–757. [Google Scholar] [CrossRef]
- Zhan, F.; Li, B.; Jiang, M.; Yue, X.; He, Y.; Xia, Y.; Wang, Y. Arbuscular Mycorrhizal Fungi Enhance Antioxidant Defense in the Leaves and the Retention of Heavy Metals in the Roots of Maize. Environ. Sci. Pollut. Res. 2018, 25, 24338–24347. [Google Scholar] [CrossRef]
- Shi, Z.; Zhang, J.; Lu, S.; Li, Y.; Wang, F. Arbuscular Mycorrhizal Fungi Improve the Performance of Sweet Sorghum Grown in a Mo-Contaminated Soil. J. Fungi 2020, 6, 44. [Google Scholar] [CrossRef]
- Rehman, S.; Mansoora, N.; Al-Dhumri, S.A.; Amjad, S.F.; Al-Shammari, W.B.; Almutari, M.M.; Alhusayni, F.S.; Al Bakre, D.A.; Lalarukh, I.; Alshahri, A.H.; et al. Associative Effects of Activated Carbon Biochar and Arbuscular Mycorrhizal Fungi on Wheat for Reducing Nickel Food Chain Bioavailability. Environ. Technol. Innov. 2022, 26, 102539. [Google Scholar] [CrossRef]
- Li, T.; Sun, Y.; Ruan, Y.; Xu, L.; Hu, Y.; Hao, Z.; Zhang, X.; Li, H.; Wang, Y.; Yang, L.; et al. Potential Role of D-Myo-Inositol-3-Phosphate Synthase and 14-3-3 Genes in the Crosstalk between Zea mays and Rhizophagus intraradices under Drought Stress. Mycorrhiza 2016, 26, 879–893. [Google Scholar] [CrossRef]
- Zhang, Y.; Malzahn, A.A.; Sretenovic, S.; Qi, Y. The Emerging and Uncultivated Potential of CRISPR Technology in Plant Science. Nat. Plants 2019, 5, 778–794. [Google Scholar] [CrossRef]
- Zhang, B.; Zhang, H.; Wang, H.; Wang, P.; Wu, Y.; Wang, M. Effect of Phosphorus Additions and Arbuscular Mycorrhizal Fungal Inoculation on the Growth, Physiology, and Phosphorus Uptake of Wheat Under Two Water Regimes. Commun. Soil Sci. Plant Anal. 2018, 49, 862–874. [Google Scholar] [CrossRef]
- Fileccia, V.; Ingraffia, R.; Amato, G.; Giambalvo, D.; Martinelli, F. Identification of microRNAS Differentially Regulated by Water Deficit in Relation to Mycorrhizal Treatment in Wheat. Mol. Biol. Rep. 2019, 46, 5163–5174. [Google Scholar] [CrossRef]
- Sendek, A.; Karakoç, C.; Wagg, C.; Domínguez-Begines, J.; do Couto, G.M.; van der Heijden, M.G.A.; Naz, A.A.; Lochner, A.; Chatzinotas, A.; Klotz, S.; et al. Drought Modulates Interactions between Arbuscular Mycorrhizal Fungal Diversity and Barley Genotype Diversity. Sci. Rep. 2019, 9, 1–15. [Google Scholar] [CrossRef]
- Fiorilli, V.; Maghrebi, M.; Novero, M.; Votta, C.; Mazzarella, T.; Buffoni, B.; Astolfi, S.; Vigani, G. Arbuscular Mycorrhizal Symbiosis Differentially Affects the Nutritional Status of Two Durum Wheat Genotypes under Drought Conditions. Plants 2022, 11, 804. [Google Scholar] [CrossRef]
- Wang, H.; Liang, L.; Liu, B.; Huang, D.; Liu, S.; Liu, R.; Siddique, K.H.M.; Chen, Y. Arbuscular Mycorrhizas Regulate Photosynthetic Capacity and Antioxidant Defense Systems to Mediate Salt Tolerance in Maize. Plants 2020, 9, 1430. [Google Scholar] [CrossRef] [PubMed]
- Sadia, K.; Asma, B.; Riffat, N.M. Role of Arbuscular Mycorrhizal Fungi in Phytoremediation of Heavy Metals and Effects on Growth and Biochemical Activities of Wheat (Triticum aestivum L.) Plants in Zn Contaminated Soils. Afr. J. Biotechnol. 2016, 15, 872–883. [Google Scholar] [CrossRef]
- Mustafa, G.; Randoux, B.; Tisserant, B.; Fontaine, J.; Magnin-Robert, M.; Lounès-Hadj Sahraoui, A.; Reignault, P. Phosphorus Supply, Arbuscular Mycorrhizal Fungal Species, and Plant Genotype Impact on the Protective Efficacy of Mycorrhizal Inoculation against Wheat Powdery Mildew. Mycorrhiza 2016, 26, 685–697. [Google Scholar] [CrossRef]
- Mustafa, G.; Khong, N.G.; Tisserant, B.; Randoux, B.; Fontaine, J.; Magnin-Robert, M.; Reignault, P.; Sahraoui, A.L.H. Defence Mechanisms Associated with Mycorrhiza-Induced Resistance in Wheat against Powdery Mildew. Funct. Plant Biol. 2017, 44, 443–454. [Google Scholar] [CrossRef] [PubMed]
- Fiorilli, V.; Vannini, C.; Ortolani, F.; Garcia-Seco, D.; Chiapello, M.; Novero, M.; Domingo, G.; Terzi, V.; Morcia, C.; Bagnaresi, P.; et al. Omics Approaches Revealed How Arbuscular Mycorrhizal Symbiosis Enhances Yield and Resistance to Leaf Pathogen in Wheat. Sci. Rep. 2018, 8, 9625. [Google Scholar] [CrossRef] [PubMed]
- Vannini, C.; Domingo, G.; Fiorilli, V.; Seco, D.G.; Novero, M.; Marsoni, M.; Wisniewski-Dye, F.; Bracale, M.; Moulin, L.; Bonfante, P. Proteomic Analysis Reveals How Pairing of a Mycorrhizal Fungus with Plant Growth-Promoting Bacteria Modulates Growth and Defense in Wheat. Plant Cell Environ. 2021, 44, 1946–1960. [Google Scholar] [CrossRef]
- Imperiali, N.; Chiriboga, X.; Schlaeppi, K.; Fesselet, M.; Villacrés, D.; Jaffuel, G.; Bender, S.F.; Dennert, F.; Blanco-Pérez, R.; van der Heijden, M.G.A.; et al. Combined Field Inoculations of Pseudomonas Bacteria, Arbuscular Mycorrhizal Fungi, and Entomopathogenic Nematodes and Their Effects on Wheat Performance. Front. Plant Sci. 2017, 8, 1809. [Google Scholar] [CrossRef]
- Pons, C.; Voß, A.C.; Schweiger, R.; Müller, C. Effects of Drought and Mycorrhiza on Wheat and Aphid Infestation. Ecol. Evol. 2020, 10, 10481–10491. [Google Scholar] [CrossRef]
- Spagnoletti, F.N.; Carmona, M.; Balestrasse, K.; Chiocchio, V.; Giacometti, R.; Lavado, R.S. The Arbuscular Mycorrhizal Fungus Rhizophagus intraradices Reduces the Root Rot Caused by Fusarium pseudograminearum in Wheat. Rhizosphere 2021, 19, 100369. [Google Scholar] [CrossRef]
- Campo, S.; Martín-Cardoso, H.; Olivé, M.; Pla, E.; Catala-Forner, M.; Martínez-Eixarch, M.; San Segundo, B. Effect of Root Colonization by Arbuscular Mycorrhizal Fungi on Growth, Productivity and Blast Resistance in Rice. Rice 2020, 13, 42. [Google Scholar] [CrossRef] [PubMed]
- Prasetyo, J.; Ginting, C.; Akin, H.M.; Suharjo, R.; Niswati, A.; Afandi, A.; Adiwijaya, R.; Sudiono, S.; Nurdin, M. The Effect of Biological Agent and Botanical Fungicides on Maize Downy Mildew. Biodiversitas 2021, 22, 1652–1657. [Google Scholar] [CrossRef]
- Tian, L.; Chang, C.; Ma, L.; Nasir, F.; Zhang, J.; Li, W.; Tran, L.-S.P.; Tian, C. Comparative Study of the Mycorrhizal Root Transcriptomes of Wild and Cultivated Rice in Response to the Pathogen Magnaporthe oryzae. Rice 2019, 12, 35. [Google Scholar] [CrossRef]
- Bhavanam, S.; Stout, M.J. Assessment of Silicon-and Mycorrhizae-Mediated Constitutive and Induced Systemic Resistance in Rice, Oryza sativa L., against the Fall Armyworm, Spodoptera frugiperda Smith. Plants 2021, 10, 2126. [Google Scholar] [CrossRef]
- Charters, M.D.; Durant, E.K.; Sait, S.M.; Field, K.J. Impacts of Aphid Herbivory on Mycorrhizal Growth Responses across Three Cultivars of Wheat. Plants People Planet 2022, 4, 655–666. [Google Scholar] [CrossRef]
- Liu, Z.; Li, Y.; Ma, L.; Wei, H.; Zhang, J.; He, X.; Tian, C. Coordinated Regulation of Arbuscular Mycorrhizal Fungi and Soybean MAPK Pathway Genes Improved Mycorrhizal Soybean Drought Tolerance. Mol. Plant-Microbe Interact. 2015, 28, 408–419. [Google Scholar] [CrossRef]
- Igiehon, O.N.; Babalola, O.O. Rhizobium and Mycorrhizal Fungal Species Improved Soybean Yield Under Drought Stress Conditions. Curr. Microbiol. 2021, 78, 1615–1627. [Google Scholar] [CrossRef]
- Sheteiwy, M.S.; Abd Elgawad, H.; Xiong, Y.C.; Macovei, A.; Brestic, M.; Skalicky, M.; Shaghaleh, H.; Alhaj Hamoud, Y.; El-Sawah, A.M. Inoculation with Bacillus amyloliquefaciens and Mycorrhiza Confers Tolerance to Drought Stress and Improve Seed Yield and Quality of Soybean Plant. Physiol. Plant. 2021, 172, 2153–2169. [Google Scholar] [CrossRef] [PubMed]
- Al-Karaki, G.N.; Williams, M. Mycorrhizal Mixtures Affect the Growth, Nutrition, and Physiological Responses of Soybean to Water Deficit. Acta Physiol. Plant. 2021, 43, 75. [Google Scholar] [CrossRef]
- Askari, A.; Ardakani, M.R.; Vazan, S.; Paknejad, F.; Hosseini, Y. The Effect of Mycorrhizal Symbiosis and Seed Priming on the Amount of Chlorophyll Index and Absorption of Nutrients under Drought Stress in Sesame Plant under Field Conditions. Appl. Ecol. Environ. Res. 2018, 16, 335–357. [Google Scholar] [CrossRef]
- Gholinezhad, E.; Darvishzadeh, R. Field Crops Research Influence of Arbuscular Mycorrhiza Fungi and Drought Stress on Fatty Acids Profile of Sesame (Sesamum indicum L.). Field Crops Res. 2021, 262, 108035. [Google Scholar] [CrossRef]
- Rahimzadeh, S.; Pirzad, A. Microorganisms (AMF and PSB) Interaction on Linseed Productivity under Water-Deficit Condition. Int. J. Plant Prod. 2017, 11, 259–274. [Google Scholar] [CrossRef]
- Ansari, A.; Razmjoo, J.; Zarei, M.; Karimmojeni, H. Salicylic Acid Affects Mycorrhizal Features, Antioxidant Enzyme Activities and Seed Yield of Linseed under Water—Deficit Stress in Open—Field Conditions. Biol. Futur. 2021, 72, 211–227. [Google Scholar] [CrossRef]
- Oliveira, T.C.; Cabral, J.S.R.; Santana, L.R.; Tavares, G.G.; Santos, L.D.S.; Paim, T.P.; Müller, C.; Silva, F.G.; Costa, A.C.; Souchie, E.L.; et al. The Arbuscular Mycorrhizal Fungus Rhizophagus clarus Improves Physiological Tolerance to Drought Stress in Soybean Plants. Sci. Rep. 2022, 12, 9044. [Google Scholar] [CrossRef]
- Ghasemi, N.; Gholamhoseini, M.; Bazrafshan, F.; Habibzadeh, F. Yield, Irrigation Water Productivity and Nutrient Uptake of Arbuscular Mycorrhiza Inoculated Sesame under Drought Stress Conditions. Agric. Water Manag. 2022, 266, 107569. [Google Scholar] [CrossRef]
- Pereira, S.I.A.; Moreira, H.; Argyras, K.; Castro, P.M.L.; Marques, A.P.G.C. Promotion of Sunflower Growth under Saline Water Irrigation by the Inoculation of Beneficial Microorganisms. Appl. Soil Ecol. 2016, 105, 36–47. [Google Scholar] [CrossRef]
- Ghouchani, R.; Abbaspour, H.; Saed-Moucheshi, A.; Pessarakli, M. Colonization with Endo-Mycorrhiza Affects the Resistance of Safflower in Response to Salinity Condition. J. Plant Nutr. 2017, 40, 1856–1867. [Google Scholar] [CrossRef]
- Heydari, S.; Pirzad, A. Efficiency of Funneliformis mosseae and Thiobacillus Sp. on the Secondary Metabolites (Essential Oil, Seed Oil and Mucilage) of Lallemantia iberica under Salinity Stress. J. Hortic. Sci. Biotechnol. 2021, 96, 249–259. [Google Scholar] [CrossRef]
- Qin, W.; Yan, H.; Zou, B.; Guo, R.; Ci, D.; Tang, Z.; Zou, X.; Zhang, X.; Yu, X.; Wang, Y.; et al. Arbuscular Mycorrhizal Fungi Alleviate Salinity Stress in Peanut: Evidence from Pot-Grown and Field Experiments. Food Energy Secur. 2021, 10, e314. [Google Scholar] [CrossRef]
- Hashem, A.; Abd_Allah, E.F.; Alqarawi, A.A.; Wirth, S.; Egamberdieva, D. Comparing Symbiotic Performance and Physiological Responses of Two Soybean Cultivars to Arbuscular Mycorrhizal Fungi under Salt Stress. Saudi J. Biol. Sci. 2019, 26, 38–48. [Google Scholar] [CrossRef] [PubMed]
- Păun, A.; Neagoe, A.; Păun, M.; Baciu, I.; Iordache, V. Heavy Metal-Induced Differential Responses to Oxidative Stress and Protection by Mycorrhization in Sunflowers Grown in Lab and Field Scales. Pol. J. Environ. Stud. 2015, 24, 1235–1247. [Google Scholar] [CrossRef]
- Molina, A.S.; Lugo, M.A.; Pérez Chaca, M.V.; Vargas-Gil, S.; Zirulnik, F.; Leporati, J.; Ferrol, N.; Azcón-Aguilar, C. Effect of Arbuscular Mycorrhizal Colonization on Cadmium-Mediated Oxidative Stress in Glycine max (L.) Merr. Plants 2020, 9, 108. [Google Scholar] [CrossRef]
- Adeyemi, N.O.; Atayese, M.O.; Sakariyawo, O.S.; Azeez, J.O.; Abayomi Sobowale, S.P.; Olubode, A.; Mudathir, R.; Adebayo, R.; Adeoye, S. Alleviation of Heavy Metal Stress by Arbuscular Mycorrhizal Symbiosis in Glycine max L. Grown in Copper, Lead and Zinc Contaminated Soils. Rhizosphere 2021, 18, 100325. [Google Scholar] [CrossRef]
- Sheteiwy, M.S.; Ali, D.F.I.; Xiong, Y.C.; Brestic, M.; Skalicky, M.; Hamoud, Y.A.; Ulhassan, Z.; Shaghaleh, H.; AbdElgawad, H.; Farooq, M.; et al. Physiological and Biochemical Responses of Soybean Plants Inoculated with Arbuscular Mycorrhizal Fungi and Bradyrhizobium under Drought Stress. BMC Plant Biol. 2021, 21, 195. [Google Scholar] [CrossRef]
- Hoang, D.T.T.; Rashtbari, M.; Anh, L.T.; Wang, S.; Tu, D.T.; Hiep, N.V.; Razavi, B.S. Mutualistic Interaction between Arbuscular Mycorrhiza Fungi and Soybean Roots Enhances Drought Resistant through Regulating Glucose Exudation and Rhizosphere Expansion. Soil Biol. Biochem. 2022, 171, 108728. [Google Scholar] [CrossRef]
- Spagnoletti, F.N.; Cornero, M.; Chiocchio, V.; Lavado, R.S.; Roberts, I.N. Arbuscular Mycorrhiza Protects Soybean Plants against Macrophomina phaseolina Even under Nitrogen Fertilization. Eur. J. Plant Pathol. 2020, 156, 839–849. [Google Scholar] [CrossRef]
- Marquez, N.; Giachero, M.L.; Gallou, A.; Debat, H.J.; Declerck, S.; Ducasse, D.A. Transcriptome Analysis of Mycorrhizal and Nonmycorrhizal Soybean Plantlets upon Infection with Fusarium virguliforme, One Causal Agent of Sudden Death Syndrome. Plant Pathol. 2019, 68, 470–480. [Google Scholar] [CrossRef]
- Pawlowski, M.L.; Hartman, G.L. Reduction of Sudden Death Syndrome Foliar Symptoms and Fusarium virguliforme DNA in Roots Inoculated with Rhizophagus intraradices. Plant Dis. 2020, 104, 1415–1420. [Google Scholar] [CrossRef] [PubMed]
- Benedetti, T.; Antoniolli, Z.I.; Sordi, E.; Carvalho, I.R.; Bortoluzzi, E.C. Use of the Glomus etunicatum as Biocontrol Agent of the Soybean Cyst Nematode. Res., Soc. Dev. 2021, 10, e7310615132. [Google Scholar] [CrossRef]
- Dabré, É.E.; Hijri, M.; Favret, C. Influence on Soybean Aphid by the Tripartite Interaction between Soybean, a Rhizobium Bacterium, and an Arbuscular Mycorrhizal Fungus. Microorganisms 2022, 10, 1196. [Google Scholar] [CrossRef] [PubMed]
- Spagnoletti, F.; Carmona, M.; Gómez, N.E.T.; Chiocchio, V.; Lavado, R.S. Arbuscular Mycorrhiza Reduces the Negative Effects of M. Phaseolina on Soybean Plants in Arsenic-Contaminated Soils. Appl. Soil Ecol. 2017, 121, 41–47. [Google Scholar] [CrossRef]
- Bán, R.; Baglyas, G.; Virányi, F.; Barna, B.; Posta, K.; Kiss, J.; Körösi, K. The Chemical Inducer, BTH (Benzothiadiazole) and Root Colonization by Mycorrhizal Fungi (Glomus Spp.) Trigger Resistance against White Rot (Sclerotinia sclerotiorum) in Sunflower. Acta Biol. Hung. 2017, 68, 50–59. [Google Scholar] [CrossRef] [PubMed]
- Marquez, N.; Giachero, M.L.; Gallou, A.; Debat, H.J.; Cranenbrouck, S.; Di Rienzo, J.A.; Pozo, M.J.; Ducasse, D.A.; Declerck, S. Transcriptional Changes in Mycorrhizal and Nonmycorrhizal Soybean Plants upon Infection with the Fungal Pathogen Macrophomina phaseolina. Mol. Plant-Microbe Interact. 2018, 31, 842–855. [Google Scholar] [CrossRef]
- Quiroga, G.; Erice, G.; Ding, L.; Chaumont, F.; Aroca, R.; Ruiz-Lozano, J.M. The Arbuscular Mycorrhizal Symbiosis Regulates Aquaporins Activity and Improves Root Cell Water Permeability in Maize Plants Subjected to Water Stress. Plant Cell Environ. 2019, 42, 2274–2290. [Google Scholar] [CrossRef]
- Li, T.; Hu, Y.J.; Hao, Z.P.; Li, H.; Wang, Y.S.; Chen, B.D. First Cloning and Characterization of Two Functional Aquaporin Genes from an Arbuscular Mycorrhizal Fungus Glomus intraradices. New Phytol. 2013, 197, 617–630. [Google Scholar] [CrossRef] [PubMed]
- Quiroga, G.; Erice, G.; Aroca, R.; Chaumont, F.; Ruiz-Lozano, J.M. Contribution of the Arbuscular Mycorrhizal Symbiosis to the Regulation of Radial Root Water Transport in Maize Plants under Water Deficit. Environ. Exp. Bot. 2019, 167, 103821. [Google Scholar] [CrossRef]
- Quiroga, G.; Erice, G.; Aroca, R.; Delgado-Huertas, A.; Ruiz-Lozano, J.M. Elucidating the Possible Involvement of Maize Aquaporins and Arbuscular Mycorrhizal Symbiosis in the Plant Ammonium and Urea Transport under Drought Stress Conditions. Plants 2020, 9, 148. [Google Scholar] [CrossRef] [PubMed]
- Gong, M.; Bai, N.; Wang, P.; Su, J.; Chang, Q.; Zhang, Q. Co-Inoculation with Arbuscular Mycorrhizal Fungi and Dark Septate Endophytes under Drought Stress: Synergistic or Competitive Effects on Maize Growth, Photosynthesis, Root Hydraulic Properties and Aquaporins? Plants 2023, 12, 2596. [Google Scholar] [CrossRef]
- Asadollahi, M.; Iranbakhsh, A.; Ahmadvand, R.; Ebadi, M.; Mehregan, I. Synergetic Effect of Water Deficit and Arbuscular Mycorrhizal Symbiosis on the Expression of Aquaporins in Wheat (Triticum aestivum L.) Roots: Insights from NGS RNA-Sequencing. Physiol. Mol. Biol. Plants 2023, 29, 195–208. [Google Scholar] [CrossRef]
- Wipf, D.; Krajinski, F.; van Tuinen, D.; Recorbet, G.; Courty, P.E. Trading on the Arbuscular Mycorrhiza Market: From Arbuscules to Common Mycorrhizal Networks. New Phytol. 2019, 223, 1127–1142. [Google Scholar] [CrossRef]
- Calabrese, S.; Cusant, L.; Sarazin, A.; Niehl, A.; Erban, A.; Brulé, D.; Recorbet, G.; Wipf, D.; Roux, C.; Kopka, J.; et al. Imbalanced Regulation of Fungal Nutrient Transports According to Phosphate Availability in a Symbiocosm Formed by Poplar, Sorghum, and Rhizophagus irregularis. Front. Plant Sci. 2019, 10, 1617. [Google Scholar] [CrossRef]
- Chen, Z.; Zhou, M.; Newman, I.A.; Mendham, N.J.; Zhang, G.; Shabala, S. Potassium and Sodium Relations in Salinised Barley Tissues as a Basis of Differential Salt Tolerance. Funct. Plant Biol. 2007, 34, 150–162. [Google Scholar] [CrossRef]
- Estrada, B.; Aroca, R.; Maathuis, F.J.M.; Barea, J.M.; Ruiz-Lozano, J.M. Arbuscular Mycorrhizal Fungi Native from a Mediterranean Saline Area Enhance Maize Tolerance to Salinity through Improved Ion Homeostasis. Plant Cell Environ. 2013, 36, 1771–1782. [Google Scholar] [CrossRef] [PubMed]
- Wang, H.; An, T.; Huang, D.; Liu, R.; Xu, B.; Zhang, S.; Deng, X.; Siddique, K.H.M.; Chen, Y. Arbuscular Mycorrhizal Symbioses Alleviating Salt Stress in Maize Is Associated with a Decline in Root-to-Leaf Gradient of Na+/K+ Ratio. BMC Plant Biol. 2021, 21, 457. [Google Scholar] [CrossRef]
- Liu, F.; Xu, Y.; Han, G.; Wang, W.; Li, X.; Cheng, B. Identification and Functional Characterization of a Maize Phosphate Transporter Induced by Mycorrhiza Formation. Plant Cell Physiol. 2018, 59, 1683–1694. [Google Scholar] [CrossRef] [PubMed]
- Ferrol, N.; Azcón-Aguilar, C.; Pérez-Tienda, J. Review: Arbuscular Mycorrhizas as Key Players in Sustainable Plant Phosphorus Acquisition: An Overview on the Mechanisms Involved. Plant Sci. 2019, 280, 441–447. [Google Scholar] [CrossRef] [PubMed]
- Yang, S.Y.; Grønlund, M.; Jakobsen, I.; Grotemeyer, M.S.; Rentsch, D.; Miyao, A.; Hirochika, H.; Kumar, C.S.; Sundaresan, V.; Salamin, N.; et al. Nonredundant Regulation of Rice Arbuscular Mycorrhizal Symbiosis by Two Members of the PHOSPHATE TRANSPORTER1 Gene Family. Plant Cell 2012, 24, 4236–4251. [Google Scholar] [CrossRef]
- Liu, F.; Xu, Y.; Jiang, H.; Jiang, C.; Du, Y.; Gong, C.; Wang, W.; Zhu, S.; Han, G.; Cheng, B. Systematic Identification, Evolution and Expression Analysis of the Zea mays PHT1 Gene Family Reveals Several New Members Involved in Root Colonization by Arbuscular Mycorrhizal Fungi. Int. J. Mol. Sci. 2016, 17, 930. [Google Scholar] [CrossRef] [PubMed]
- Xie, X.; Huang, W.; Liu, F.; Tang, N.; Liu, Y.; Lin, H.; Zhao, B. Functional Analysis of the Novel Mycorrhiza-Specific Phosphate Transporter AsPT1 and PHT1 Family from Astragalus sinicus during the Arbuscular Mycorrhizal Symbiosis. New Phytol. 2013, 198, 836–852. [Google Scholar] [CrossRef] [PubMed]
- Wang, D.; Dong, W.; Murray, J.; Wang, E. Innovation and Appropriation in Mycorrhizal and Rhizobial Symbioses. Plant Cell 2022, 34, 1573–1599. [Google Scholar] [CrossRef] [PubMed]
- Walder, F.; Brulé, D.; Koegel, S.; Wiemken, A.; Boller, T.; Courty, P.E. Plant Phosphorus Acquisition in a Common Mycorrhizal Network: Regulation of Phosphate Transporter Genes of the Pht1 Family in Sorghum and Flax. New Phytol. 2015, 205, 1632–1645. [Google Scholar] [CrossRef]
- Liu, L.; Li, J.; Yue, F.; Yan, X.; Wang, F.; Bloszies, S.; Wang, Y. Effects of arbuscular mycorrhizal inoculation and biochar amendment on maize growth, cadmium uptake and soil cadmium speciation in Cd-contaminated soil. Chemosphere 2018, 194, 495–503. [Google Scholar] [CrossRef]
- Gu, L.; Zhao, M.; Ge, M.; Zhu, S.; Cheng, B.; Li, X. Transcriptome Analysis Reveals Comprehensive Responses to Cadmium Stress in Maize Inoculated with Arbuscular Mycorrhizal Fungi. Ecotoxicol. Environ. Saf. 2019, 186, 109744. [Google Scholar] [CrossRef]
- Zhang, B.; Shi, F.; Zheng, X.; Pan, H.; Wen, Y.; Song, F. Effects of AMF Compound Inoculants on Growth, Ion Homeostasis, and Salt Tolerance-Related Gene Expression in Oryza sativa L. Under Salt Treatments. Rice 2023, 16, 18. [Google Scholar] [CrossRef]
- Olalde-Portugal, V.; Cabrera-Ponce, J.L.; Gastelum-Arellanez, A.; Guerrero-Rangel, A.; Winkler, R.; Valdés-Rodríguez, S. Proteomic Analysis and Interactions Network in Leaves of Mycorrhizal and Nonmycorrhizal Sorghum Plants under Water Deficit. PeerJ 2020, 8, e8991. [Google Scholar] [CrossRef]
- Puccio, G.; Ingraffia, R.; Mercati, F.; Amato, G.; Giambalvo, D.; Martinelli, F.; Sunseri, F.; Frenda, A.S. Transcriptome Changes Induced by Arbuscular Mycorrhizal Symbiosis in Leaves of Durum Wheat (Triticum durum Desf.) Promote Higher Salt Tolerance. Sci. Rep. 2023, 13, 116. [Google Scholar] [CrossRef]
- El-Sawah, A.M.; Abdel-Fattah, G.G.; Holford, P.; Korany, S.M.; Alsherif, E.A.; AbdElgawad, H.; Ulhassan, Z.; Jośko, I.; Ali, B.; Sheteiwy, M.S. Funneliformis constrictum Modulates Polyamine Metabolism to Enhance Tolerance of Zea mays L. to Salinity. Microbiol. Res. 2023, 266, 127254. [Google Scholar] [CrossRef]
- Li, S.; Yang, W.; Guo, J.; Li, X.; Lin, J.; Zhu, X. Changes in Photosynthesis and Respiratory Metabolism of Maize Seedlings Growing under Low Temperature Stress May Be Regulated by Arbuscular Mycorrhizal Fungi. Plant Physiol. Biochem. 2020, 154, 1–10. [Google Scholar] [CrossRef] [PubMed]
- Kosakivska, I.V.; Vedenicheva, N.P.; Babenko, L.M.; Voytenko, L.V.; Romanenko, K.O.; Vasyuk, V.A. Exogenous Phytohormones in the Regulation of Growth and Development of Cereals under Abiotic Stresses. Mol. Biol. Rep. 2022, 49, 617–628. [Google Scholar] [CrossRef]
- Ge, J.; Li, D.; Ding, J.; Xiao, X.; Liang, Y. Microbial Coexistence in the Rhizosphere and the Promotion of Plant Stress Resistance: A Review. Environ. Res. 2023, 222, 115298. [Google Scholar] [CrossRef] [PubMed]
- Jung, S.C.; Martinez-Medina, A.; Lopez-Raez, J.A.; Pozo, M.J. Mycorrhiza-Induced Resistance and Priming of Plant Defenses. J. Chem. Ecol. 2012, 38, 651–664. [Google Scholar] [CrossRef] [PubMed]
- Zipfel, C. Pattern-Recognition Receptors in Plant Innate Immunity. Curr. Opin. Immunol. 2008, 20, 10–16. [Google Scholar] [CrossRef] [PubMed]
- Boller, T.; Felix, G. A Renaissance of Elicitors: Perception of Microbe-Associated Molecular Patterns and Danger Signals by Pattern-Recognition Receptors. Annu. Rev. Plant Biol. 2009, 60, 379–406. [Google Scholar] [CrossRef]
- Hohmann, P.; Messmer, M.M. Breeding for Mycorrhizal Symbiosis: Focus on Disease Resistance. Euphytica 2017, 213, 113. [Google Scholar] [CrossRef]
- Li, M.; Wang, R.; Tian, H.; Gao, Y. Transcriptome Responses in Wheat Roots to Colonization by the Arbuscular Mycorrhizal Fungus Rhizophagus irregularis. Mycorrhiza 2018, 28, 747–759. [Google Scholar] [CrossRef]
- Campos-Soriano, L.; García-Martínez, J.; Segundo, B.S. The Arbuscular Mycorrhizal Symbiosis Promotes the Systemic Induction of Regulatory Defence-related Genes in Rice Leaves and Confers Resistance to Pathogen Infection. Mol. Plant Pathol. 2012, 13, 579–592. [Google Scholar] [CrossRef]
- Sousa, T.P.D.; Chaibub, A.A.; Silva, G.B.D.; Filippi, M.C.C.D. Trichoderma asperellum Modulates Defense Genes and Potentiates Gas Exchanges in Upland Rice Plants. Physiol. Mol. Plant Pathol. 2020, 112, 101561. [Google Scholar] [CrossRef]
- El-Sharkawy, H.H.A.; Rashad, Y.M.; Elazab, N.T. Induction of Multiple Defense Responses in Wheat Plants against Stripe Rust Using Mycorrhizal Fungi and HH1. BioControl 2023, 68, 525–535. [Google Scholar] [CrossRef]
- Anjali; Kumar, S.; Korra, T.; Thakur, R.; Arutselvan, R.; Kashyap, A.S.; Nehela, Y.; Chaplygin, V.; Minkina, T.; Keswani, C. Role of Plant Secondary Metabolites in Defence and Transcriptional Regulation in Response to Biotic Stress. Plant Stress 2023, 8, 100154. [Google Scholar] [CrossRef]
- Rashad, Y.M.; El-Sharkawy, H.H.; Abdalla, S.A.; Ibrahim, O.M.; Elazab, N.T. Mycorrhizal Colonization and Streptomyces viridosporus HH1 Synergistically Up-Regulate the Polyphenol Biosynthesis Genes in Wheat against Stripe Rust. BMC Plant Biol. 2023, 23, 388. [Google Scholar] [CrossRef]
- Recchia, G.H.; Konzen, E.R.; Cassieri, F.; Caldas, D.G.G.; Tsai, S.M. Arbuscular Mycorrhizal Symbiosis Leads to Differential Regulation of Drought-Responsive Genes in Tissue-Specific Root Cells of Common Bean. Front. Microbiol. 2018, 9, 1339. [Google Scholar] [CrossRef]
- Vangelisti, A.; Natali, L.; Bernardi, R.; Sbrana, C.; Hassani-pak, K.; Hughes, D.; Cavallini, A.; Giovannetti, M. Transcriptome Changes Induced by Arbuscular Mycorrhizal Fungi in Sunflower (Helianthus annuus L.) Roots. Sci. Rep. 2018, 8, 4. [Google Scholar] [CrossRef]
- Siddique, I.; Shah, T.; Ali, A.; Ahmad, I.; Roberto, D.; Munsif, F. Arbuscular Mycorrhizal Fungi and Biofilm Forming Bacteria Act Synergistically to Modulate Proline Metabolism, Antioxidant Defense System and Aquaporin Genes Expression Under Drought Stress. Preprints 2022, 2022080133. [Google Scholar] [CrossRef]
- Singh, R.K.; Shweta, S.; Muthamilarasan, M.; Rani, R.; Prasad, M. Study on Aquaporins of Setaria italica Suggests the Involvement of SiPIP3;1 and SiSIP1;1 in Abiotic Stress Response. Funct. Integr. Genom. 2019, 19, 587–596. [Google Scholar] [CrossRef]
- Kabir, A.H.; Debnath, T.; Das, U.; Prity, S.A.; Haque, A.; Rahman, M.M.; Parvez, M.S. Arbuscular Mycorrhizal Fungi Alleviate Fe-Deficiency Symptoms in Sunflower by Increasing Iron Uptake and Its Availability along with Antioxidant Defense. Plant Physiol. Biochem. 2020, 150, 254–262. [Google Scholar] [CrossRef]
- Liao, Q.; Zhou, T.; Yao, J.; Han, Q.; Song, H.; Guan, C.; Hua, Y.; Zhang, Z. Genome-Scale Characterization of the Vacuole Nitrate Transporter Chloride Channel (CLC) Genes and Their Transcriptional Responses to Diverse Nutrient Stresses in Allotetraploid Rapeseed. PLoS ONE 2018, 13, e0208648. [Google Scholar] [CrossRef]
- Abbaspour, H.; Pour, F.S.N.; Abdel-Wahhab, M.A. Arbuscular Mycorrhizal Symbiosis Regulates the Physiological Responses, Ion Distribution and Relevant Gene Expression to Trigger Salt Stress Tolerance in Pistachio. Physiol. Mol. Biol. Plants 2021, 27, 1765–1778. [Google Scholar] [CrossRef]
- Salloum, M.S.; Insani, M.; Monteoliva, M.I.; Menduni, M.F.; Silvente, S.; Carrari, F.; Luna, C. Metabolic Responses to Arbuscular Mycorrhizal Fungi Are Shifted in Roots of Contrasting Soybean Genotypes. Mycorrhiza 2019, 29, 459–473. [Google Scholar] [CrossRef]
- You, J.; Zhang, Y.; Liu, A.; Li, D.; Wang, X.; Dossa, K.; Zhou, R.; Yu, J.; Zhang, Y.; Wang, L.; et al. Transcriptomic and Metabolomic Profiling of Drought-Tolerant and Susceptible Sesame Genotypes in Response to Drought Stress. BMC Plant Biol. 2019, 19, 267. [Google Scholar] [CrossRef]
- Shimoia, E.P.; Da-Silva, C.J.; Posso, D.A.; Martins, T.d.S.; Agualongo, D.A.P.; de Oliveira, A.C.B.; Amarante, L.D. Co-inoculation of Seeds with Bradyrhizobium, Azospirillum, and Rhizophagus Improves Nitrogen Assimilation and Growth in Soybean Plants Subjected to Waterlogging. Russ. J. Plant Physiol. 2023, 70, 146. [Google Scholar] [CrossRef]
- Zhao, M.-J.; Yin, L.-J.; Liu, Y.; Ma, J.; Zheng, J.-C.; Lan, J.-H.; Fu, J.-D.; Chen, M.; Xu, Z.-S.; Ma, Y.-Z. The ABA-Induced Soybean ERF Transcription Factor Gene GmERF75 Plays a Role in Enhancing Osmotic Stress Tolerance in Arabidopsis and Soybean. BMC Plant Biol. 2019, 19, 506. [Google Scholar] [CrossRef]
- Spagnoletti, F.N.; Leiva, M.; Chiocchio, V.; Lavado, R.S. Phosphorus Fertilization Reduces the Severity of Charcoal Rot (Macrophomina phaseolina) and the Arbuscular Mycorrhizal Protection in Soybean. J. Plant Nutr. Soil Sci. 2018, 181, 855–860. [Google Scholar] [CrossRef]
- Slimani, A.; Oufdou, K.; Meddich, A. Intercropping with Alfalfa and Co-Inoculation of AMF and PGPR Improve Growth, Yield, Grain Bioactive Quality, and Soil Fertility of Barley. Arch. Agron. Soil Sci. 2023, 69, 3469–3483. [Google Scholar] [CrossRef]
- Nafady, N.A.; Hashem, M.; Hassan, E.A.; Ahmed, H.A.M.; Alamri, S.A. The Combined Effect of Arbuscular Mycorrhizae and Plant-Growth-Promoting Yeast Improves Sunflower Defense against Macrophomina phaseolina Diseases. Biol. Control 2019, 138, 104049. [Google Scholar] [CrossRef]
- Elewa, I.S.; Mostafa, M.H.; Sahab, A.F.; Ziedan, E.H. Direct Effect of Biocontrol Agents on Wilt and Root-Rot Diseases of Sesame. Arch. Phytopathol. Plant Prot. 2011, 44, 493–504. [Google Scholar] [CrossRef]
- Rashad, Y.; Aseel, D.; Hammad, S.; Elkelish, A. Rhizophagus irregularis and Rhizoctonia solani Differentially Elicit Systemic Transcriptional Expression of Polyphenol Biosynthetic Pathways Genes in Sunflower. Biomolecules 2020, 10, 379. [Google Scholar] [CrossRef]
- Ahmed, W.K.; Alsalim, H.A.A.; Mohammed, A.T.; Youssef, H.M. Evaluation of the Effectiveness of Some Mycorrhizal Fungi Isolates against Charcoal Rot Disease. Egypt. J. Biol. Pest Control 2023, 33, 104. [Google Scholar] [CrossRef]
- Malik, R.J.; Dixon, M.H.; Bever, J.D. Mycorrhizal Composition Can Predict Foliar Pathogen Colonization in Soybean. Biol. Control. 2016, 103, 46–53. [Google Scholar] [CrossRef]
- Gao, X.; Lu, X.; Wu, M.; Zhang, H.; Pan, R.; Tian, J.; Li, S.; Liao, H. Co-Inoculation with Rhizobia and AMF Inhibited Soybean Red Crown Rot: From Field Study to Plant Defense-Related Gene Expression Analysis. PLoS ONE 2012, 7, e33977. [Google Scholar] [CrossRef]
- Aquino, B.; Bradley, J.M.; Lumba, S. On the Outside Looking in: Roles of Endogenous and Exogenous Strigolactones. Plant J. 2021, 105, 322–334. [Google Scholar] [CrossRef]
- Clark, J.; Bennett, T. Cracking the Enigma: Understanding Strigolactone Signalling in the Rhizosphere. J. Exp. Bot. 2023, 75, 1159–1173. [Google Scholar] [CrossRef]
- Weng, W.; Yan, J.; Zhou, M.; Yao, X.; Gao, A.; Ma, C.; Cheng, J.; Ruan, J. Roles of Arbuscular Mycorrhizal Fungi as a Biocontrol Agent in the Control of Plant Diseases. Microorganisms 2022, 10, 1266. [Google Scholar] [CrossRef]
- Liu, F.; Rice, J.H.; Lopes, V.; Grewal, P.; Lebeis, S.L.; Hewezi, T.; Staton, M.E. Overexpression of Strigolactone-Associated Genes Exerts Fine-Tuning Selection on Soybean Rhizosphere Bacterial and Fungal Microbiome. Phytobiomes J. 2020, 4, 239–251. [Google Scholar] [CrossRef]
- Belmondo, S.; Marschall, R.; Tudzynski, P.; López Ráez, J.A.; Artuso, E.; Prandi, C.; Lanfranco, L. Identification of Genes Involved in Fungal Responses to Strigolactones Using Mutants from Fungal Pathogens. Curr. Genet. 2017, 63, 201–213. [Google Scholar] [CrossRef]
- Dor, E.; Joel, D.M.; Kapulnik, Y.; Koltai, H.; Hershenhorn, J. The Synthetic Strigolactone GR24 Influences the Growth Pattern of Phytopathogenic Fungi. Planta 2011, 234, 419–427. [Google Scholar] [CrossRef]
- Foo, E.; Blake, S.N.; Fisher, B.J.; Smith, J.A.; Reid, J.B. The Role of Strigolactones during Plant Interactions with the Pathogenic Fungus Fusarium oxysporum. Planta 2016, 243, 1387–1396. [Google Scholar] [CrossRef]
- Fiorilli, V.; Vallino, M.; Biselli, C.; Faccio, A.; Bagnaresi, P.; Bonfante, P. Host and Non-Host Roots in Rice: Cellular and Molecular Approaches Reveal Differential Responses to Arbuscular Mycorrhizal Fungi. Front. Plant Sci. 2015, 6, 636. [Google Scholar] [CrossRef]
- Guo, R.; Wu, Y.; Liu, C.; Liu, Y.; Tian, L.; Cheng, J.; Pan, Z.; Wang, D.; Wang, B. OsADK1, a Novel Kinase Regulating Arbuscular Mycorrhizal Symbiosis in Rice. New Phytol. 2022, 234, 256–268. [Google Scholar] [CrossRef]
- Guo, N.; Li, L.; Cui, J.; Cai, B. Effects of Funneliformis mosseae on the Fungal Community in and Soil Properties of a Continuously Cropped Soybean System. Appl. Soil Ecol. 2021, 164, 103930. [Google Scholar] [CrossRef]
- Pozo, M.J.; Azcón-Aguilar, C. Unraveling Mycorrhiza-Induced Resistance. Curr. Opin. Plant Biol. 2007, 10, 393–398. [Google Scholar] [CrossRef]
- Boller, T.; He, S.Y. Innate Immunity in Plants: An Arms Race Between Pattern Recognition Receptors in Plants and Effectors in Microbial Pathogens. Science 2009, 324, 742–744. [Google Scholar] [CrossRef]
- Thomma, B.P.H.J.; Nürnberger, T.; Joosten, M.H.A.J. Of PAMPs and Effectors: The Blurred PTI-ETI Dichotomy. Plant Cell 2011, 23, 4–15. [Google Scholar] [CrossRef]
- Nishad, R.; Ahmed, T.; Rahman, V.J.; Kareem, A. Modulation of Plant Defense System in Response to Microbial Interactions. Front. Microbiol. 2020, 11, 1298. [Google Scholar] [CrossRef]
- Oldroyd, G.E.D. Speak, Friend, and Enter: Signalling Systems That Promote Beneficial Symbiotic Associations in Plants. Nat. Rev. Microbiol. 2013, 11, 252–263. [Google Scholar] [CrossRef]
- Lapin, D.; Van Den Ackerveken, G. Susceptibility to Plant Disease: More than a Failure of Host Immunity. Trends Plant Sci. 2013, 18, 546–554. [Google Scholar] [CrossRef]
- Cameron, D.D.; Neal, A.L.; Van Wees, S.C.M.; Ton, J. Mycorrhiza-Induced Resistance: More than the Sum of Its Parts? Trends Plant Sci. 2013, 18, 539–545. [Google Scholar] [CrossRef]
- Pieterse, C.M.J.; Van Der Does, D.; Zamioudis, C.; Leon-Reyes, A.; Van Wees, S.C.M. Hormonal Modulation of Plant Immunity. Annu. Rev. Cell Dev. Biol. 2012, 28, 489–521. [Google Scholar] [CrossRef]
- Song, F.; Song, G.; Dong, A.; Kong, X. Regulatory Mechanisms of Host Plant Defense Responses to Arbuscular Mycorrhiza. Acta Ecol. Sin. 2011, 31, 322–327. [Google Scholar] [CrossRef]
- Siciliano, V.; Genre, A.; Balestrini, R.; deWit, P.J.G.M.; Bonfante, P. Pre-Penetration Apparatus Formation During AM Infection Is Associated with a Specific Transcriptome Response in Epidermal Cells. Plant Signal. Behav. 2007, 2, 533–535. [Google Scholar] [CrossRef]
- Bai, L.; Sun, H.-B.; Liang, R.-T.; Cai, B.-Y. iTRAQ Proteomic Analysis of Continuously Cropped Soybean Root Inoculated With Funneliformis mosseae. Front. Microbiol. 2019, 10, 61. [Google Scholar] [CrossRef]


| Stress | Crops | AMF | AMF Colonization Effects | Ref |
|---|---|---|---|---|
| Drought | ||||
| Maize (Zea mays) | Rhizoglomus intraradices | - Improved growth traits, P, fresh leaf moisture content, WUE, and reduced C/P and N/P. | [30] | |
| Maize (Zea mays) | R. intraradices | - Amelioration of maize growth, water status, and Pi concentration. - Down-regulation of aldehyde oxidase expression and induced ABA signal transduction gene expression (D-myo-inositol-3-phosphate synthase and 14-3-3-like protein GF14). | [74] | |
| Wheat (Triticum Aestivum) | R. intraradices | - Improved leaf area, RWC, and WUE. - Enhanced N and P content and grain yielding. | [75] | |
| Maize (Zea mays) | Rhizophagus irregularis | - Ameliorated PSII efficiency and membrane stability and decreased lipids’ oxidative damage. -TSS overaccumulation and aquaporin gene expression (down-regulated ZmPIP1;1, ZmPIP1;3, ZmPIP1;4, ZmPIP1;6, ZmPIP2;2, ZmPIP2;4, ZmTIP1;1, and ZmTIP2;3 and up-regulated ZmTIP4;1). | [34] | |
| Sorghum (Sorghum bicolor) | F. mosseae | - Improved biomass and SLA and extended plant lifetime duration. | [47] | |
| Finger Millet (Eleusinecoracana) | R. intraradices | - Decreased EL, MDA, and hydrogen peroxide content. - Improved PRO, TSS, total phenol, and flavonoid content and antioxidant enzyme activities. | [50] | |
| Sorghum (Sorghum bicolor) | R. arabicus, R. irregularis | - Ameliorated transpiration efficiency and N and P acquisition. | [44] | |
| Wheat (Triticum Aestivum) | Glomus intraradices | - Improved RWC, flag leaf SLA, WUE, and mineral assimilation, particularly P. | [76] | |
| Maize (Zea mays) | G. versiforme | - Improved PH, SDM, and chlorophyll content. - Enhanced PRO, glycine betaine, TSS, free AAa, and phenols. - Decreased stress marker levels and improved GSH, CAT, POX, and SOD activities. | [51] | |
| Wheat (Triticum aestivum) | F. mosseae | - Enhanced SFW and SDW, N in roots, C/N ratio, and WUE. - Regulated plant secondary, oxidative stress metabolisms, and phytohormones’ crosstalk. | [35] | |
| Wheat (Triticum aestivum) | R. irregularis, F. mosseae | - Improved growth traits and N assimilation. - Regulated miR167, miR5384-3p, and miR156e-3p, influencing trafficking functionalities and cellular redox homeostasis. | [77] | |
| Maize (Zea mays) | R. irregularis | - Enhanced SFW and Pi acquisition - Moderated gs, transpiration, and WUE. | [45] | |
| Wheat (Triticum aestivum) | R. intraradices, F. mosseae, F. geosporum | - Improved plant and soil RWC. - Improved PSI and PSII quantum efficiency and photochemistry. | [43] | |
| Barley (Hordeum vulagre) | R. intraradices, F. mossea, C. claroideum, | - Improved plant growth traits. | [78] | |
| Rice (Oryza sativa) | F. mosseae, F. geosporus, R. irregularis, G.microaggregatum, C. claroideum | - Maintained rice growth and improved P acquisition. - Ameliorated nutrient, ABA, and IAA balance. - Higher grain yield, Chl fluorescence, and gs. | [42] | |
| Wheat (Triticum aestivum) | F. mosseae | - Enhanced plant shoot, root, and spike FW. - De-regulated transcriptional profiling, cell wall, and its membrane components. - Induced carbohydrate and lipid metabolism, cellulose synthase activity, membrane transport systems, N compound metabolic, and chitinase activity genes’ expression. | [37] | |
| Sorghum (Sorghum bicolor) | G. mosseae | - Ameliorated Chl a, b, and total Chl content, WUE, RWC, N, soluble proteins, and proline. - Enhanced yield, panicle length, number of panicles per plant, number of grains per panicle, and 1000-grain weight, and reduced EL and water saturation. | [36] | |
| Maize (Zea mays) | F. mosseae | - Enhanced Chl content, the net rate of photosynthesis, gs, rate of transpiration, and WUE. | [46] | |
| Wheat (Triticum aestivum) | F. mosseae | - Up-regulated water stress response-related genes (TdsHN1 and TdDRF1). | [79] | |
| Salinity | ||||
| Maize (Zea mays) | G. etunicatum | - Improved dry biomass and nutrient content and decreased Na+ assimilation. | [60] | |
| Wheat (Triticum aestivum) | G. intraradices | - Reduced shoot Na+ and enhanced N, P, K+, proline, Chl, protein, SA content, and total grain yielding. | [58] | |
| Rice (Oryza sativa) | C. etunicatum | - Improved SDW and root length. - Ameliorated gs and PSII efficiency, P and K+ content, K+/Na+ in shoots and reduced it in the roots. - Up-regulated OsNHX3, OsSOS1, OsHKT2;1, and OsHKT1;5. | [63] | |
| Maize (Zea mays) | R. intraradices | - Improved SDW and mineral uptake (P and N) and reduced leaf proline levels. | [52] | |
| Maize (Zea mays) | C. lamellosum Gigaspora margarita | - Ameliorated SDW, RDW, and nutrient content. - Reduced proline in the shoots and Na+ in the roots and higher K+/Na+ in roots. - Higher ZmAKT2, ZmSOS1, and ZmSKOR gene expression, sustaining K+ and Na+ homeostasis. | [61] | |
| Sorghum (Sorghum bicolor) | Acaulospora mellea | - Increased biomass, minerals, K+/Na+, leaf TSS content, and SOD, POX, CAT activities. | [67] | |
| Wheat (Triticum aestivum) | R. irregularis, F. mosseae, F. geosporum, C. claroideum | - Upgraded net photosynthesis rate, gs, reduced intrinsic WUE. - Higher carbon use efficiency and grain yielding. | [53] | |
| Maize (Zea mays) | R. irregulare | - Decreased Na+ level in root and its roots-to-shoots translocation. - K+ accumulation and Mg2+ reduction in roots. - Ca2+ fluctuation interacting with salinity. | [62] | |
| Rice (Oryza sativa) | R. irregularis | - Reduced H2O2 and enhanced CAT activity and leaf N content. - Enhanced tiller, panicle, grain number, and yield. | [54] | |
| Rice (Oryza sativa) | F. mosseae, A. laevis, G. margarita | - Improved total Chl, shoot K+/Na+ ratio, and grain yield. - Decreased shoot Na+/root Na+. | [64] | |
| Rice (Oryza sativa) | G. etunicatum, G. geosporum, G. mosseae | - Lower Na+/K+ ratio. - Maintained sucrose in flag leaf tissues and fructose and free proline overaccumulation. - Regulated cyanidin-3-glucoside and peonidin-3-glucosi.de in salt-sensitive rice. | [65] | |
| Maize (Zea mays) | F. mosseae | - Improved WUE, Chl, An, CAT, SOD, APX, and GSH activities - Decreased EL, MDA, and H2O2. | [80] | |
| Wheat (Triticum aestivum) | F. geosporum, F. mosseae, R. clarus, Scutellospora persica | - Promoted N, P, and K+ acquisition, Chl content, and K+/Na+ ratio. - Reduced Na+, Cl−, PRO, and MDA content. | [59] | |
| Maize (Zea mays) | G. mosseae | - Improved PH, leaf number, and plant FW and DW. - Enhanced nutrient content (N, P, and K+), and increased antioxidant enzyme activities. - Improved palmitoleic, oleic, linoleic, and linolenic acid contents. | [56] | |
| Sorghum (Sorghum bicolor) | F. mosseae, F. geosporum | - Improved PH, FW, DW, P, K+/Na+, and glomalin in soil. - Enhanced dehydrogenase and alkaline phosphatase activity. | [66] | |
| Maize (Zea mays) | R. irregularis | - Improved SDW, RDW, and RWC. | [57] | |
| Heavy Metals | ||||
| Wheat (Triticum aestivum) | Glomus sp. | - Reduced Zn content in shoots. | [81] | |
| Rice (Oryza sativa) | R. intraradices, F. mosseae | - Reduced Cd concentration in shoots and roots. | [31] | |
| Maize (Zea mays) | F. mosseae, Diversispora sphurcum | - Enhanced biomass, Chl, SOD, and CAT activities, and T-AOC and reduced H2O2, and MDA levels. - Limited Pb, Zn, and Cd transfer and contents in shoots. | [71] | |
| Rice (Oryza sativa) | F. mosseae, R. intraradices | - Reduced Cd concentrations in shoots and roots. - Iduced Nramp5 and HMA3 expression in roots. | [70] | |
| Sorghum (Sorghum bicolor) | C. etunicatum | - Enhanced biomass, PSII efficiency, and P, N, and S assimilation, and Mo accumulation. | [72] | |
| Maize (Zea mays) | C. etunicatum | - Increased SDW and SFW, and K+, P, Ca2+, Mg2+ in shoots. - Reduced lanthanum concentration in shoot and root. | [69] | |
| Wheat (Triticum aestivum) | R. intraradices | - Ameliorated growth and reduced Ni assimilation. | [73] | |
| AMF | Biotic Stress Agent | Plant | AMF Colonization Effects | Ref. |
|---|---|---|---|---|
| F. mosseae R. irregularis | Blumeria graminis f. sp. tritici | Wheat (Triticum aestivum) | - Reduced pathogenic fungus’ conidia number. - Overaccumulation of polyphenolic compounds. | [82] |
| C. claroideum, F. mosseae, R. irregulare | Oscinella frit | Wheat (Triticum aestivum) | - Ameliorated plant health status and production under pest’s heavy infestation. | [86] |
| F. mosseae | B. graminis f. sp. tritici | Wheat (Triticum aestivum) | - Decreased foliar biotrophic pathogen infection. - ISR and phenolic components’ overproduction. | [83] |
| F. mosseae | Xanthomonas translucens | Wheat (Triticum aestivum) | - Higher biomass, yield, and protein oxidation levels and decreased lesion area. - Up-regulated CYP enzymes, NTRs genes, and disease resilience genes. | [84] |
| R. intraradices | Magnaporthe oryzae | Rice (Oryza sativa) | - Induced IAA-/SA-related genes, key in pathogenesis-related protein synthesis. - Enriched JA, α-linolenic acid-, phenol-, and terpenoid syntheses-related genes. | [91] |
| F. mosseae, R. intraradices | M. oryzae | Rice (Oryza sativa) | - Improved Pi content and grain yield. - Higher resilience to the rice blast fungus. | [89] |
| R. irregulare | Sitobion avenae | Wheat (Triticum aestivum) | - Better harvest index and lower aphid population size. | [87] |
| R. intraradices | Spodoptera frugiperda | Rice (Oryza sativa) | - ISR activation and higher PPO and POX activity. | [92] |
| Enthropospora sp., Gigaspora sp., Glomus sp. | Peronosclerospora spp. | Maize (Zea mays) | - Enhanced SDM. - Attenuated downy mildew. - Expanded incubation period. | [90] |
| R. intraradices | Fusarium pseudograminearum | Wheat (Triticum aestivum) | - Improved biomass, spikes number, and height. - Enhanced antioxidant enzyme activity and reduced lipid peroxidation levels. - Decreased F. pseudograminearum density (76%) and disease severity (40%). | [88] |
| F. mosseae | X. translucens | Wheat (Triticum aestivum) | - Higher N acquisition, photosynthesis, and glucose and amino acid content. - Elicited defense-related proteins, immune response, and JA biosynthesis. | [85] |
| R. irregularis | Rhopalosiphum padi | Wheat (Triticum aestivum) | - Boosted root growth and P and N assimilation. | [93] |
| AMF | Biotic Stress Agent | Plant | AMF Colonization Effects | Ref. |
|---|---|---|---|---|
| R. intraradices, G. mosseae, G. etunicatum, G. claroideum, G. microaggregatum, G. geosporum | Sclerotinia sclerotiorum | Sunflower (Helianthus annuus) | - Hinder pathogenic hyphae development. - Localized and systemic resistance to white rot. | [120] |
| R. intraradices | Macrophomina phaseolina | Soybean (Glycine max) | - Reduced charcoal rot in soybean. | [119] |
| R. irregularis | M. phaseolina | Soybean (Glycine max) | - Up-regulated secondary metabolism genes. - Repressed genes encoding fasciclin-like arabinogalactan-protein, SKU5 similar 5, endo-chitinase, MYB, and POX. | [121] |
| R. irregularis | F. virguliforme | Soybean (Glycine max) | - Up-regulated defense-related genes like disease resistance proteins, WRKY, auxins, receptor kinases, proteases, thaumatin-like protein, pleiotropic drug resistance proteins/genes. - Down-regulated cell wall and POX genes. | [115] |
| R. intraradices | F. virguliforme | Soybean (Glycine max) | - Improved growth and P, K, Na, and S. - Decreased death syndrome severity. | [116] |
| R. intraradices | M. phaseolina | Soybean (Glycine max) | - Ameliorated biomass and greenness index. | [114] |
| G. etunicatum | Heterodera glycines | Soybean (Glycine max) | - Improved plant height and root system. - Decreased female nematodes in roots. | [117] |
| R. irregularis | Aphis glycines | Soybean (Glycine max) | - Improved biomass, nodulation, and N and C content. | [118] |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Slimani, A.; Ait-El-Mokhtar, M.; Ben-Laouane, R.; Boutasknit, A.; Anli, M.; Abouraicha, E.F.; Oufdou, K.; Meddich, A.; Baslam, M. Signals and Machinery for Mycorrhizae and Cereal and Oilseed Interactions towards Improved Tolerance to Environmental Stresses. Plants 2024, 13, 826. https://0-doi-org.brum.beds.ac.uk/10.3390/plants13060826
Slimani A, Ait-El-Mokhtar M, Ben-Laouane R, Boutasknit A, Anli M, Abouraicha EF, Oufdou K, Meddich A, Baslam M. Signals and Machinery for Mycorrhizae and Cereal and Oilseed Interactions towards Improved Tolerance to Environmental Stresses. Plants. 2024; 13(6):826. https://0-doi-org.brum.beds.ac.uk/10.3390/plants13060826
Chicago/Turabian StyleSlimani, Aiman, Mohamed Ait-El-Mokhtar, Raja Ben-Laouane, Abderrahim Boutasknit, Mohamed Anli, El Faiza Abouraicha, Khalid Oufdou, Abdelilah Meddich, and Marouane Baslam. 2024. "Signals and Machinery for Mycorrhizae and Cereal and Oilseed Interactions towards Improved Tolerance to Environmental Stresses" Plants 13, no. 6: 826. https://0-doi-org.brum.beds.ac.uk/10.3390/plants13060826