
Shifting Mountain Tree Line Increases Soil Organic Carbon Stability Regardless of Land Use
 , , , and
, , , and
Abstract
:1. Introduction
2. Results
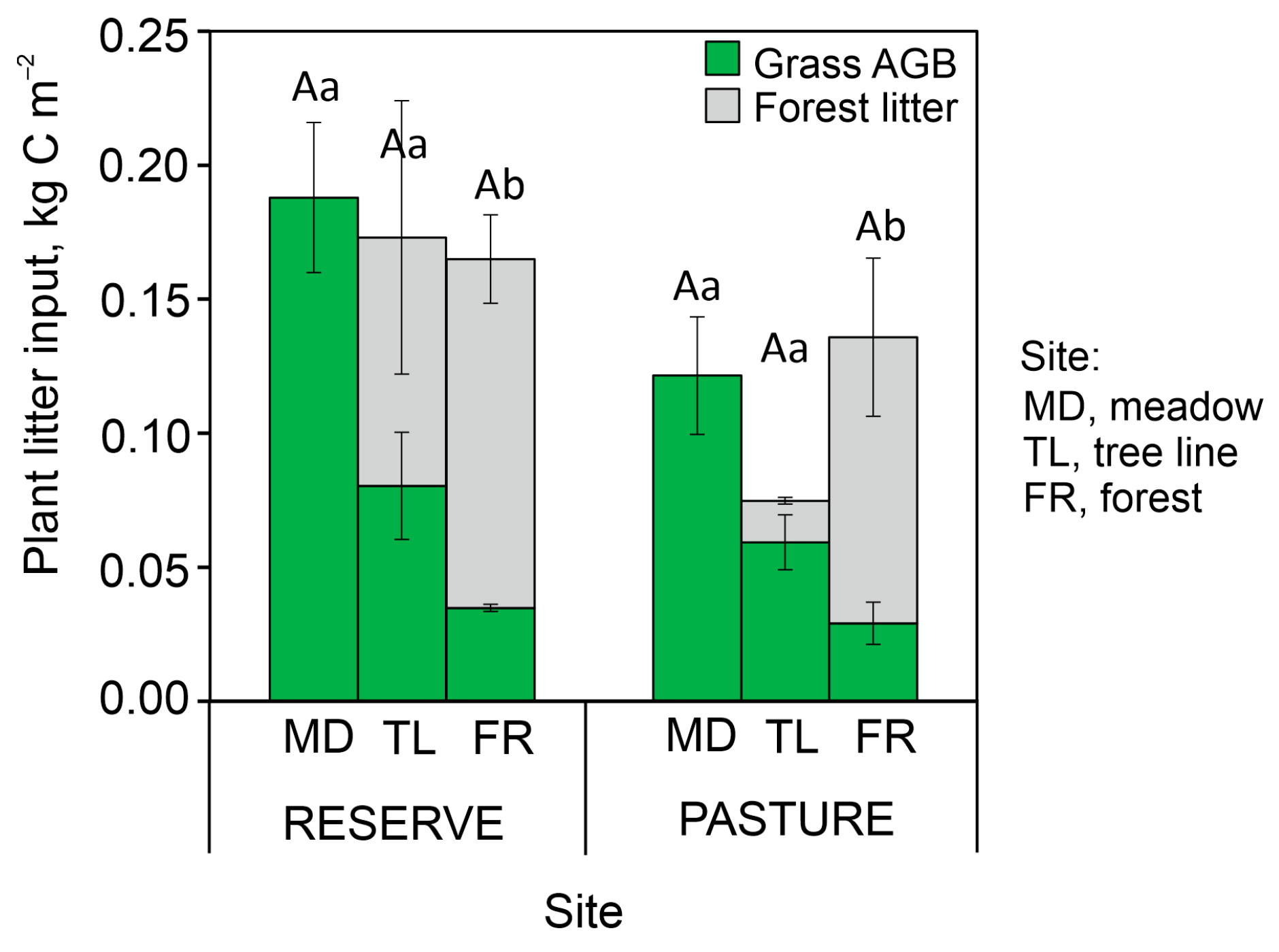
2.1. Plant Aboveground Litter and Soil Texture
2.2. Topsoil C Stock Distribution between POM–C and MAOM–C Pools
3. Discussion
4. Materials and Methods
4.1. Study Area and Sampling Design
4.2. Plant and Soil Samples Analysis
4.3. Data Analysis
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
Appendix A

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Site | Alkyl C (0–45 ppm) | O-Alkyl C (45–110 ppm) | Aromatic C (110–165 ppm) | Aromaticity | C/N |
|---|---|---|---|---|---|
| Reserve | |||||
| Grass AGB: | |||||
| Meadow | 5.6 ± 0.3 | 62.3 ± 0.5 | 18.6 ± 0.1 | 0.22 ± 0.00 | 33 ± 2 |
| Tree line | 6.2 ± 0.1 | 61.2 ± 0.5 | 18.9 ± 0.2 | 0.22 ± 0.00 | 24 ± 1 |
| Forest | 10.4 ± 0.8 | 58.8 ± 1.0 | 16.7 ± 0.5 | 0.19 ± 0.01 | 22 ± 1 |
| Forest litter | 14.9 ± 1.0 | 49.8 ± 1.1 | 23.0 ± 1.1 | 0.26 ± 0.01 | 24 ± 1 |
| Pasture | |||||
| Grass AGB: | |||||
| Meadow | 6.9 ± 0.5 | 62.1 ± 1.0 | 18.3 ± 0.5 | 0.21 ± 0.01 | 29 ± 1 |
| Tree line | 7.8 ± 0.7 | 61.2 ± 0.9 | 18.1 ± 0.1 | 0.21 ± 0.00 | 28 ± 2 |
| Forest | 8.0 ± 0.3 | 63.1 ± 2.3 | 17.4 ± 0.5 | 0.20 ± 0.01 | 25 ± 3 |
| Forest litter | 14.1 ± 0.7 | 48.0 ± 1.7 | 23.8 ± 0.7 | 0.28 ± 0.01 | 27 ± 1 |
| Site | POM–C | MAOM–C | ||||
|---|---|---|---|---|---|---|
| g kg−1 | kg C m−2 | % of Total C | g kg−1 | kg C m−2 | % of Total C | |
| Reserve | ||||||
| Meadow | 148 ± 16 | 3.7 ± 0.2 | 92 ± 2 | 12 ± 3 | 0.3 ± 0.1 | 7 ± 2 |
| Tree line | 116 ± 12 | 4.8 ± 0.4 | 89 ± 1 | 14 ± 0 | 0.6 ± 0.0 | 11 ± 1 |
| Forest | 93 ± 26 | 3.9 ± 1.4 | 82 ± 5 | 18 ± 0 | 0.7 ± 0.1 | 18 ± 5 |
| Pasture | ||||||
| Meadow | 81 ± 9 | 4.3 ± 0.7 | 74 ± 6 | 28 ± 7 | 1.5 ± 0.3 | 26 ± 6 |
| Tree line | 85 ± 12 | 4.6 ± 1.0 | 73 ± 6 | 30 ± 6 | 1.5 ± 0.1 | 27 ± 6 |
| Forest | 33 ± 2 | 1.9 ± 0.2 | 49 ± 2 | 34 ± 1 | 1.9 ± 0.0 | 51 ± 2 |
| Parameter | Reserve | Pasture |
|---|---|---|
| Plant litter quantity and quality | ||
| Grass AGB, kg C m−2 | 0.86 ** | 0.54 |
| Total plant litter 1, kg C m−2 | 0.45 | −0.18 |
| Aromatic C, % | −0.35 | −0.65 |
| Aromaticity | −0.36 | −0.68 * |
| C/N ratio | 0.69 * | 0.55 |
| Soil texture | ||
| Silt, % | −0.58 | -0.18 |
| Clay, % | −0.79 ** | -0.60 |
| Site | PC (%) | NS | Dominant Species (≥10% of PC) |
|---|---|---|---|
| Reserve | |||
| Meadow | 98 ± 2 | 12 ± 1 | Calamagrostis arundinacea, Stachys macrantha, Ranunculus caucasicus subsp. subleiocarpus, Astrantia maxima |
| Tree line | 62 ± 6 | 7 ± 1 | Calamagrostis arundinacea, Pulsatilla aurea, Veratrum album, Gentiana septemfida |
| Forest | 40 ± 6 | 8 ± 1 | Aconitum orientale, Prenantes abietina, Myosotis amoena, Petasites albus |
| Pasture | |||
| Meadow | 93 ± 2 | 10 ± 0 | Nardus stricta, Cirsium obvalatum, Potentilla erecta, Agrostis capillaris, Stachys macrantha |
| Tree line | 75 ± 9 | 9 ± 2 | Agrostis capillaris, Prunella vulgaris, Myosotis amoena |
| Forest | 53 ± 7 | 10 ± 0 | Alchemilla vulgaris, Oxalis acetosella, Trifolium repens |

References
- Hansen, A.J.; Neilson, R.P.; Dale, V.H.; Flather, C.H.; Iverson, L.R.; Currie, D.J.; Shafer, S.; Cook, R.; Bartlein, P.J. Global change in forests: Responses of species, communities, and biomes. BioScience 2001, 51, 765–779. [Google Scholar] [CrossRef]
- Jump, A.S.; Huang, T.J.; Chou, C.H. Rapid altitudinal migration of mountain plants in Taiwan and its implications for high altitude biodiversity. Ecography 2012, 35, 204–210. [Google Scholar] [CrossRef]
- Cudlin, P.; Klopcic, M.; Tognetti, R.; Malis, F.; Alados, C.L.; Bebi, P.; Grunewald, K.; Zhiyanski, M.; Andonowski, V.; La Porta, N.; et al. Drivers of treeline shift in different European mountains. Clim. Res. 2017, 73, 135–150. [Google Scholar] [CrossRef]
- Guidi, C.; Vesterdal, L.; Gianelle, D.; Rodeghiero, M. Changes in soil organic carbon and nitrogen following forest expansion on grassland in the Southern Alps. For. Ecol. Manag. 2014, 28, 103–116. [Google Scholar] [CrossRef]
- Kammer, A.; Hagedorn, F.; Shevchenko, I.; Leifeld, J.; Guggenberger, G.; Goryacheva, T.; Riging, A.; Moiseev, P. Treeline shifts in the Ural mountains affect soil organic matter dynamics. Glob. Chang. Biol. 2009, 15, 1570–1583. [Google Scholar] [CrossRef]
- Zimmermann, M.; Meir, P.; Silman, M.R.; Fedders, A.; Gibbon, A.; Malhi, Y.; Urrego, D.H.; Bush, M.B.; Feeley, K.J.; Garcia, K.C.; et al. No differences in soil carbon stocks across the tree line in the Peruvian Andes. Ecosystems 2010, 13, 62–74. [Google Scholar] [CrossRef]
- Cotrufo, M.F.; Ranalli, M.G.; Haddix, M.L.; Six, J.; Lugato, E. Soil carbon storage informed by particulate and mineral-associated organic matter. Nat. Geosci. 2019, 12, 989–994. [Google Scholar] [CrossRef]
- Lavallee, J.M.; Soong, J.L.; Cotrufo, M.F. Conceptualizing soil organic matter into particulate and mineral-associated forms to address global change in the 21st century. Glob. Chang. Biol. 2020, 26, 261–273. [Google Scholar] [CrossRef] [PubMed]
- Guidi, C.; Magid, J.; Rodeghiero, M.; Gianelle, D.; Vesterdal, L. Effects of forest expansion on mountain grassland: Changes within soil organic carbon fractions. Plant Soil 2014, 385, 373–387. [Google Scholar] [CrossRef]
- Ortiz, C.; Fernandez-Alonso, M.J.; Kitzler, B.; Diaz-Pines, E.; Saiz, G.; Rubio, A.; Benito, M. Variations in soil aggregation, microbial community structure and soil organic matter cycling associated to long-term afforestation and woody encroachment in a Mediterranean alpine ecotone. Geoderma 2022, 405, 115450. [Google Scholar] [CrossRef]
- Briggs, J.M.; Knapp, A.K.; Blair, J.M.; Heisler, J.L.; Hoch, G.A.; Lett, M.S.; McCarron, J.K. An ecosystem in transition: Causes and consequences of the conversion of mesic grassland to shrubland. BioScience 2005, 55, 243–254. [Google Scholar] [CrossRef]
- Pilon, N.A.L.; Durigan, G.; Rickenback, J.; Pennington, R.T.; Dexter, K.G.; Hoffmann, W.A.; Abreu, R.C.R.; Lehmann, C.E.R. Shade alters savanna grass layer structure and function along a gradient of canopy cover. J. Veg. Sci. 2021, 32, e12959. [Google Scholar] [CrossRef]
- Almeida, L.F.J.; Souza, I.F.; Hurtarte, L.C.C.; Teixeira, P.P.C.; Inagaki, T.M.; Silva, I.R.; Mueller, C.W. Forest litter constraints on the pathways controlling soil organic matter formation. Soil Biol. Biochem. 2021, 163, 108447. [Google Scholar] [CrossRef]
- Gunina, A.; Kuzyakov, Y. Pathways of litter C by formation of aggregates and SOM density fractions: Implications from 13C natural abundance. Soil Biol. Biochem. 2014, 71, 95–104. [Google Scholar] [CrossRef]
- Parwada, C.; Van Tol, J. Effects of litter source on the dynamics of particulate organic matter fractions and rates of macroaggregate turnover in different soil horizons. Eur. J. Soil Sci. 2018, 69, 1126–1136. [Google Scholar] [CrossRef]
- Mainka, M.; Summerauer, L.; Wasner, D.; Garland, G.; Griepentrog, M.; Berhe, A.A.; Doetterl, S. Soil geochemistry as a driver of soil organic matter composition: Insights from a soil chronosequence. Biogeosciences 2022, 19, 1675–1689. [Google Scholar] [CrossRef]
- Sowinski, P. Effect of slope position on soil particle-size distribution in young glacial landscape (Łyna River valley, NE Poland). Soil Sci. Annu. 2016, 67, 140–150. [Google Scholar] [CrossRef]
- Austin, A.T.; Ballare, C.L. Dual role of lignin in plant litter decomposition in terrestrial ecosystems. Proc. Natl. Acad. Sci. USA 2010, 107, 4618–4622. [Google Scholar] [CrossRef] [PubMed]
- Witkamp, M.; van der Drift, J. Breakdown of forest litter in relation to environmental factors. Plant Soil 1961, 15, 295–311. [Google Scholar] [CrossRef]
- Thurow, T.L. Decomposition of grasses and forbs in coastal savanna of southern Somalia. Afr. J. Ecol. 1989, 27, 201–206. [Google Scholar] [CrossRef]
- Gavazov, K.S. Dynamics of alpine plant litter decomposition in a changing climate. Plant Soil 2010, 337, 19–32. [Google Scholar] [CrossRef]
- Su, Y.; Gong, Y.; Han, W.; Li, K.; Liu, X. Dependency of litter decomposition on litter quality, climate change, and grassland type in the alpine grassland of Tianshan Mountains, Northwest China. J. Arid Land 2022, 14, 691–703. [Google Scholar] [CrossRef]
- Leifeld, J.; Zimmermann, M.; Fuhrer, J. Storage and turnover of carbon in grassland soils along an elevation gradient in the Swiss Alps. Glob. Chang. Biol. 2009, 15, 668–679. [Google Scholar] [CrossRef]
- Budge, K.; Leifeld, J.; Hiltbrunner, E.; Fuhrer, J. Litter quality and pH are strong drivers of carbon turnover and distribution in alpine grassland soils. Biogeosci. Discuss. 2010, 7, 6207–6242. [Google Scholar] [CrossRef]
- Liu, Y.-F.; Liu, Y.; Shi, Z.-H.; López-Vicente, M.; Wu, G.-L. Effectiveness of re-vegetated forest and grassland on soil erosion control in the semi-arid Loess Plateau. Catena 2020, 195, 104787. [Google Scholar] [CrossRef]
- Li, G.; Wan, L.; Cui, M.; Wu, B.; Zhou, J. Influence of canopy interception and rainfall kinetic energy on soil erosion under forests. Forests 2019, 10, 509. [Google Scholar] [CrossRef]
- Wilson, M.J. The origin and formation of clay minerals in soils: Past, present and future perspectives. Clay Miner. 1999, 34, 7–25. [Google Scholar] [CrossRef]
- Lützow, M.V.; Kögel-Knabner, I.; Ekschmitt, K.; Matzner, E.; Guggenberger, G.; Marschner, B.; Flessa, H. Stabilization of organic matter in temperate soils: Mechanisms and their relevance under different soil conditions—A review. Eur. J. Soil Sci. 2006, 57, 426–445. [Google Scholar] [CrossRef]
- Selezneva, A.E.; Ivashchenko, K.V.; Sushko, S.V.; Zhuravleva, A.I.; Ananyeva, N.D.; Blagodatsky, S.A. Microbial respiration and functional diversity of soil microbial community under treeline shifts in the Northwestern Caucasus. RUDN J. Agron. Anim. Ind. 2021, 16, 226–237. (In Russian) [Google Scholar] [CrossRef]
- Angst, G.; Mueller, K.E.; Nierop, K.G.J.; Simpson, M.J. Plant- or microbial-derived? A review on the molecular composition of stabilized soil organic matter. Soil Biol. Biochem. 2021, 156, 108189. [Google Scholar] [CrossRef]
- Caucasian State Nature Biosphere Reserve. Available online: https://www.kavkazzapoved.ru/ (accessed on 20 March 2024).
- IUSS Working Group WRB. World Reference Base for Soil Resources 2014: International Soil Classification System for Naming Soils and Creating Legends for Soil Maps; Schad, P., Huysteen, C., Micheli, E., Eds.; Food and Agriculture Organization: Rome, Italy, 2015. [Google Scholar]
- Regional Geological Survey Data of Russian Geological Research Institute (VSEGEI). Available online: https://karpinskyinstitute.ru/en/ (accessed on 15 February 2024).
- Komarova, A.; Ivashchenko, K.; Sushko, S.; Zhuravleva, A.; Vasenev, V.; Blagodatsky, S. Temperature sensitivity of topsoil organic matter decomposition does not depend on vegetation types in mountains. Plants 2022, 11, 2765. [Google Scholar] [CrossRef] [PubMed]
- Kögel-Knabner, I. 13C and 15N NMR spectroscopy as a tool in soil organic matter studies. Geoderma 1997, 80, 243–270. [Google Scholar] [CrossRef]
- Carvalho, A.M.; Bustamante, M.M.C.; Alcantara, F.A.; Resck, I.S.; Lemos, S.S. Characterization by solid-state CPMAS 13C NMR spectroscopy of decomposing plant residues in conventional and no-tillage systems in Central Brazil. Soil Tillage Res. 2009, 102, 144–150. [Google Scholar] [CrossRef]
- Yudina, A.V.; Fomin, D.S.; Valdes-Korovkin, I.A.; Churilin, N.A.; Kovda, I.V.; Milanovskiy, E.Y.; Aleksandrova, M.S.; Golovleva, Y.A.; Philipov, N.V.; Dymov, A.A. The ways to develop soil texture classification for laser diffraction method. Eurasian Soil Sci. 2020, 53, 1579–1595. [Google Scholar] [CrossRef]
- Soil Survey Staff. Keys to Soil Taxonomy, 12th ed.; USDA-Natural Resources Conservation Service: Washington, DC, USA, 2014.
- Cambardella, C.A.; Elliott, E.T. Particulate soil organic-matter changes across a grassland cultivation sequence. Soil Sci. Soc. Am. J. 1992, 56, 777–783. [Google Scholar] [CrossRef]
- Guo, Z.; Wang, Y.; Wan, Z.; Zuo, Y.; He, L.; Li, D.; Yuan, F.; Wang, N.; Liu, J.; Song, Y.; et al. Soil dissolved organic carbon in terrestrial ecosystems: Global budget, spatial distribution and controls. Glob. Ecol. Biogeogr. 2020, 29, 2159–2175. [Google Scholar] [CrossRef]





| Site | Reserve | Pasture | ||||
|---|---|---|---|---|---|---|
| Aromatic C, % | Aromaticity | C/N | Aromatic C, % | Aromaticity | C/N | |
| Meadow | 18.6 ± 0.1 b | 0.22 ± 0.00 | 33 ± 2 | 18.3 ± 0.5 b | 0.21 ± 0.01 b | 29 ± 1 |
| Tee line | 20.2 ± 0.7 ab | 0.23 ± 0.01 | 23 ± 0 | 19.3 ± 0.2 ab | 0.22 ± 0.00 ab | 28 ± 2 |
| Forest | 21.7 ± 1.1 a | 0.25 ± 0.01 | 23 ± 0 | 22.3 ± 0.3 a | 0.26 ± 0.00 a | 26 ± 1 |
| p-value | 0.05 | 0.06 | 0.07 | 0.03 | 0.04 | 0.25 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Sushko, S.; Ivashchenko, K.; Komarova, A.; Yudina, A.; Makhantseva, V.; Elsukova, E.; Blagodatsky, S. Shifting Mountain Tree Line Increases Soil Organic Carbon Stability Regardless of Land Use. Plants 2024, 13, 1193. https://0-doi-org.brum.beds.ac.uk/10.3390/plants13091193
Sushko S, Ivashchenko K, Komarova A, Yudina A, Makhantseva V, Elsukova E, Blagodatsky S. Shifting Mountain Tree Line Increases Soil Organic Carbon Stability Regardless of Land Use. Plants. 2024; 13(9):1193. https://0-doi-org.brum.beds.ac.uk/10.3390/plants13091193
Chicago/Turabian StyleSushko, Sofia, Kristina Ivashchenko, Alexandra Komarova, Anna Yudina, Victoria Makhantseva, Ekaterina Elsukova, and Sergey Blagodatsky. 2024. "Shifting Mountain Tree Line Increases Soil Organic Carbon Stability Regardless of Land Use" Plants 13, no. 9: 1193. https://0-doi-org.brum.beds.ac.uk/10.3390/plants13091193