
Effects of PEG-Induced Water Deficit in Solanum nigrum on Zn and Ni Uptake and Translocation in Split Root Systems



{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
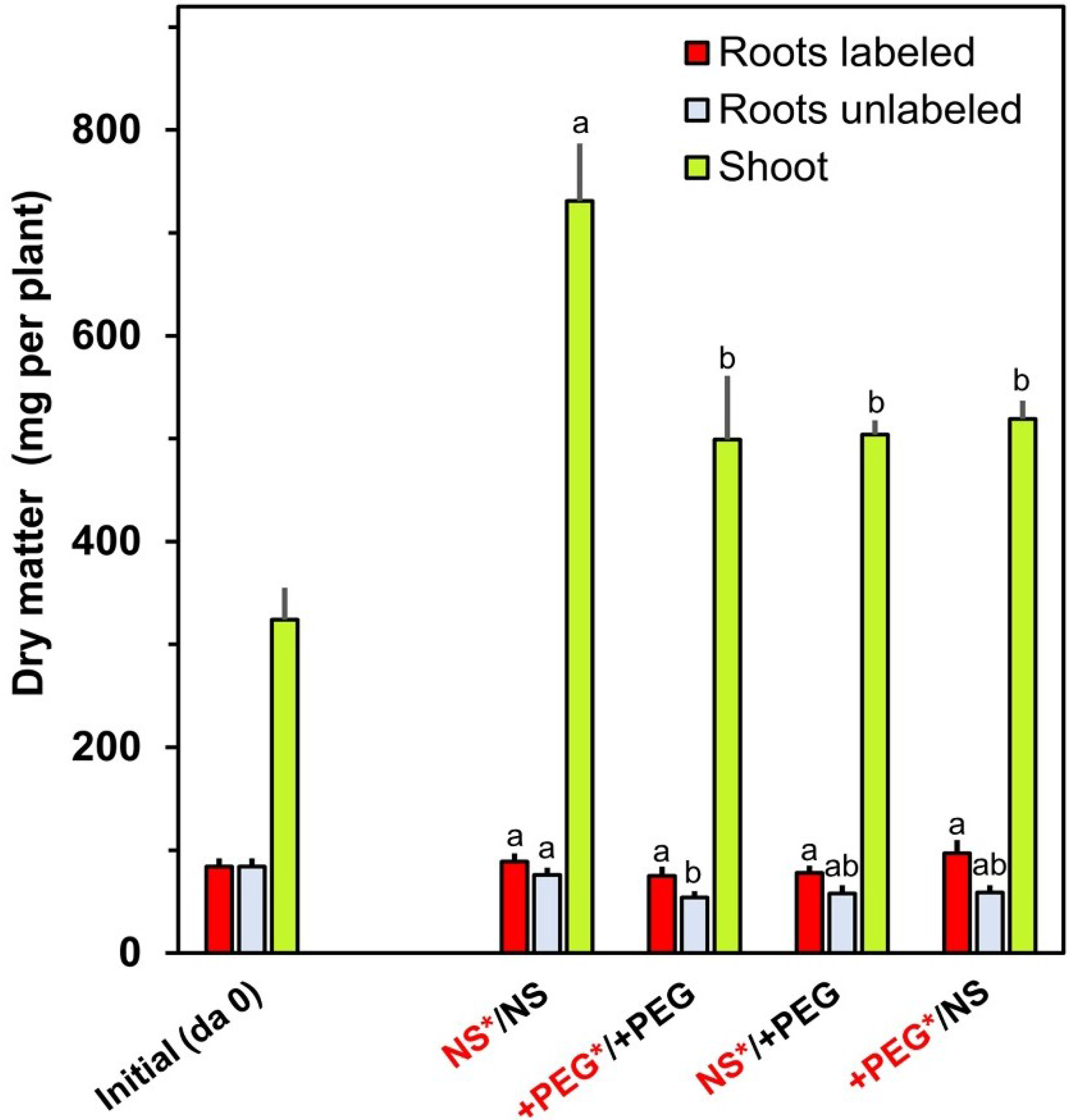
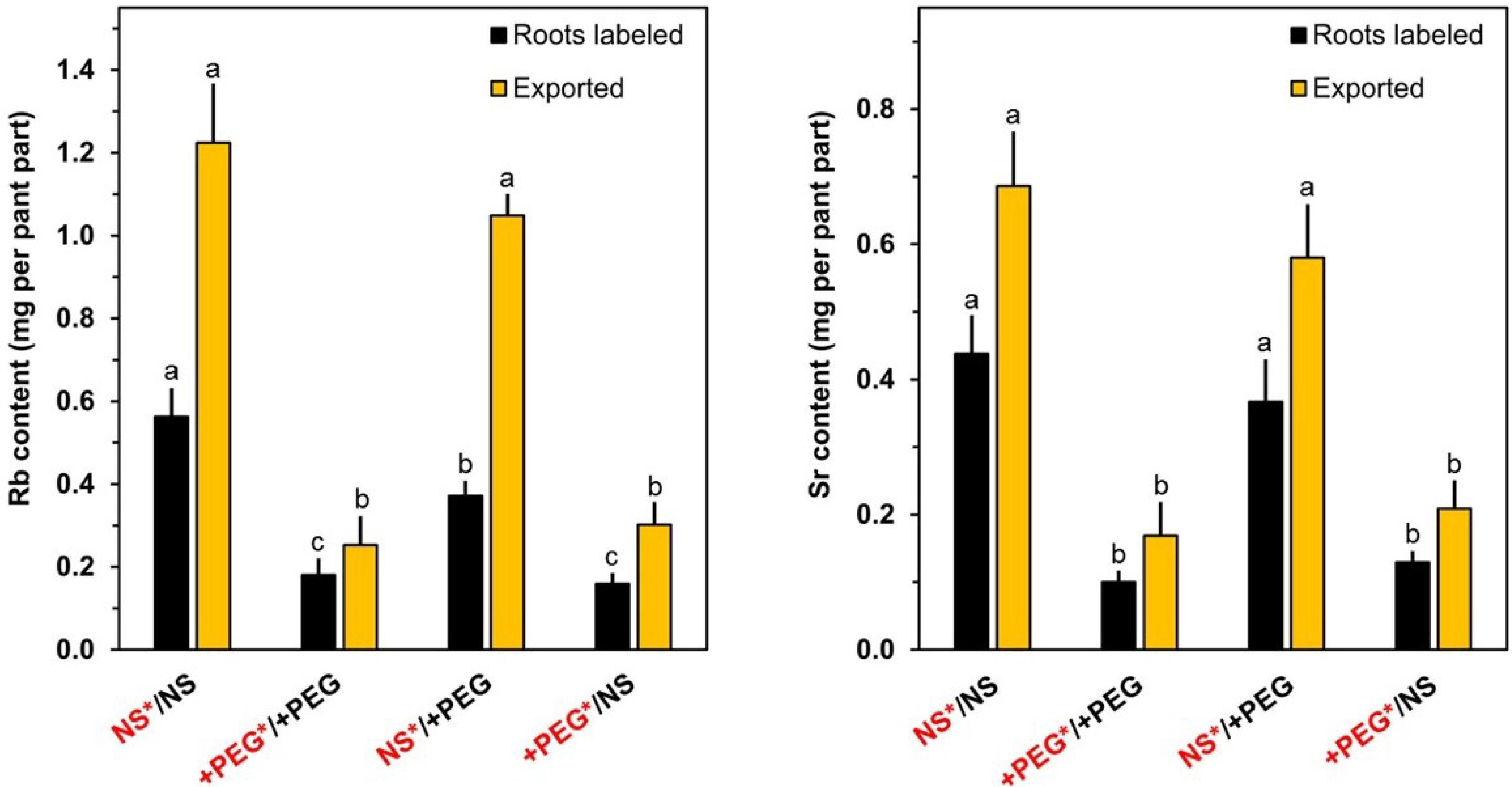
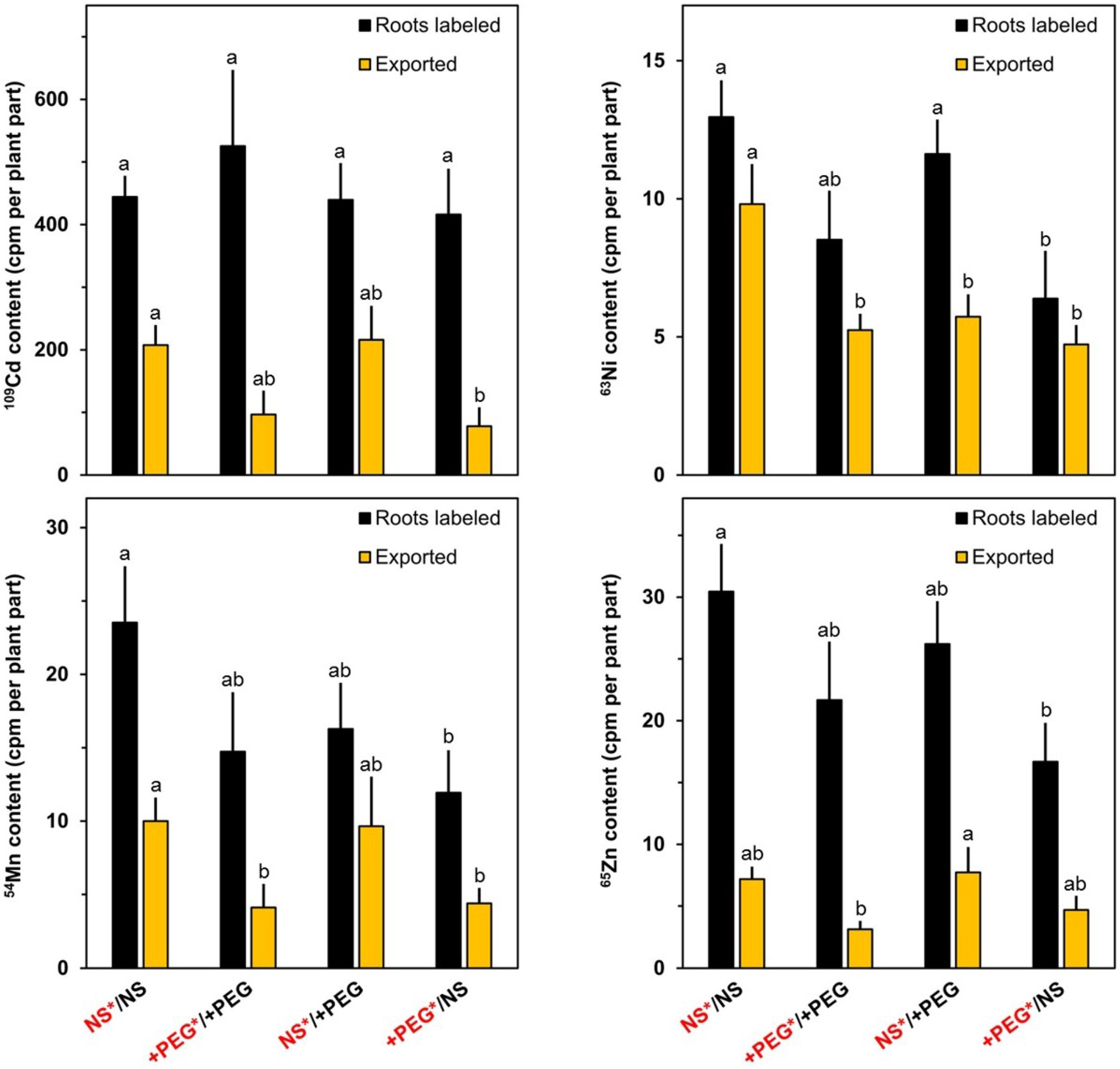
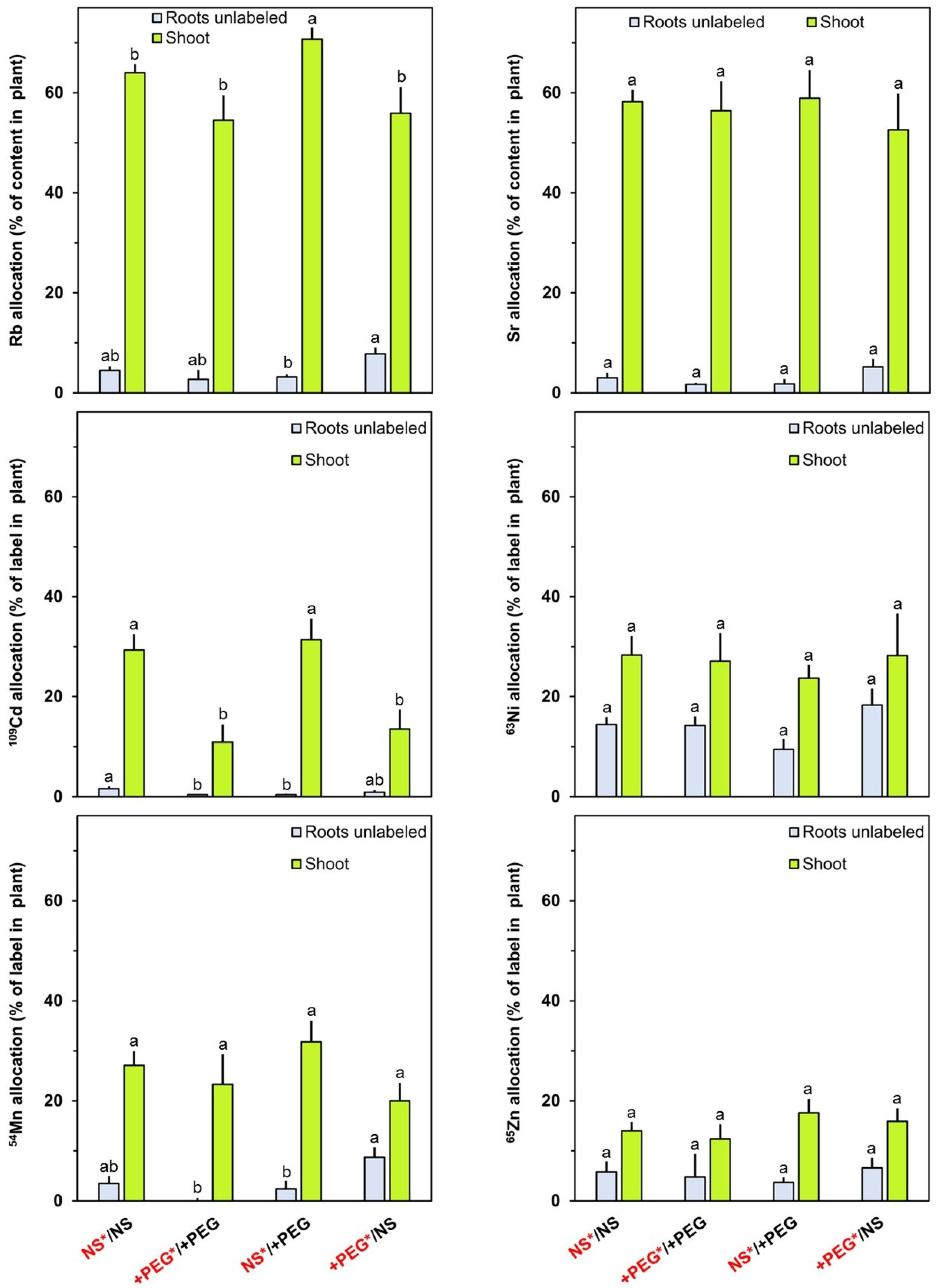
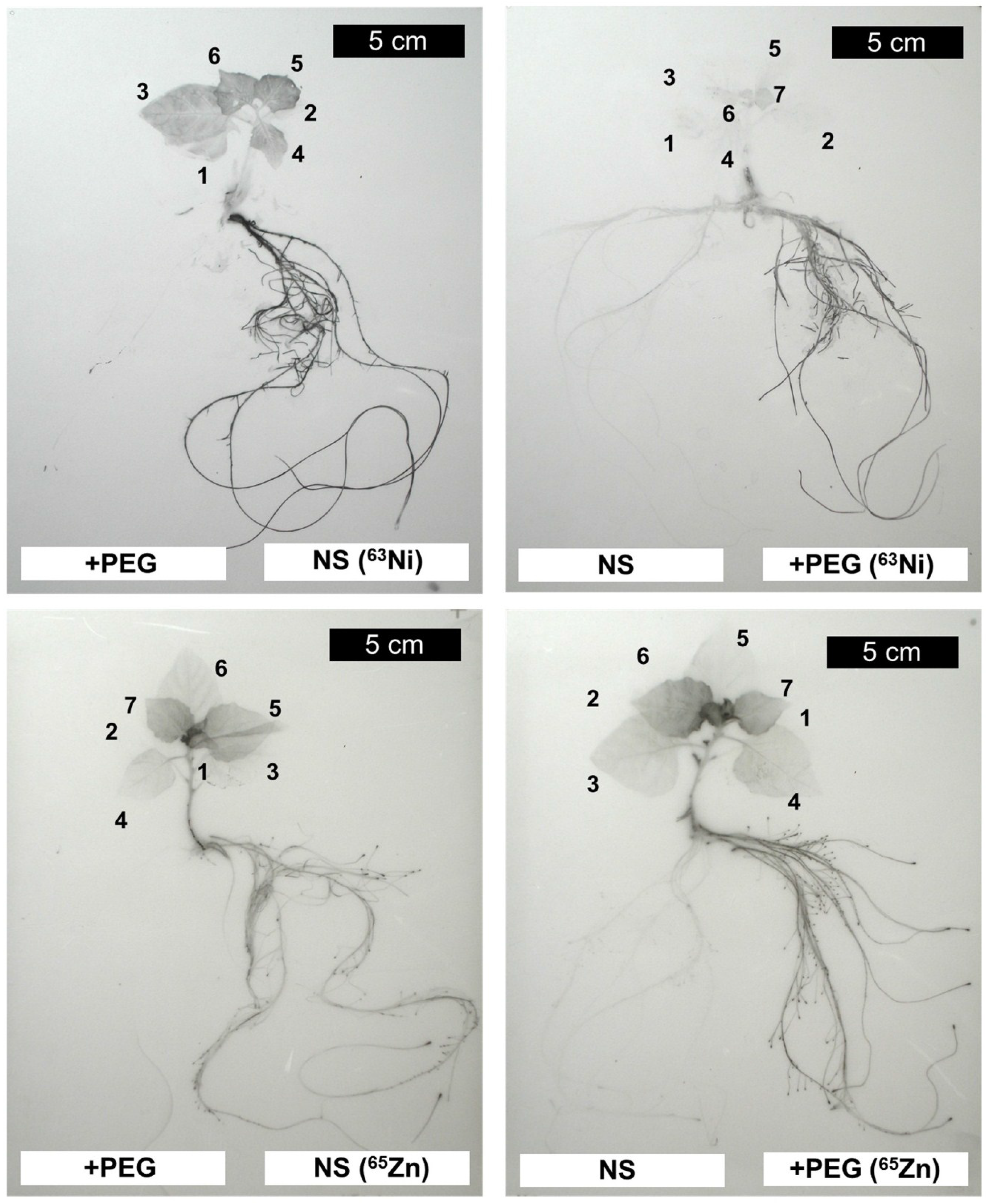
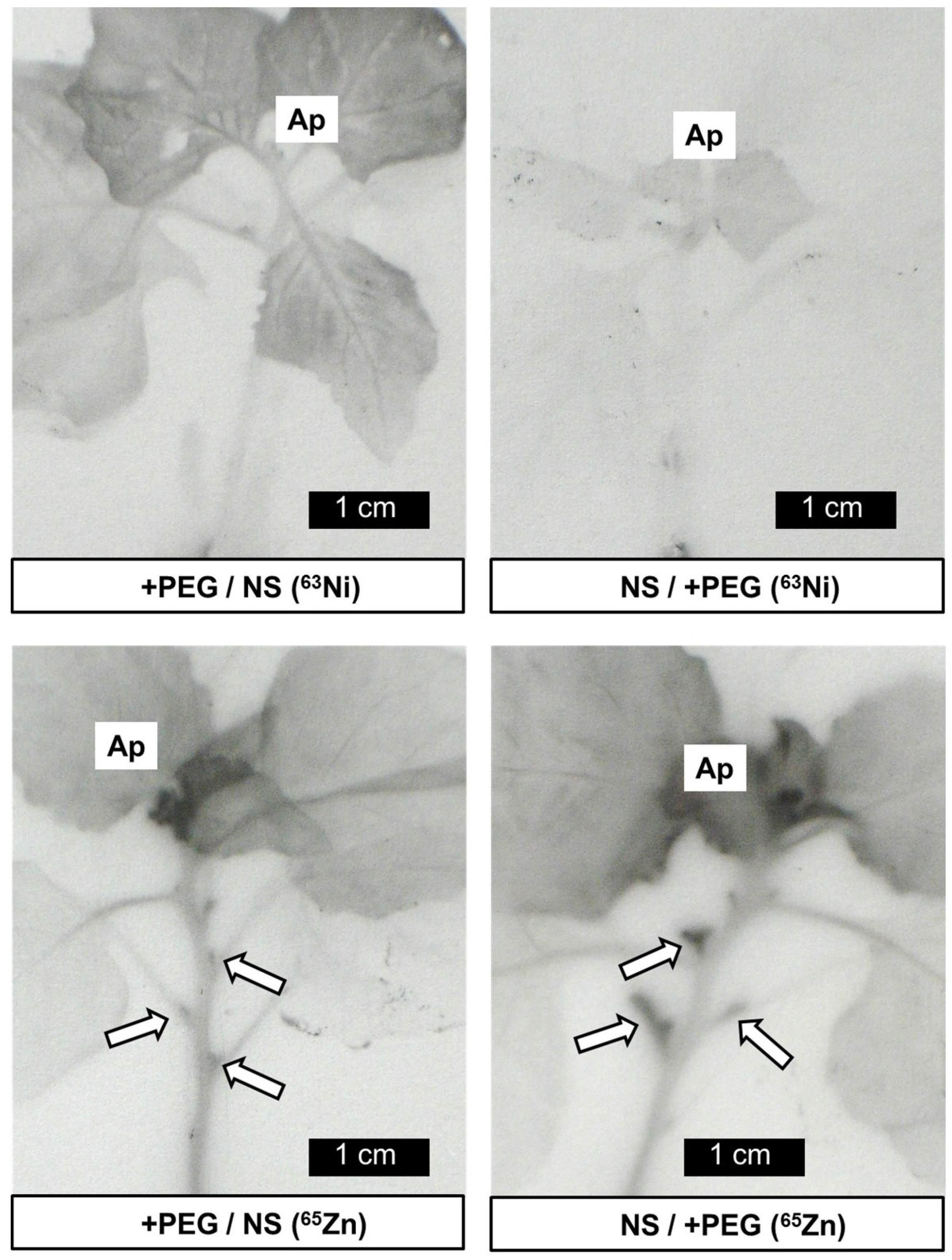
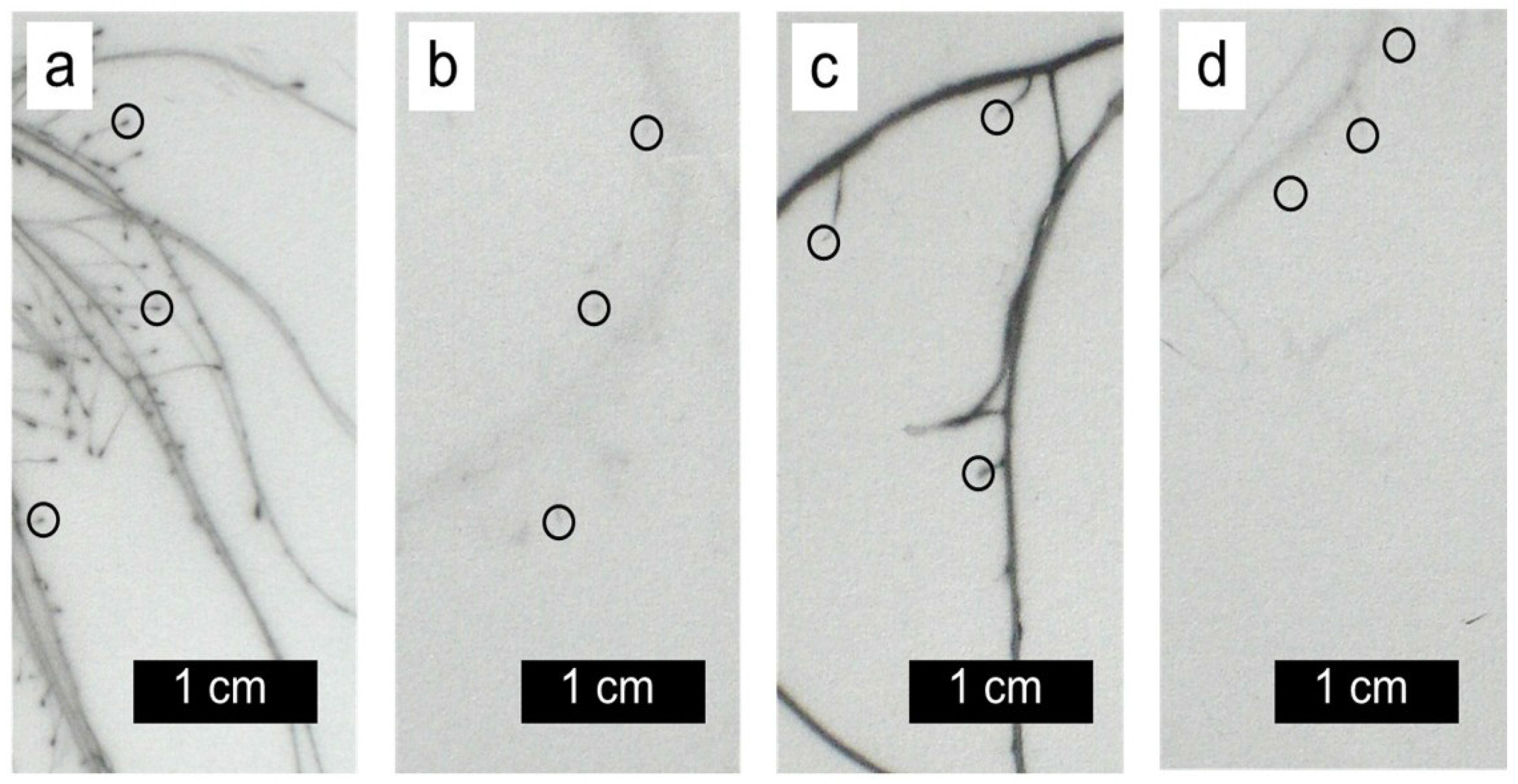
2. Results and Discussion







3. Materials and Methods
3.1. Plant Material and Culture Conditions
3.2. Experimental Setup and Quantification of Labels
3.3. Autoradiography and Localization of 63Ni and 65Zn
4. Conclusions
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Cantarel, A.A.M.; Bloor, J.M.G.; Sousanna, J.-F. Four years of simulated climate change reduces above-ground productivity and alters functional diversity in a grassland ecosystem. J. Veg. Sci. 2013, 24, 113–126. [Google Scholar] [CrossRef]
- Burri, S.; Sturm, P.; Prechsl, U.E.; Knohl, A.; Buchmann, N. The impact of extreme summer drought on the short-term carbon coupling of photosynthesis to soil CO2 efflux in a temperate grassland. Biogeosciences 2014, 11, 961–975. [Google Scholar] [CrossRef]
- Gilgen, A.K.; Signarbieux, C.; Feller, U.; Buchmann, N. Competitive advantage of Rumex obtusifolius L. might increase in intensively managed temperate grasslands under dryer climate. Agric. Ecosyst. Environ. 2010, 135, 15–23. [Google Scholar] [CrossRef] [Green Version]
- Van den Berge, J.; Naudts, K.; de Boeck, H.J.; Ceulemans, R.; Nijs, I. Do interactions with neighbours modify the above-ground productivity response to drought? A test with two grassland species. Environ. Exp. Bot. 2014, 105, 18–24. [Google Scholar] [CrossRef]
- Grant, K.; Kreyling, J.; Heilmeier, H.; Beierkuhnlein, C.; Jentsch, A. Extreme weather events and plant-plant interactions: Shifts between competition and facilitation among grassland species in the face of drought and heavy rainfall. Ecol. Res. 2014, 29, 991–1001. [Google Scholar] [CrossRef]
- Khan, M.A.A.; Grant, K.; Beierkuhnlein, C.; Kreyling, J.; Jentsch, A. Climatic extremes lead to species-specific legume facilitation in an experimental temperate grassland. Plant Soil 2014, 379, 161–175. [Google Scholar] [CrossRef]
- Bollig, C.; Feller, U. Impacts of drought stress on water relations and carbon assimilation in grassland species at different altitudes. Agric. Ecosyst. Environ. 2014, 188, 212–220. [Google Scholar] [CrossRef]
- Signarbieux, C.; Feller, U. Effects of an extended drought period on physiological properties of grassland species in the field. J. Plant Res. 2012, 125, 251–261. [Google Scholar] [CrossRef] [PubMed]
- Marschner, H. Mineral Nutrition of Higher Plants, 2nd ed.; Academic Press: London, UK, 1995; p. 38. [Google Scholar]
- Vallejo, A.J.; Peralta, M.L.; Santa-Maria, G.E. Expression of potassium-transporter coding genes, and kinetics of rubidium uptake, along a longitudinal root axis. Plant Cell Environ. 2005, 28, 850–862. [Google Scholar] [CrossRef]
- Herren, T.; Feller, U. Transport of cadmium via xylem and phloem in maturing wheat shoots: Comparison with the translocation of zinc, strontium and rubidium. Ann. Bot. 1997, 80, 623–628. [Google Scholar] [CrossRef]
- Tyson, M.J.; Sheffield, E.; Callaghan, T.V. Uptake, allocation, accumulation and ecological implications of Sr-85 in bracken (Pteridium aquilinum L.-Kuhn). J. Environ. Radioact. 1999, 46, 15–25. [Google Scholar] [CrossRef]
- Seregin, I.V.; Kozhevnikova, A.D. Strontium transport, distribution, and toxic effects on maize seedling growth. Russ. J. Plant Physiol. 2004, 51, 215–221. [Google Scholar] [CrossRef]
- Page, V.; Feller, U. Selective transport of zinc, manganese, nickel, cobalt and cadmium in the root system and transfer to the leaves in young wheat plants. Ann. Bot. 2005, 96, 425–434. [Google Scholar] [CrossRef] [PubMed]
- Riesen, O.; Feller, U. Redistribution of nickel, cobalt, manganese, zinc and cadmium via the phloem in young and maturing wheat. J. Plant Nutr. 2005, 28, 421–430. [Google Scholar] [CrossRef]
- Page, V.; Weisskopf, L.; Feller, U. Heavy metals in white lupin: Uptake, root-to-shoot transfer and redistribution within the plant. New Phytol. 2006, 171, 329–341. [Google Scholar] [CrossRef] [PubMed]
- Wei, S.; Anders, I.; Feller, U. Selective uptake, distribution, and redistribution of Cd-109, Co-57, Zn-65, Ni-63, and Cs-134 via xylem and phloem in the heavy metal hyperaccumulator Solanum nigrum L. Environ. Sci. Pollut. Res. 2014, 21, 7624–7630. [Google Scholar] [CrossRef] [PubMed]
- Volaire, F. Drought survival, summer dormancy and dehydrin accumulation in contrasting cultivars of Dactylis glomerata. Physiol. Plant. 2002, 116, 42–51. [Google Scholar] [CrossRef] [PubMed]
- Simova-Stoilova, L.; Demirevska, K.; Petrova, T.; Tsenov, N.; Feller, U. Antioxidative protection and proteolytic activity in tolerant and sensitive wheat (Triticum aestivum L.) varieties subjected to long-term field drought. Plant Growth Regul. 2009, 58, 107–117. [Google Scholar] [CrossRef]
- Plaut, Z.; Butow, B.J.; Blumenthal, C.S.; Wrigley, C.W. Transport of dry matter into developing wheat kernels and its contribution to grain yield under post-anthesis water deficit and elevated temperature. Field Crops Res. 2004, 86, 185–198. [Google Scholar] [CrossRef]
- Bencze, S.; Bamberger, Z.; Janda, T.; Balla, K.; Varga, B.; Bedo, Z.; Veisz, O. Physiological response of wheat to elevated atmospheric CO2 and low water supply levels. Photosynthetica 2014, 52, 71–82. [Google Scholar] [CrossRef]
- Demirevska, K.; Simova-Stoilova, L.; Vassileva, V.; Feller, U. Rubisco and some chaperone protein responses to water stress and rewatering at early seedling growth of drought sensitive and tolerant wheat varieties. Plant Growth Regul. 2008, 56, 97–106. [Google Scholar] [CrossRef]
- Grigorova, B.; Vaseva, I.; Demirevska, K.; Feller, U. Combined drought and heat stress in wheat: Changes in some heat shock proteins. Biol. Plant. 2011, 55, 105–111. [Google Scholar] [CrossRef]
- Vassileva, V.; Simova-Stoilova, L.; Demirevska, K.; Feller, U. Variety-specific response of wheat (Triticum aestivum L.) leaf mitochondria to drought stress. J. Plant Res. 2009, 122, 445–454. [Google Scholar] [CrossRef] [PubMed]
- Vassileva, V.; Signarbieux, C.; Anders, I.; Feller, U. Genotypic variation in drought stress response and subsequent recovery of (Triticum aestivum L.). J. Plant Res. 2011, 124, 147–154. [Google Scholar] [CrossRef] [PubMed]
- Cougnon, M.; Baert, J.; van Waes, C.; Reheul, D. Performance and quality of of tall fescue (Festuca arundinacea Schreb.) and perennial ryegrass (Lolium perenne L.) and mixtures of both species grown with or without white clover (Trifolium repens L.) under cutting management. Grass Forage Sci. 2014, 69, 666–677. [Google Scholar] [CrossRef] [Green Version]
- Turner, L.R.; Holloway-Phillips, M.M.; Rawnsley, R.P.; Donaghy, D.J.; Pembleton, K.G. The morphological and physiological responses of perennial ryegrass (Lolium perenne L.), cocksfoot (Dactylis glomerata L.) and tall fescue (Festuca arundinacea Schreb.; syn. Schoedonorus phoenix Scop.) to variable water availability. Grass Forage Sci. 2012, 67, 507–518. [Google Scholar] [CrossRef]
- Feller, U.; Vaseva, I.I. Extreme climatic events: Impacts of drought and high temperature on physiological processes in agronomically important plants. Front. Environ. Sci. 2014, 2, 39. [Google Scholar] [CrossRef]
- Gilgen, A.K.; Feller, U. Drought stress alters solute allocation in broadleaf dock (Rumex obtusifolius). Weed Sci. 2013, 61, 104–108. [Google Scholar] [CrossRef]
- Wei, S.; Zhou, Q.X.; Koval, P.V. Flowering stage characteristics of cadmium hyperaccumulator Solanum nigrum L. and their significance to phytoremediation. Sci. Total Environ. 2005, 369, 441–446. [Google Scholar] [CrossRef] [PubMed]
- Yang, X.E.; Long, X.X.; Ye, Z.L.; Calvert, D.V.; Stoffella, P.J. Cadmium tolerance and hyperaccumulation in a new Zn-hyperaccumulator (Sedum alfredii Hance). Plant Soil 2004, 259, 181–189. [Google Scholar] [CrossRef]
- Brown, S.L.; Chaney, R.L.; Angle, J.S.; Baker, A.J.M. Zinc and cadmium uptake by hyperaccumulator Thlaspi caerulescens grown in nutrient solution. Soil Sci. Soc. Am. J. 1995, 59, 125–133. [Google Scholar] [CrossRef]
- Chaney, R.L.; Malik, M.; Li, Y.M.; Brown, S.L.; Brewer, E.P.; Angle, J.S.; Baker, A.J.M. Phytoremediation of soil metals. Curr. Opin. Biotechnol. 1997, 8, 279–284. [Google Scholar] [CrossRef]
- Wei, S.; Zhou, Q.X.; Wang, X.; Zhang, K.S.; Guo, G.L.; Ma, Q.Y.L. A newly-discovered Cd-hyperaccumulator Solanum nigrum L. Chin. Sci. Bull. 2005, 50, 33–38. [Google Scholar] [CrossRef]
- Hänsch, R.; Mendel, R.R. Physiological functions of mineral micronutrients (Cu, Zn, Mn, Fe, Ni, Mo, B, Cl). Curr. Opin. Plant Biol. 2009, 12, 259–266. [Google Scholar] [CrossRef] [PubMed]
- Gerendas, J.; Polacco, J.C.; Freyermuth, S.K.; Sattelmacher, B. Significance of nickel for plant growth and metabolism. J. Plant Nutr Soil Sci.-Z. Pflanzenernähr. Bodenk. 1999, 162, 241–256. [Google Scholar] [CrossRef]
- Bai, C.; Liu, L.P.; Wood, B.W. Nickel affects xylem sap RNase and converts RNase A to a urease. BMC Plant Biol. 2013, 13, 207. [Google Scholar] [CrossRef] [PubMed]
- Gowing, D.J.G.; Davies, W.J.; Jones, H.G. A positive root-sourced signal as an indicator of soil drying in apple, Malus x domestica Borkh. J. Exp. Bot. 1990, 41, 1535–1540. [Google Scholar] [CrossRef]
- Steuter, A.A.; Mozafar, A.; Goodin, J.R. Water potential of aqueous polyethylene glycol. Plant Physiol. 1981. [Google Scholar] [CrossRef]
- Michel, B.E.; Kaufmann, M.R. The osmotic potential of polyethylene glycol 6000. Plant Physiol. 1973, 51, 914–916. [Google Scholar] [CrossRef] [PubMed]
- Fahn, A. Plant Anatomy, 3rd ed.; Pergamon Press: Oxford, UK, 1982; pp. 46–73. [Google Scholar]
© 2015 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Feller, U.; Anders, I.; Wei, S. Effects of PEG-Induced Water Deficit in Solanum nigrum on Zn and Ni Uptake and Translocation in Split Root Systems. Plants 2015, 4, 284-297. https://0-doi-org.brum.beds.ac.uk/10.3390/plants4020284
Feller U, Anders I, Wei S. Effects of PEG-Induced Water Deficit in Solanum nigrum on Zn and Ni Uptake and Translocation in Split Root Systems. Plants. 2015; 4(2):284-297. https://0-doi-org.brum.beds.ac.uk/10.3390/plants4020284
Chicago/Turabian StyleFeller, Urs, Iwona Anders, and Shuhe Wei. 2015. "Effects of PEG-Induced Water Deficit in Solanum nigrum on Zn and Ni Uptake and Translocation in Split Root Systems" Plants 4, no. 2: 284-297. https://0-doi-org.brum.beds.ac.uk/10.3390/plants4020284