
A Substantial Fraction of Barley (Hordeum vulgare L.) Low Phytic Acid Mutations Have Little or No Effect on Yield across Diverse Production Environments

Abstract
:1. Introduction
2. Results
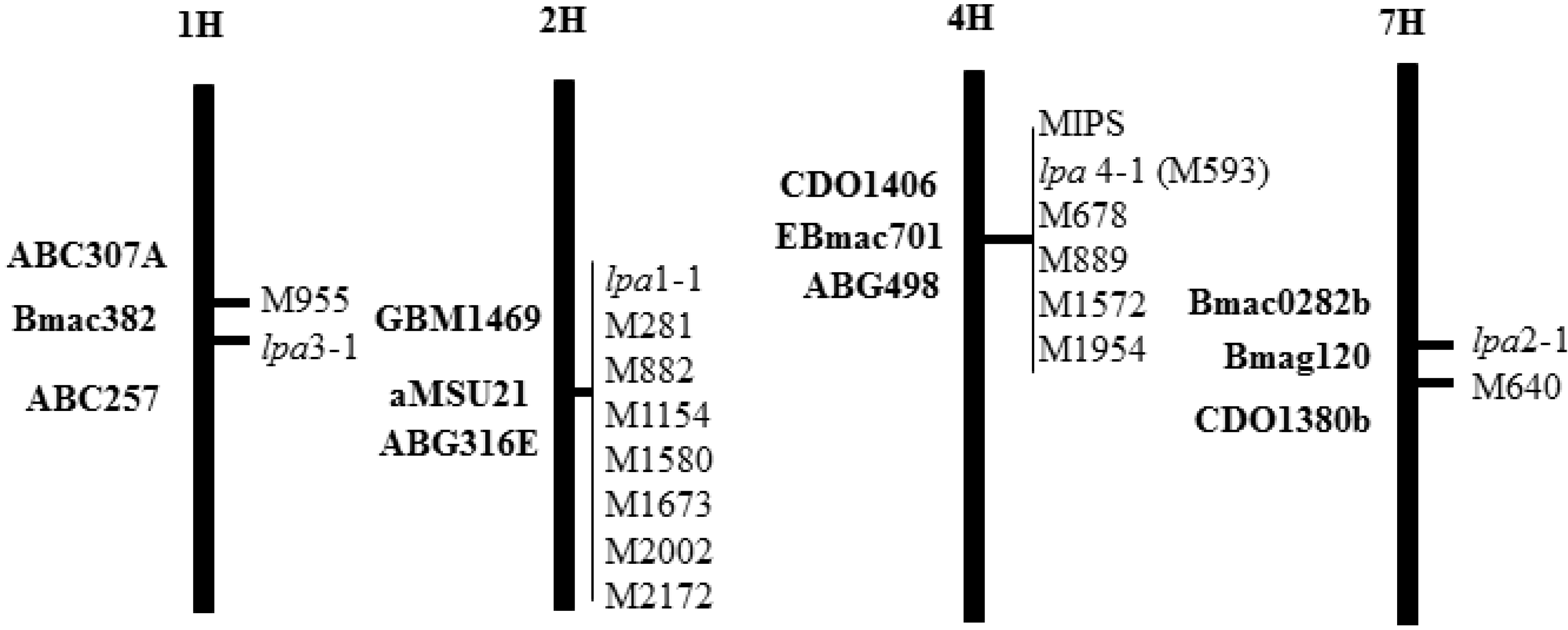
2.1. Barley Low Phytic Acid Mutations Selected for Isoline Development

2.2. Confirmation of Isoline Seed Phytic Acid Phenotype
2.3. Yield Evaluations

{kind=link}
| Isoline Pair | Genotype | Idaho Field Location | ||||||
|---|---|---|---|---|---|---|---|---|
| Aberdeen | Soda Springs | |||||||
| Total P | Phytic Acid P | Inorganic P | Total P | Phytic Acid P | Inorganic P | |||
| mg·g−1 | ||||||||
| 1 | WT | 2.96 | 2.86 | 0.29 | 3.10 | 2.62 | 0.26 | |
| 1 | Hvlpa1-1 | 2.52 | 1.51 | 1.05 | 2.44 | 1.35 | 0.83 | |
| 2 | WT | 2.86 | 2.47 | 0.24 | 3.09 | 2.75 | 0.25 | |
| 2 | Hvlpa3-1 | 2.76 | 1.10 | 1.45 | 2.88 | 1.15 | 1.66 | |
| 3 | WT | 2.78 | 2.32 | 0.22 | 3.12 | 2.52 | 0.24 | |
| 3 | Hvlpa-M955 | 2.91 | 0.53 | 2.12 | 3.33 | 0.51 | 2.21 | |
| 4 | WT | 2.75 | 2.38 | 0.23 | 2.93 | 2.54 | 0.25 | |
| 4 | Hvlpa2-1 | 2.93 | 1.83 | 0.67 | 3.18 | 1.99 | 0.90 | |
| 5 | WT | 3.10 | 2.49 | 0.39 | 2.98 | 2.59 | 0.27 | |
| 5 | Hvlpa4-1 | 2.91 | 1.53 | 1.18 | 3.31 | 1.76 | 1.34 | |
| 6 | WT | 2.86 | 2.44 | 0.26 | 3.15 | 2.69 | 0.26 | |
| 6 | Hvlpa-M640 | 2.98 | 1.61 | 0.80 | 3.36 | 1.51 | 1.32 | |
| Cultivar Controls | ||||||||
| Harrington | 2.73 | 2.37 | 0.26 | 3.07 | 2.59 | 0.30 | ||
| Baronesse | 2.89 | 2.41 | 0.26 | 3.12 | 2.65 | 0.28 | ||
| Conrad | 3.01 | 2.52 | 0.19 | 3.49 | 3.10 | 0.31 | ||
| Std. Dev. of Mean | 0.12 | 0.14 | 0.07 | 0.15 | 0.14 | 0.05 | ||
| LSD p = 0.05 | 0.34 | 0.41 | 0.20 | 0.43 | 0.39 | 0.15 | ||
| Source of Variation | df | Mean Square | F Value | Prob. > F |
|---|---|---|---|---|
| Model | 132 | 10.20 | 28.82 | <0.0001 |
| Year | 1 | 97.24 | 274.70 | <0.0001 |
| Location | 4 | 224.79 | 635.05 | <0.0001 |
| Year × Loc | 4 | 61.43 | 173.54 | <0.0001 |
| Rep (Loc) | 15 | 0.65 | 1.85 | 0.03 |
| Isoline | 11 | 3.70 | 10.45 | <0.0001 |
| Year × Line | 11 | 0.22 | 0.62 | 0.82 |
| Loc × Line | 44 | 0.73 | 2.06 | 0.0002 |
| Year × Loc × Line | 42 | 0.47 | 1.31 | 0.11 |
| Residual Error | 330 | 0.35 |
| Isoline Pair | Genotype | Idaho Location | Line Mean | ||||
|---|---|---|---|---|---|---|---|
| Aberdeen | Ashton | Idaho Falls | Rupert | Soda Springs | |||
| Grain Yield (Mg·ha−1) | |||||||
| 1 | WT | 6.04 | 2.52 | 6.15 | 6.20 | 2.13 | 4.61 |
| 1 | Hvlpa1-1 | 6.61 | 2.78 | 6.57 | 5.47 | 2.00 | 4.69 |
| 2 | WT | 5.83 | 2.37 | 5.84 | 6.20 | 2.22 | 4.49 |
| 2 | Hvlpa3-1 | 6.17 | 2.03 | 5.71 | 5.15 | 1.88 | 4.19 |
| 3 | WT | 5.72 | 2.59 | 5.98 | 5.45 | 2.80 | 4.60 |
| 3 | Hvlpa-M955 | 5.27 | 1.88 | 5.71 | 4.74 | 1.47 | 3.81 |
| 4 | WT | 6.48 | 2.34 | 6.15 | 5.89 | 2.41 | 4.91 |
| 4 | Hvlpa2-1 | 6.51 | 2.59 | 5.98 | 5.23 | 1.81 | 3.86 |
| 5 | WT | 6.14 | 2.32 | 6.45 | 5.71 | 2.52 | 4.63 |
| 5 | Hvlpa4-1 | 6.06 | 2.30 | 6.74 | 5.45 | 1.96 | 4.50 |
| 6 | WT | 6.59 | 2.30 | 6.77 | 5.89 | 2.10 | 4.73 |
| 6 | Hvlpa-M640 | 6.30 | 2.40 | 6.45 | 4.44 | 1.61 | 3.58 |
| Cultivar Controls | |||||||
| Harrington | 6.38 | 2.42 | 5.54 | 5.54 | 2.22 | 4.42 | |
| Baronesse | 7.17 | 3.57 | 6.81 | 7.45 | 2.59 | 5.52 | |
| Conrad | 7.06 | 2.37 | 6.08 | 6.91 | 2.05 | 4.89 | |
| Location Mean ± SE | 6.29 ± 0.34 | 2.43 ± 0.21 | 6.19 ± 0.29 | 5.78 ± 0.22 | 2.10 ± 0.19 | 4.52 ± 0.22 | |
| LSD p = 0.05 | 0.98 | 0.59 | 0.83 | 0.63 | 0.55 | 0.64 | |
| Isoline Pair | Genotype | Idaho Location | Line Mean | ||||
|---|---|---|---|---|---|---|---|
| Aberdeen | Ashton | Idaho Falls | Rupert | Soda Springs | |||
| Grain Yield (Mg·ha−1) | |||||||
| 1 | WT | 6.51 | 5.79 | 6.10 | 5.22 | 3.05 | 5.33 |
| 1 | Hvlpa1-1 | 7.35 | 6.69 | 5.57 | 5.57 | 3.25 | 5.68 |
| 2 | WT | 6.80 | 6.15 | 4.93 | 4.98 | 3.55 | 5.37 |
| 2 | Hvlpa3-1 | 6.98 | 5.32 | 4.96 | 5.01 | 2.78 | 5.01 |
| 3 | WT | 6.40 | 6.34 | 5.57 | 4.84 | 3.59 | 5.35 |
| 3 | Hvlpa-M955 | 5.07 | 4.83 | 4.86 | 4.61 | 2.47 | 4.37 |
| 4 | WT | 7.03 | 6.32 | 5.52 | 5.10 | 3.42 | 5.48 |
| 4 | Hvlpa2-1 | 7.51 | 4.57 | 6.27 | 5.03 | 2.78 | 5.23 |
| 5 | WT | 7.80 | 6.37 | 5.96 | 5.25 | 3.44 | 5.76 |
| 5 | Hvlpa4-1 | 6.95 | 5.84 | 5.27 | 5.88 | 2.99 | 5.51 |
| 6 | WT | 7.29 | 6.01 | 5.40 | 5.48 | 3.20 | 5.48 |
| 6 | Hvlpa-M640 | 6.98 | 5.03 | 5.93 | 4.95 | 2.47 | 5.21 |
| Cultivar Controls | |||||||
| Harrington | 6.96 | 6.27 | 5.69 | 5.30 | 3.17 | 5.44 | |
| Baronesse | 8.61 | 7.13 | 6.86 | 6.08 | 4.05 | 6.55 | |
| Conrad | 8.00 | 6.71 | 6.64 | 6.74 | 3.03 | 6.22 | |
| Location Mean ± SE | 7.08 ± 0.37 | 5.96 ± 0.32 | 5.69 ± 0.29 | 5.34 ± 0.36 | 3.16 ± 0.21 | 5.47 ± 0.28 | |
| LSD p = 0.05 | 1.05 | 0.93 | 0.83 | 1.02 | 0.59 | 0.79 | |
3. Discussion
4. Experimental Section
4.1. Isoline Pair Development and Cultivar Controls
4.2. Test Sites
4.3. Data Collection
4.4. Statistical Analyses
5. Conclusions
Supplementary Files
Supplementary File 1Acknowledgments
Author Contributions
Conflicts of Interest
References
- Cichy, K.; Raboy, V. Evaluation and development of low-phytate crops. In Modification of Seed Composition to Promote Health and Nutrition; Krishnan, H., Ed.; American Society of Agronomy: Madison, WI, USA, 2009; pp. 177–200. [Google Scholar]
- Raboy, V. Approaches and challenges to engineering seed phytate and total phosphorus. Plant Sci. 2009, 177, 281–296. [Google Scholar] [CrossRef]
- Shi, J.; Wang, H.; Schellin, K.; Li, B.; Faller, M.; Stoop, J.M.; Meeley, R.B.; Ertl, D.S.; Ranch, J.P.; Glassman, K. Embryo-specific silencing of a transporter reduces phytic acid content of maize and soybean seeds. Nat. Biotechnol. 2007, 25, 930–937. [Google Scholar] [CrossRef] [PubMed]
- Shukla, V.K.; Donyon, Y.; Miller, J.C.; DeKelver, R.C.; Moehle, E.A.; Worden, S.E.; Mitchell, J.C.; Arnold, N.L.; Gopolan, S.; Meng, X.; et al. Precise genome modification in the crop species Zea mays using zinc-finger nucleases. Nature 2009, 459, 437–443. [Google Scholar] [CrossRef] [PubMed]
- Bregitzer, P.; Raboy, V. Effects of four independent Low-Phytate mutations on barley agronomic performance. Crop Sci. 2006, 46, 1318–1322. [Google Scholar] [CrossRef]
- Bregitzer, P.; Raboy, V.; Obert, D.E.; Windes, J.M.; Whitmore, J.C. Registration of “Herald” Barley. Crop Sci. 2007, 47, 441–442. [Google Scholar] [CrossRef]
- Ye, H.; Zhang, X.-Q.; Broughton, S.; Westcott, S.; Wu, D.; Lance, R.; Li, C. A nonsense mutation in a putative sulphate transporter gene results in low phytic acid in barley. Funct. Integr. Genomics 2011, 11, 103–110. [Google Scholar] [CrossRef] [PubMed]
- Ockenden, I.; Dorsch, J.A.; Reid, M.M.; Lin, L.; Grant, L.K.; Raboy, V.; Lott, J.N.A. Characterization of the storage of phosphorus, inositol phosphate and cations in grain tissues of four barley (Hordeum vulgare L.) low phytic acid genotypes. Plant Sci. 2004, 167, 1131–1142. [Google Scholar] [CrossRef]
- Bregitzer, P.; Raboy, V.; Obert, D.E. Registration of “Clearwater” low-phytate hulless spring barley. J. Plant Regist. 2008, 2, 1–4. [Google Scholar] [CrossRef]
- Stelpflug, S.C.; Eichten, S.R.; Hermanson, P.J.; Springer, N.M.; Kaeppler, S.M. Consistent and heritable alterations of DNA methylation are induced by tissue culture in maize. Genetics 2014, 198, 209–218. [Google Scholar] [CrossRef] [PubMed]
- Brown, R.H.; Raboy, V.; Bregitzer, P. Unintended consequences: High phosphinothricin acetyltransferase activity related to reduced fitness in barley. In Vitro Cell. Dev. Biol. 2013, 49, 240–247. [Google Scholar] [CrossRef]
- Ertl, D.; Young, K.A.; Raboy, V. Plant genetic approaches to phosphorus management in agricultural production. J. Environ. Qual. 1998, 27, 299–304. [Google Scholar] [CrossRef]
- Raboy, V.; Gerbasi, P.F.; Young, K.A.; Stoneberg, S.D.; Pickett, S.G.; Bauman, A.T.; Murthy, P.P.N.; Sheridan, W.F.; Ertl, D.S. Origin and seed phenotype of maize low phytic acid 1-1 and low phytic acid 2-1. Plant Physiol. 2000, 124, 355–368. [Google Scholar] [CrossRef] [PubMed]
- Meis, S.J.; Fehr, W.R.; Schnebly, S.R. Seed source effect on field emergence of soybean lines with reduced phytate and raffinose saccharides. Crop Sci. 2003, 43, 1336–1339. [Google Scholar] [CrossRef]
- Oltmans, S.E.; Fehr, W.R.; Welke, G.A.; Raboy, V.; Peterson, K.L. Agronomic and seed traits of soybean lines with low-phytate phosphorus. Crop Sci. 2005, 45, 593–598. [Google Scholar] [CrossRef]
- Anderson, B.P.; Fehr, W.R. Seed source affects field emergence of low-phytate soybean lines. Crop Sci. 2008, 48, 929–932. [Google Scholar] [CrossRef]
- Hitz, W.; Carlson, T.J.; Kerr, P.S.; Sebastian, S.A. Biochemical and molecular characterization of a mutation that confers a decreased raffinosaccharide and phytic acid phenotype on soybean seeds. Plant Physiol. 2002, 128, 650–660. [Google Scholar] [CrossRef] [PubMed]
- Wilcox, J.R.; Premachandra, G.S.; Young, K.A.; Raboy, V. Isolation of high seed inorganic P, low-phytate soybean mutants. Crop Sci. 2000, 40, 1601–1605. [Google Scholar] [CrossRef]
- Yuan, F.J.; Zhao, H.J.; Ren, X.L.; Zhu, S.L.; Fu, X.J.; Shu, Q.Y. Generation and characterization of two novel low phytate mutations in soybean (Glycine max L. Merr.). Theor. Appl. Genet. 2007, 115, 945–957. [Google Scholar] [CrossRef] [PubMed]
- Dorsch, A.J.; Cook, A.; Young, A.K.; Anderson, N.J.; Bauman, T.A.; Volkmann, J.C.; Murthy, P.N.P.; Raboy, V. Seed phosphorus and inositol phosphate phenotype of barley low phytic acid genotypes. Phytochemistry 2003, 62, 691–706. [Google Scholar] [CrossRef]
- Haug, W.; Lantzsch, H.-J. Sensitive method for the rapid determination of phytate in cereals and cereal products. J. Sci. Food Agric. 1983, 34, 1423–1426. [Google Scholar] [CrossRef]
© 2015 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Raboy, V.; Peterson, K.; Jackson, C.; Marshall, J.M.; Hu, G.; Saneoka, H.; Bregitzer, P. A Substantial Fraction of Barley (Hordeum vulgare L.) Low Phytic Acid Mutations Have Little or No Effect on Yield across Diverse Production Environments. Plants 2015, 4, 225-239. https://0-doi-org.brum.beds.ac.uk/10.3390/plants4020225
Raboy V, Peterson K, Jackson C, Marshall JM, Hu G, Saneoka H, Bregitzer P. A Substantial Fraction of Barley (Hordeum vulgare L.) Low Phytic Acid Mutations Have Little or No Effect on Yield across Diverse Production Environments. Plants. 2015; 4(2):225-239. https://0-doi-org.brum.beds.ac.uk/10.3390/plants4020225
Chicago/Turabian StyleRaboy, Victor, Kevin Peterson, Chad Jackson, Juliet M. Marshall, Gongshe Hu, Hirofumi Saneoka, and Phil Bregitzer. 2015. "A Substantial Fraction of Barley (Hordeum vulgare L.) Low Phytic Acid Mutations Have Little or No Effect on Yield across Diverse Production Environments" Plants 4, no. 2: 225-239. https://0-doi-org.brum.beds.ac.uk/10.3390/plants4020225