GWAS for Starch-Related Parameters in Japonica Rice (Oryza sativa L.)


 , , ,
, , ,  , , and
, , and
Abstract
:1. Introduction
2. Results
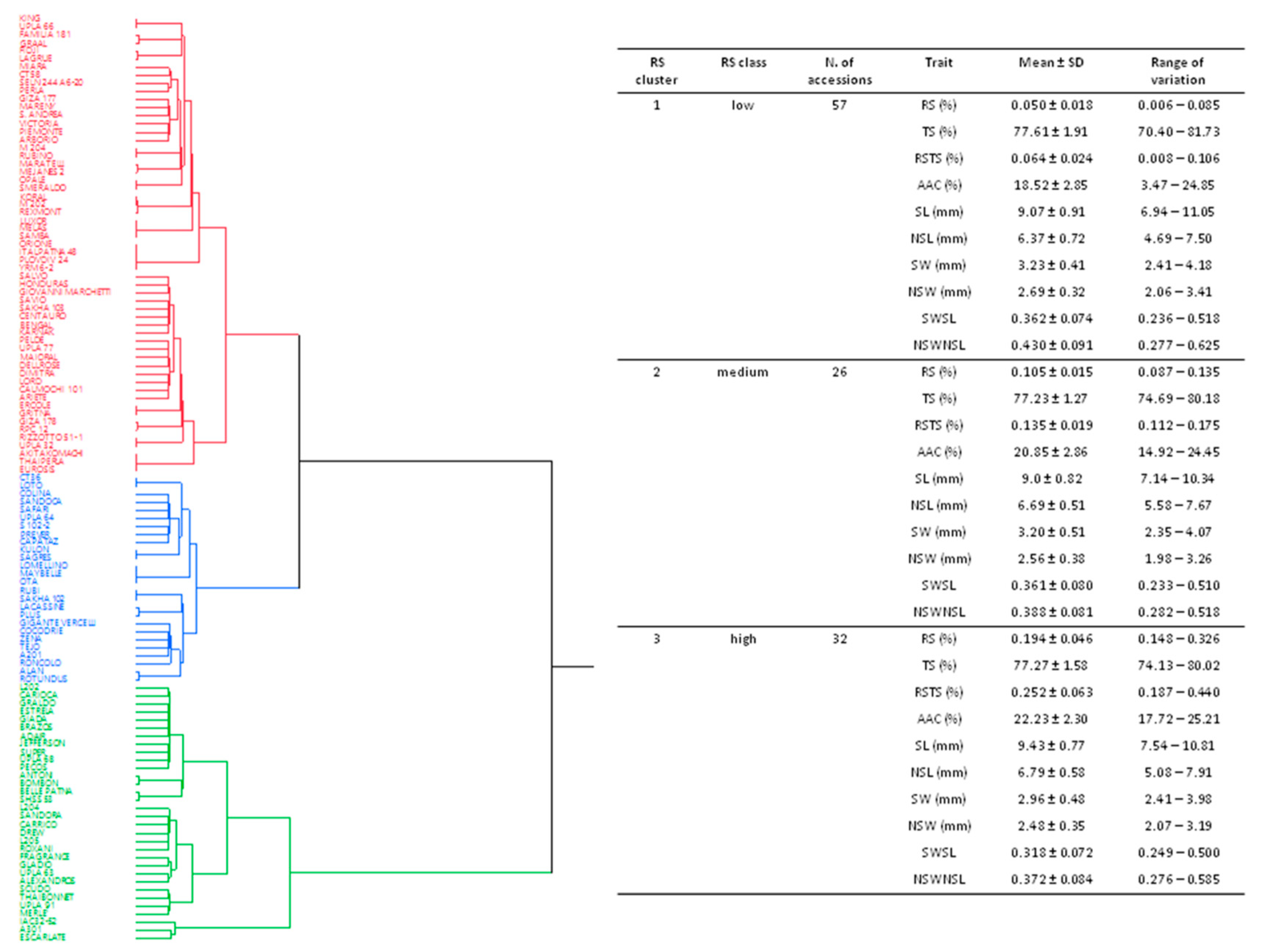
2.1. Assessment of Starch-Related Parameters in the Japonica Rice Population
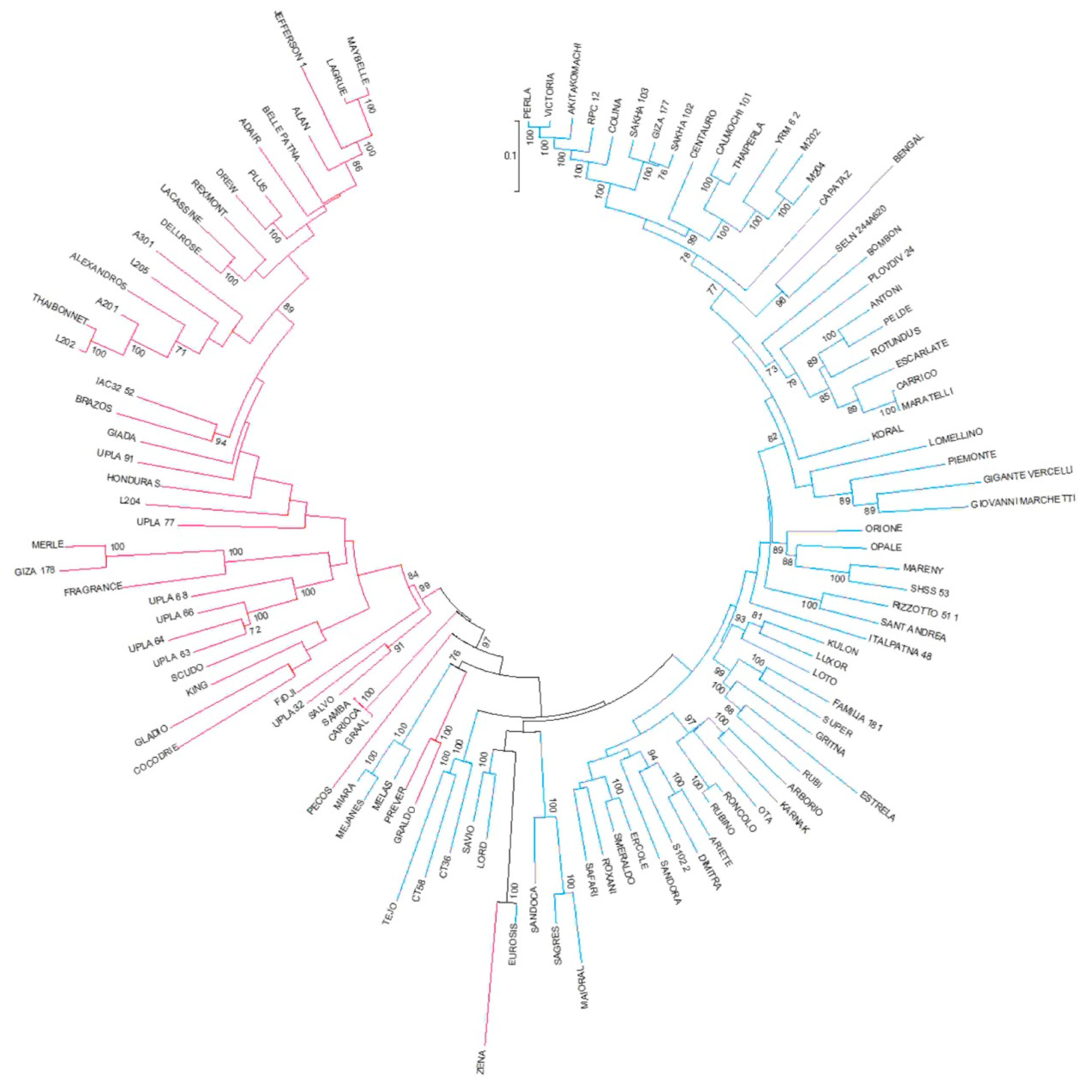
2.2. Genotypic Analysis of the Japonica Rice Population
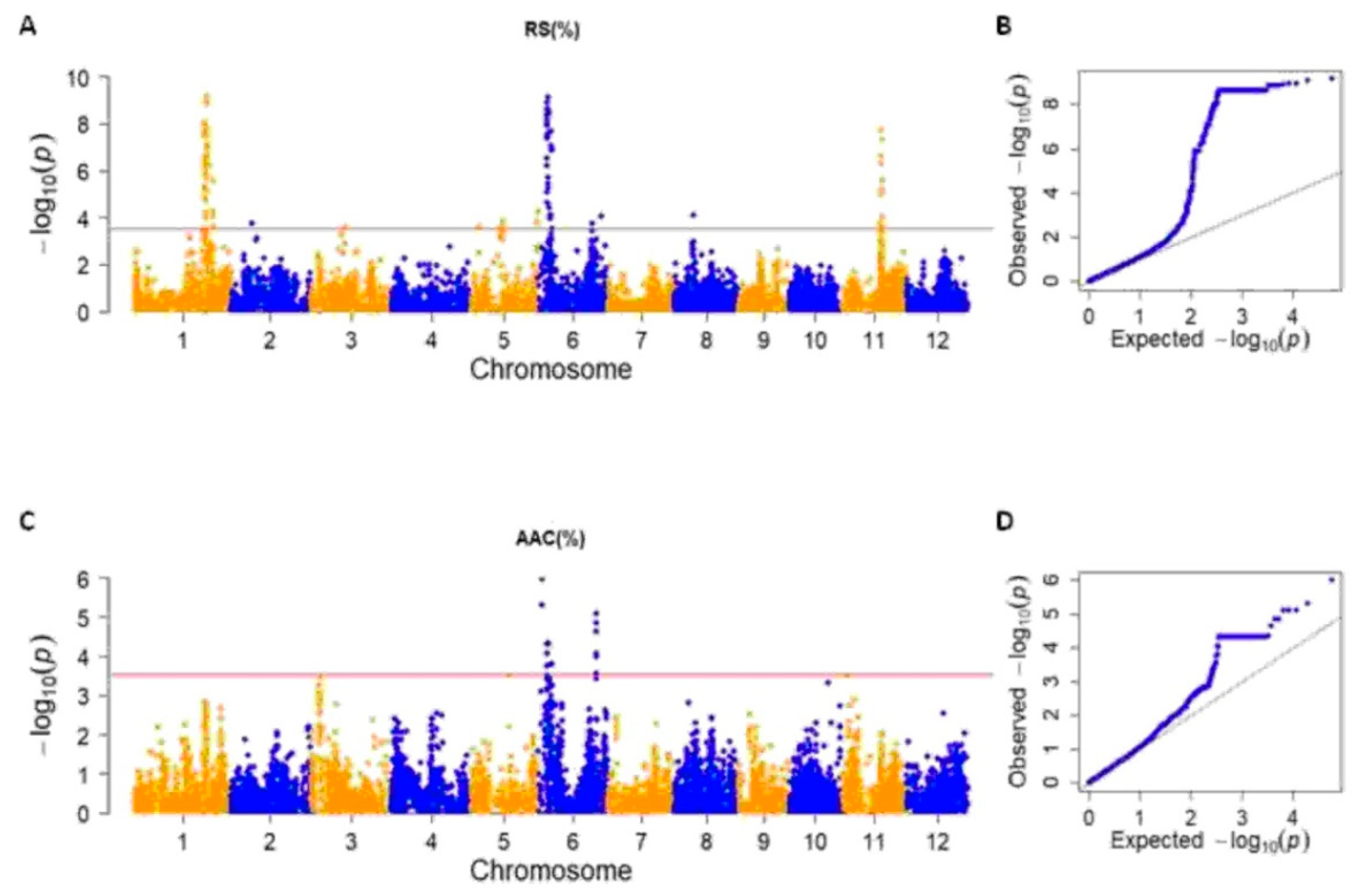
2.3. Genome Wide Association Analysis for RS, AAC and TS
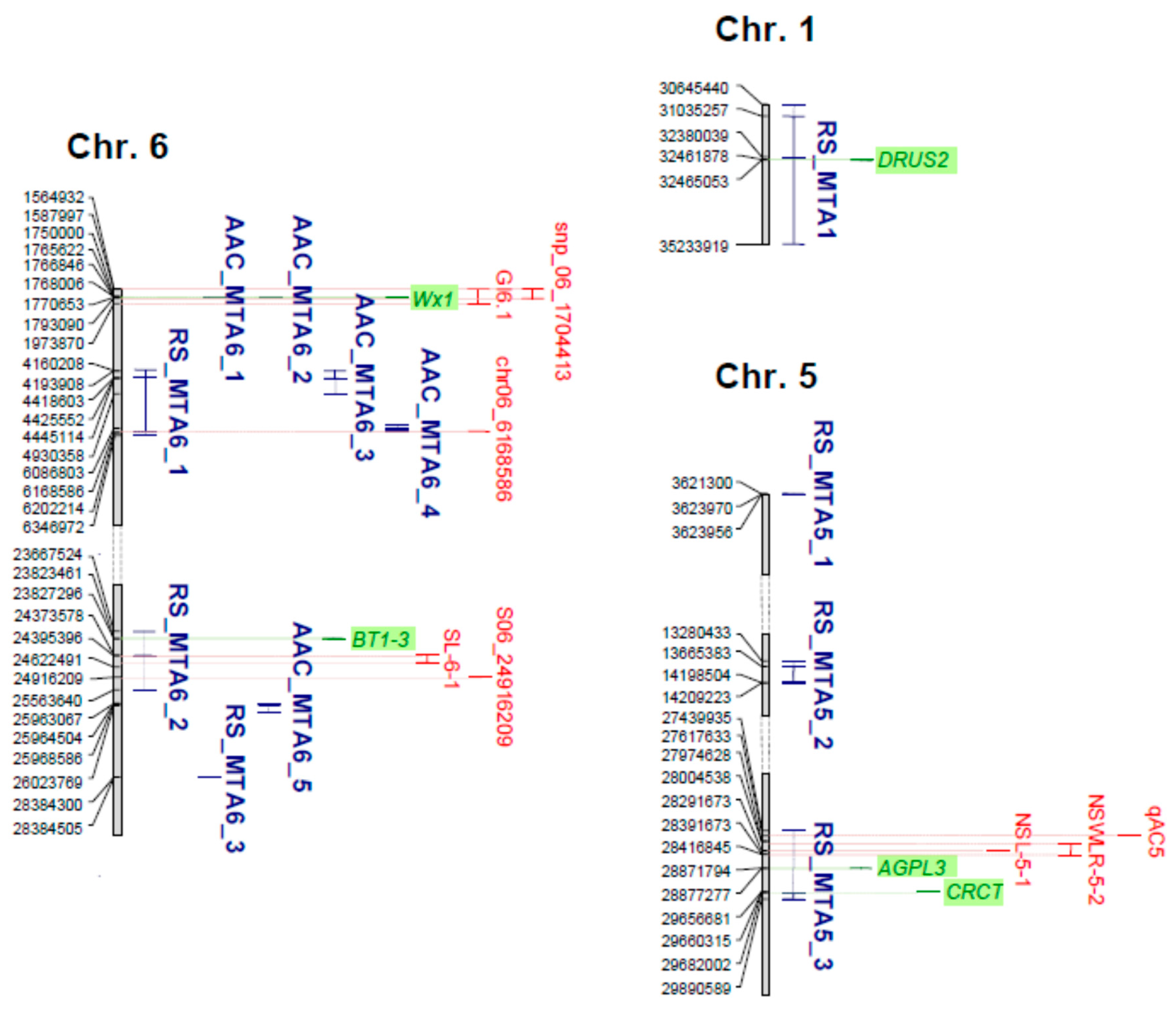
2.4. Identification of Candidate Genes
3. Discussion
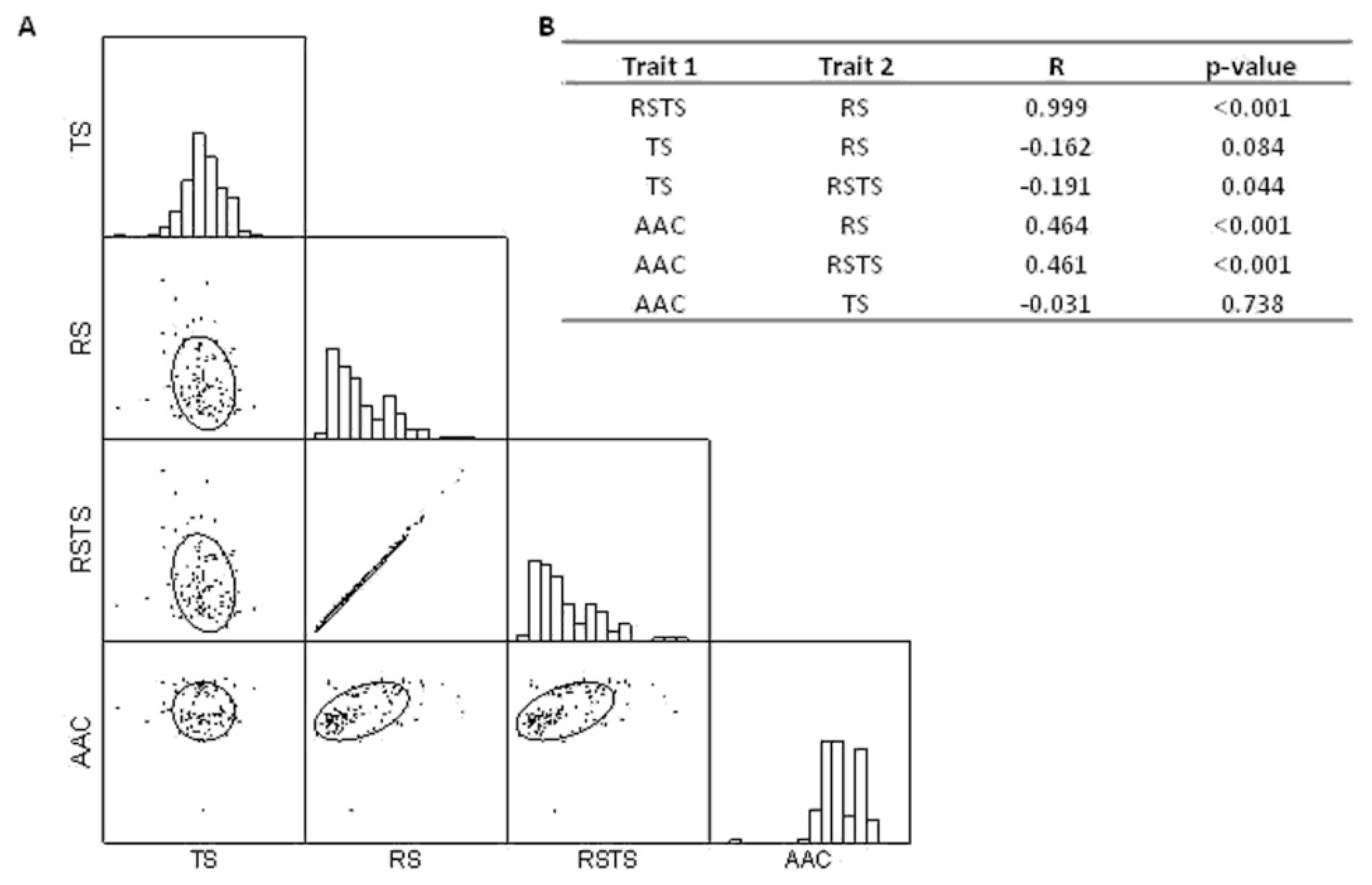
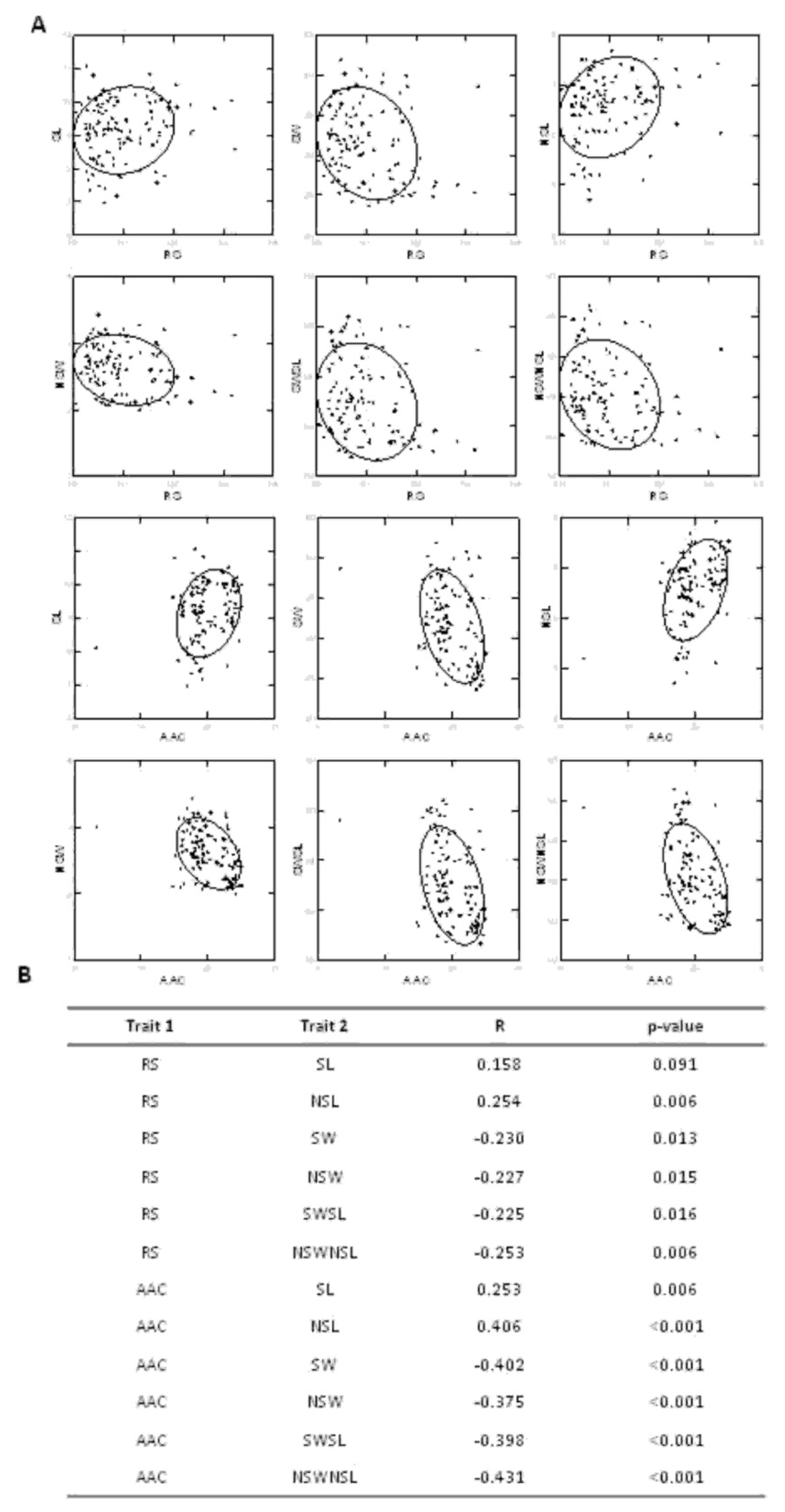
3.1. Correlation Between Grain Quality Related Traits
3.2. GWAS Analyses for RS and AAC and Identification of Candidate Genes
4. Materials and Methods
4.1. Plant Materials
4.2. Phenotypic Analyses
4.3. Statistical Analyses of Phenotypic Data
4.4. Genotyping
4.5. Population Structure and Linkage Disequilibrium Decay
4.6. Association Mapping
4.7. Identification of Candidate Genes Affecting RS and/or AAC
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Conflicts of Interest
Abbreviations
| AAC | apparent amylose content |
| AGPase | ADP-glucose pyrosphorylase |
| DBE | starch debranching enzyme |
| GBS | genotyping by sequencing |
| GBSSI | granule bound starch synthase I |
| GWAS | Genome Wide Association Scan |
| MTA | marker-trait association |
| QTL | quantitative trait locus |
| RS | resistant starch |
| SBE | starch branching enzyme |
| SS | starch synthase |
| TS | total starch |
| Wx | Waxy |
References
- Biselli, C.; Cavalluzzo, D.; Perrini, R.; Gianinetti, A.; Bagnaresi, P.; Urso, S.; Orasen, G.; Desiderio, F.; Lupotto, E.; Cattivelli, L.; et al. Improvement of marker-based predictability of Apparent Amylose Content in japonica rice through GBSSI allele mining. Rice 2014, 7, 1. [Google Scholar] [CrossRef] [PubMed]
- Juliano, B.O. Structure and function of the rice grain and its fractions. Cereal Foods World 1992, 37, 772–774. [Google Scholar]
- Suwannaporn, P.; Pitiphunpong, S.; Champangern, S. Classification of Rice Amylose Content by Discriminant Analysis of Physicochemical Properties. Starch 2007, 59, 171–177. [Google Scholar] [CrossRef]
- Mikami, I.; Uwatoko, N.; Ikeda, Y.; Yamaguchi, J.; Hirano, H.-Y.; Suzuki, Y.; Sano, Y. Allelic diversification at the wx locus in landraces of Asian rice. Theor. Appl. Genet. 2008, 116, 979–989. [Google Scholar] [CrossRef] [PubMed]
- Bao, J.; Corke, H.; Sun, M. Analysis of Genetic Diversity and Relationships in Waxy Rice (Oryza sativa L.) using AFLP and ISSR Markers. Genet. Resour. Crop. Evol. 2006, 53, 323–330. [Google Scholar] [CrossRef]
- Chen, M.-H.; Bergman, C.; Pinson, S.; Fjellstrom, R. Waxy gene haplotypes: Associations with apparent amylose content and the effect by the environment in an international rice germplasm collection. J. Cereal Sci. 2008, 47, 536–545. [Google Scholar] [CrossRef]
- Teng, B.; Zhang, Y.; Wu, J.; Cong, X.; Wang, R.; Han, Y.; Luo, Z. Association between allelic variation at theWaxylocus and starch physicochemical properties using single-segment substitution lines in rice (Oryza sativa L.). Starch 2013, 65, 1069–1077. [Google Scholar] [CrossRef]
- Dobo, M.; Ayres, N.; Walker, G.; Park, W.D. Polymorphism in the GBSS gene affects amylose content in US and European rice germplasm. J. Cereal Sci. 2010, 52, 450–456. [Google Scholar] [CrossRef]
- Englyst, H.N.; Cummings, J.H. Digestion of the polysaccharides of some cereal foods in the human small intestine. Am. J. Clin. Nutr. 1985, 42, 778–787. [Google Scholar] [CrossRef]
- Robertson, M.D.; Currie, J.M.; Morgan, L.M.; Jewell, D.P.; Frayn, K.N. Prior short-term consumption of resistant starch enhances postprandial insulin sensitivity in healthy subjects. Diabetologia 2003, 46, 659–665. [Google Scholar] [CrossRef] [Green Version]
- Yamada, Y.; Hosoya, S.; Nishimura, S.; Tanaka, T.; Kajimoto, Y.; Nishimura, A.; Kajimoto, O. Effect of Bread Containing Resistant Starch on Postprandial Blood Glucose Levels in Humans. Biosci. Biotechnol. Biochem. 2005, 69, 559–566. [Google Scholar] [CrossRef] [PubMed]
- Brouns, F.; Arrigoni, E.; Langkilde, A.M.; Verkooijen, I.; Fässler, C.; Andersson, H.; Kettlitz, B.; Van Nieuwenhoven, M.; Philipsson, H.; Amadò, R. Physiological and Metabolic Properties of a Digestion-Resistant Maltodextrin, Classified as Type 3 Retrograded Resistant Starch. J. Agric. Food Chem. 2007, 55, 1574–1581. [Google Scholar] [CrossRef] [PubMed]
- Haub, M.D.; Hubach, K.L.; Al-Tamimi, E.K.; Ornelas, S.; Seib, P.A. Different Types of Resistant Starch Elicit Different Glucose Reponses in Humans. J. Nutr. Metab. 2010, 2010, 230501. [Google Scholar] [CrossRef] [PubMed]
- Zenel, A.M.; Stewart, M.L. High Amylose White Rice Reduces Post-Prandial Glycemic Response but Not Appetite in Humans. Nutrients 2015, 7, 5362–5374. [Google Scholar] [CrossRef] [PubMed]
- Dronamraju, S.S.; Coxhead, J.M.; Kelly, S.B.; Mathers, J.C. Role of Resistant Starch in Colorectal Cancer Prevention: A Prospective Randomised Controlled Trial. Am. J. Gastroenterol. 2007, 102, S556–S557. [Google Scholar] [CrossRef]
- Tapsell, L.C. Diet and metabolic syndrome: Where does resistant starch fit in? J. AOAC Int. 2004, 87, 756–760. [Google Scholar] [PubMed]
- Willis, H.J.; Eldridge, A.L.; Beiseigel, J.; Thomas, W.; Slavin, J.L. Greater satiety response with resistant starch and corn bran in human subjects. Nutr. Res. 2009, 29, 100–105. [Google Scholar] [CrossRef]
- Sgrulletta, D.; Scalfati, G.; De Stefanis, E.; Conciatori, A. Dietary fibre components of high-fibre commercial products. Effect of cooking process. Ital. J. Food Sci. 2005, 17, 285–294. [Google Scholar]
- Hsu, R.J.-C.; Chen, H.-J.; Lu, S.; Chiang, W. Effects of cooking, retrogradation and drying on starch digestibility in instant rice making. J. Cereal Sci. 2015, 65, 154–161. [Google Scholar] [CrossRef]
- Patindol, J.; Gu, X.; Wang, Y.-J. Chemometric analysis of cooked rice texture in relation to starch fine structure and leaching characteristics. Starch 2010, 62, 188–197. [Google Scholar] [CrossRef]
- Hu, P.; Zhao, H.; Duan, Z.; Linlin, Z.; Wu, D. Starch digestibility and the estimated glycemic score of different types of rice differing in amylose contents. J. Cereal Sci. 2004, 40, 231–237. [Google Scholar] [CrossRef]
- Wei, C.; Qin, F.; Zhu, L.; Zhou, W.; Chen, Y.; Wang, Y.; Gu, M.; Liu, Q. Microstructure and Ultrastructure of High-Amylose Rice Resistant Starch Granules Modified by Antisense RNA Inhibition of Starch Branching Enzyme. J. Agric. Food Chem. 2010, 58, 1224–1232. [Google Scholar] [CrossRef]
- Chung, H.-J.; Liu, Q.; Lee, L.; Wei, D. Relationship between the structure, physicochemical properties and in vitro digestibility of rice starches with different amylose contents. Food Hydrocoll. 2011, 25, 968–975. [Google Scholar] [CrossRef]
- Bao, J.; Xu, F.; Park, Y.; Zhou, X.; He, Q. Genome-wide association study of the resistant starch content in rice grains. Starch 2017, 69, 1600343. [Google Scholar] [CrossRef]
- Kong, X.; Chen, Y.; Zhu, P.; Sui, Z.; Corke, H.; Bao, J. Relationships among Genetic, Structural, and Functional Properties of Rice Starch. J. Agric. Food Chem. 2015, 63, 6241–6248. [Google Scholar] [CrossRef] [PubMed]
- Butardo, V.M.; Fitzgerald, M.A.; Bird, A.R.; Gidley, M.J.; Flanagan, B.M.; Larroque, O.; Resurreccion, A.P.; Laidlaw, H.K.C.; Jobling, S.A.; Morell, M.K.; et al. Impact of down-regulation of starch branching enzyme IIb in rice by artificial microRNA- and hairpin RNA-mediated RNA silencing. J. Exp. Bot. 2011, 62, 4927–4941. [Google Scholar] [CrossRef] [Green Version]
- Daygon, V.D.; Colgrave, M.L.; Campbell, P.M.; Resurreccion, A.; Cuevas, R.P.; Jobling, S.A.; Tetlow, I.; Rahman, S.; Morell, M.; Fitzgerald, M.; et al. Biomolecular Analyses of Starch and Starch Granule Proteins in the High-Amylose Rice Mutant Goami 2. J. Agric. Food Chem. 2012, 60, 11576–11585. [Google Scholar]
- Regina, A.; Bird, A.; Topping, D.; Bowden, S.; Freeman, J.; Barsby, T.; Kosar-Hashemi, B.; Li, Z.; Rahman, S.; Morell, M. High-amylose wheat generated by RNA interference improves indices of large-bowel health in rats. Proc. Natl. Acad. Sci. USA 2006, 103, 3546–3551. [Google Scholar] [CrossRef] [Green Version]
- Yang, R.; Sun, C.; Bai, J.; Luo, Z.; Shi, B.; Zhang, J.; Yan, W.; Piao, Z. A Putative Gene sbe3-rs for Resistant Starch Mutated from SBE3 for Starch Branching Enzyme in Rice (Oryza sativa L.). PLoS ONE 2012, 7, e43026. [Google Scholar] [CrossRef]
- Shu, X.; Jia, L.; Ye, H.; Li, C.; Wu, D. Slow Digestion Properties of Rice Different in Resistant Starch. J. Agric. Food Chem. 2009, 57, 7552–7559. [Google Scholar] [CrossRef]
- Rahman, S.; Bird, A.; Regina, A.; Li, Z.; Ral, J.P.; McMaugh, S.; Topping, D.; Morell, M. Resistant starch in cereals: Exploiting genetic engineering and genetic variation. J. Cereal Sci. 2007, 46, 251–260. [Google Scholar] [CrossRef]
- Zhu, L.J.; Gu, M.H.; Meng, X.L.; Cheung, S.C.K.; Yu, H.X.; Huang, J.; Sun, Y.; Shi, Y.; Liu, Q. High-amylose rice improves indices of animal health in normal and diabetic rats. Plant Biotechnol. J. 2012, 10, 353–362. [Google Scholar] [CrossRef]
- Huang, J.; Shang, Z.; Man, J.; Liu, Q.; Zhu, C.; Wei, C. Comparison of molecular structures and functional properties of high-amylose starches from rice transgenic line and commercial maize. Food Hydrocoll. 2015, 46, 172–179. [Google Scholar] [CrossRef]
- Naguleswaran, S.; Vasanthan, T.; Hoover, R.; Bressler, D. Amylolysis of amylopectin and amylose isolated from wheat, triticale, corn and barley starches. Food Hydrocoll. 2014, 35, 686–693. [Google Scholar] [CrossRef]
- Shu, X.; Sun, J.; Wu, D. Effects of grain development on formation of resistant starch in rice. Food Chem. 2014, 164, 89–97. [Google Scholar] [CrossRef]
- Zhou, H.; Wang, L.; Liu, G.; Meng, X.; Jing, Y.; Shu, X.; Kong, X.; Sun, J.; Yu, H.; Smith, S.M.; et al. Critical roles of soluble starch synthase SSIIIa and granule-bound starch synthase Waxy in synthesizing resistant starch in rice. Proc. Natl. Acad. Sci. USA 2016, 113, 12844–12849. [Google Scholar] [CrossRef] [Green Version]
- Mou, F.G.; Yan, Z.W.; Ran, R.L.; Teng, J.X.; Chen, Y.B.; Yang, C.Z.; Li, M.H.; Wu, D.X. Preliminary studies on resistant starch-linked SSR marker in rice. Mol. Plant Breed. 2008, 6, 432–438. [Google Scholar]
- Zeng, Y.; Sun, D.; Du, J.; Pu, X.; Yang, S.; Yang, X.; Yang, T.; Yang, J. Identification of QTLs for resistant starch and total alkaloid content in brown and polished rice. Genet. Mol. Res. 2016, 15, 15037268. [Google Scholar] [CrossRef]
- Morell, M.K.; Kosar-Hashemi, B.; Cmiel, M.; Samuel, M.S.; Chandler, P.; Rahman, S.; Buleon, A.; Batey, I.L.; Li, Z. Barley sex6 mutants lack starch synthase IIa activity and contain a starch with novel properties. Plant J. 2003, 34, 173–185. [Google Scholar] [CrossRef]
- Kawahara, Y.; de la Bastide, M.; Hamilton, J.P.; Kanamori, H.; McCombie, W.R.; Ouyang, S.; Schwartz, D.C.; Tanaka, T.; Wu, J.; Zhou, S.; et al. Improvement of the Oryza sativa Nipponbare reference genome using next generation sequence and optical mapdata. Rice 2013, 6, 1–10. [Google Scholar] [CrossRef]
- Shannon, J.C. Brittle-1, an Adenylate Translocator, Facilitates Transfer of Extraplastidial Synthesized ADP-Glucose into Amyloplasts of Maize Endosperms. Plant Physiol. 1998, 117, 1235–1252. [Google Scholar] [CrossRef]
- Wang, X.; Pang, Y.; Zhang, J.; Wu, Z.; Chen, K.; Ali, J.; Ye, G.; Xu, J.; Li, Z. Genome-wide and gene-based association mapping for rice eating and cooking characteristics and protein content. Sci. Rep. 2017, 7, 17203. [Google Scholar] [CrossRef]
- Butardo, V.M.J.; Anacleto, R.; Parween, S.; Samson, I.; de Guzman, K.; Alhambra, C.M.; Misra, G.; Sreenivasulu, N. Systems Genetics Identifies a Novel Regulatory Domain of Amylose Synthesis. Plant Physiol. 2017, 173, 887–906. [Google Scholar] [CrossRef]
- Anacleto, R.; Badoni, S.; Parween, S.; Butardo, V.M.; Misra, G.; Cuevas, R.P.; Kuhlmann, M.; Trinidad, T.P.; Mallillin, A.C.; Acuin, C.; et al. Integrating a genome-wide association study with a large-scale transcriptome analysis to predict genetic regions influencing the glycaemic index and texture in rice. Plant Biotechnol. J. 2019, 17, 1261–1275. [Google Scholar] [CrossRef]
- Biscarini, F.; Cozzi, P.; Casella, L.; Riccardi, P.; Vattari, A.; Orasen, G.; Perrini, R.; Tacconi, G.; Tondelli, A.; Biselli, C.; et al. Genome-Wide Association Study for Traits Related to Plant and Grain Morphology, and Root Architecture in Temperate Rice Accessions. PLoS ONE 2016, 11, e0155425. [Google Scholar] [CrossRef]
- Volante, A.; Desiderio, F.; Tondelli, A.; Perrini, R.; Orasen, G.; Biselli, C.; Riccardi, P.; Vattari, A.; Cavalluzzo, D.; Urso, S.; et al. Genome-Wide Analysis of japonica Rice Performance under Limited Water and Permanent Flooding Conditions. Front. Plant Sci. 2017, 8, 1862. [Google Scholar] [CrossRef]
- Sinervo, B.; Svensson, E. Correlational selection and the evolution of genomic architecture. Heredity 2002, 89, 329–338. [Google Scholar] [CrossRef]
- Shu, X.; Backes, G.; Rasmussen, S.K. Genome-wide Association Study of Resistant Starch (RS) Phenotypes in a Barley Variety Collection. J. Agric. Food Chem. 2012, 60, 10302–10311. [Google Scholar] [CrossRef]
- GRiSP (Global Rice Science Partnership). Rice Almanac, 4th ed.; International Rice Research Institute: Los Baños, Philippines, 2013; p. 283. [Google Scholar]
- Mather, K.A.; Caicedo, A.L.; Polato, N.R.; Olsen, K.M.; McCouch, S.; Purugganan, M.D. The Extent of Linkage Disequilibrium in Rice (Oryza sativa L.). Genetics 2007, 177, 2223–2232. [Google Scholar] [CrossRef]
- Xu, X.; Liu, X.; Ge, S.; Jensen, J.D.; Hu, F.; Li, X.; Dong, Y.; Gutenkunst, R.N.; Fang, L.; Huang, L.; et al. Resequencing 50 accessions of cultivated and wild rice yields markers for identifying agronomically important genes. Nat. Biotechnol. 2011, 30, 105–111. [Google Scholar] [CrossRef]
- Kumar, V.; Singh, A.; Mithra, S.V.A.; Krishnamurthy, S.L.; Parida, S.K.; Jain, S.; Tiwari, K.K.; Kumar, P.; Rao, A.R.; Sharma, S.K.; et al. Genome-wide association mapping of salinity tolerance in rice (Oryza sativa). Curr. Neuropharmacol. 2015, 22, 133–145. [Google Scholar] [CrossRef]
- Morita, R.; Sugino, M.; Hatanaka, T.; Misoo, S.; Fukayama, H. CO2-Responsive CONSTANS, CONSTANS-Like, and Time of Chlorophyll a/b Binding Protein Expression1 Protein Is a Positive Regulator of Starch Synthesis in Vegetative Organs of Rice. Plant Physiol. 2015, 167, 1321–1331. [Google Scholar] [CrossRef]
- Sullivan, T.D.; Strelow, L.I.; Illingworth, C.A.; Phillips, R.L.; Nelson, O.E. Analysis of maize brittle-1 alleles and a defective Suppressor-mutator-induced mutable allele. Plant Cell 1991, 3, 1337–1348. [Google Scholar]
- Kossmann, J.; Lloyd, J. Understanding and Influencing Starch Biochemistry. Crit. Rev. Plant Sci. 2000, 19, 171–226. [Google Scholar] [CrossRef]
- Pu, C.-X.; Han, Y.-F.; Zhu, S.; Song, F.-Y.; Zhao, Y.; Wang, C.-Y.; Zhang, Y.-C.; Yang, Q.; Wang, J.; Bu, S.-L.; et al. The Rice Receptor-Like Kinases DWARF AND RUNTISH SPIKELET1 and 2 Repress Cell Death and Affect Sugar Utilization during Reproductive Development. Plant Cell 2017, 29, 70–89. [Google Scholar] [CrossRef] [Green Version]
- Kleczkowski, L.A. Glucose activation and metabolism through UDP-glucose pyrophosphorylase in plants. Phytochemistry 1994, 37, 1507–1515. [Google Scholar] [CrossRef]
- Volante, A.; Tondelli, A.; Aragona, M.; Valente, M.T.; Biselli, C.; Desiderio, F.; Bagnaresi, P.; Matic, S.; Gullino, M.L.; Infantino, A.; et al. Identification of bakanae disease resistance loci in japonica rice through genome wide association study. Rice 2017, 10, 29. [Google Scholar] [CrossRef]
- Perera, A.; Meda, V.; Tyler, R. Resistant starch: A review of analytical protocols for determining resistant starch and of factors affecting the resistant starch content of foods. Food Res. Int. 2010, 43, 1959–1974. [Google Scholar] [CrossRef]
- Ranghino, F. Valuation of rice resistance to cooking. Riso 1966, 15, 117–127. [Google Scholar]
- Purcell, S.; Neale, B.; Todd-Brown, K.; Thomas, L.; Ferreira, M.A.R.; Bender, D.; Maller, J.; Sklar, P.; De Bakker, P.I.W.; Daly, M.J.; et al. PLINK: A Tool Set for Whole-Genome Association and Population-Based Linkage Analyses. Am. J. Hum. Genet. 2007, 81, 559–575. [Google Scholar] [CrossRef] [Green Version]
- Bradbury, P.J.; Zhang, Z.; Kroon, D.E.; Casstevens, T.M.; Ramdoss, Y.; Buckler, E. TASSEL: Software for association mapping of complex traits in diverse samples. Bioinformatics 2007, 23, 2633–2635. [Google Scholar] [CrossRef]
- Kumar, S.; Stecher, G.; Tamura, K. MEGA7: Molecular evolutionary genetics analysis version 7.0. for bigger datasets. Mol. Biol. Evol. 2016, 33, 1870–1874. [Google Scholar] [CrossRef]
- Mangin, B.; Siberchicot, A.; Nicolas, S.; Doligez, A.; This, P.; Cierco-Ayrolles, C. Novel measures of linkage disequilibrium that correct the bias due to population structure and relatedness. Heredity 2012, 108, 285–291. [Google Scholar] [CrossRef]
- Cleveland, W.S. Robust Locally Weighted Regression and Smoothing Scatterplots. J. Am. Stat. Assoc. 1979, 74, 829–836. [Google Scholar] [CrossRef]
- Marroni, F.; Pinosio, S.; Zaina, G.; Fogolari, F.; Felice, N.; Cattonaro, F.; Morgante, M. Nucleotide diversity and linkage disequilibrium in Populus nigra cinnamyl alcohol dehydrogenase (CAD4) gene. Tree Genet. Genomes 2011, 7, 1011–1023. [Google Scholar] [CrossRef]
- Barrett, J.C.; Fry, B.; Maller, J.; Daly, M.J. Haploview: Analysis and visualization of LD and haplotype maps. Bioinformatics 2005, 21, 263–265. [Google Scholar] [CrossRef]
- Benjamini, Y.; Hochberg, Y. Controlling the False Discovery Rate: A Practical and Powerful Approach to Multiple Testing. J. R. Stat. Soc. Ser. B 1995, 57, 289–300. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Trait | MTA | Peak Marker/Region (bp) | Chr | -Log10(p) | SNPs | Associated Region (bp) | Peak Marker R2 | LD Block (bp) | ||
|---|---|---|---|---|---|---|---|---|---|---|
| Start | End | Size | ||||||||
| RS | RS_MTA1 | S1_32380039 | 1 | 9.15 | 167 | 31,035,257 | 35,233,919 | 4,198,662 | 0.29 | 30,645,440–33,485,305 |
| RS_MTA2 | S2_9401106 | 2 | 3.75 | 1 | 9,401,106 | - | - | 0.12 | - | |
| RS_MTA3 | S3_15491784 | 3 | 3.60 | 1 | 15,491,784 | - | - | 0.11 | - | |
| RS_MTA5_1 | S5_3623956-S5_3623970 | 5 | 3.59 | 2 | 3,623,956 | 3,623,970 | 14 | 0.11 | - | |
| RS_MTA5_2 | S5_14198504 | 5 | 3.81 | 14 | 13,665,383 | 14,209,223 | 543,840 | 0.12 | 13,280,433–14,210,256 | |
| RS_MTA5_3 | S5_29890589 | 5 | 4.24 | 2 | 29,682,002 | 29,890,589 | 208,587 | 0.13 | 27,439,935–29,890,589 | |
| RS_MTA6_1 | S6_4418603 | 6 | 9.10 | 121 | 4,160,208 | 6,202,214 | 2,042,006 | 0.29 | 4,160,208–6,346,972 | |
| RS_MTA6_2 | S6_24395396 | 6 | 3.74 | 1 | 24,395,396 | - | - | 0.12 | 23,667,524–25,563,640 | |
| RS_MTA6_3 | S6_28384300-S6_28384505 | 6 | 4.08 | 2 | 28,384,300 | 28,384,505 | 205 | 0.13 | - | |
| RS_MTA8 | S8_8352009-S8_8367922 | 8 | 4.10 | 2 | 8,352,009 | 8,367,922 | 15,913 | 0.13 | 8,313,616–8,332,478; 8,422,453–8,441,645 | |
| RS_MTA11 | S11_18059435 | 11 | 7.73 | 13 | 17,462,389 | 18,145,389 | 683,000 | 0.25 | 17,418,693–18,592,536 | |
| AAC | AAC_MTA6_1 | TBGI270314 | 6 | 5.30 | 1 | 1,766,846 | - | - | 0.17 | - |
| AAC_MTA6_2 | TBGI270316 | 6 | 5.99 | 1 | 1,768,006 | - | - | 0.19 | - | |
| AAC_MTA6_3 | S6_4425552 | 6 | 4.35 | 76 | 4,193,908 | 4,445,114 | 251,206 | 0.14 | 4,160,208–4,930,358 | |
| AAC_MTA6_4 | S6_6086803-S6_6202214 | 6 | 3.80 | 5 | 6,086,803 | 6,202,214 | 115,411 | 0.12 | 5,593,250–6,222,678 | |
| AAC_MTA6_5 | S6_25964504-S6_25968586 | 6 | 5.10 | 9 | 25,963,067 | 26,023,769 | 60,702 | 0.16 | 25,963,067–27,324,453 | |
| Trait | Gene | Gene Description | Chr | RAP ID | Position (bp) | MTAs | |
|---|---|---|---|---|---|---|---|
| Start | End | ||||||
| RS | DRUS2 | Dwarf and Runtish Spikelet 2 | 1 | Os01g0769700 | 32,461,878 | 32,465,053 | RS_MTA1 |
| RS | AGPL3 | ADPGlucose Pyrophosphorylase Large subunit 3 | 5 | Os05g0580000 | 28,871,794 | 28,877,277 | RS_MTA5_3 |
| RS | CRCT | CO2-Responsive CONSTANS, CONSTANS-like, and Time of Chlorophyll a/b Binding Protein1 (CCT) | 5 | Os05g0595300 | 29,656,681 | 29,660,315 | RS_MTA5_3 |
| AAC | Wx1, GBSSI | Waxy. Granule Bound Starch Synthase I | 6 | Os06g0133000 | 1,765,622 | 1,770,653 | AAC_MTA6_1, AAC_MTA6_2 |
| RS | BT1-3 | Brittle1-3 | 6 | Os06g0602700 | 23,823,461 | 23,827,296 | RS_MTA6_2 |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Biselli, C.; Volante, A.; Desiderio, F.; Tondelli, A.; Gianinetti, A.; Finocchiaro, F.; Taddei, F.; Gazza, L.; Sgrulletta, D.; Cattivelli, L.; et al. GWAS for Starch-Related Parameters in Japonica Rice (Oryza sativa L.). Plants 2019, 8, 292. https://0-doi-org.brum.beds.ac.uk/10.3390/plants8080292
Biselli C, Volante A, Desiderio F, Tondelli A, Gianinetti A, Finocchiaro F, Taddei F, Gazza L, Sgrulletta D, Cattivelli L, et al. GWAS for Starch-Related Parameters in Japonica Rice (Oryza sativa L.). Plants. 2019; 8(8):292. https://0-doi-org.brum.beds.ac.uk/10.3390/plants8080292
Chicago/Turabian StyleBiselli, Chiara, Andrea Volante, Francesca Desiderio, Alessandro Tondelli, Alberto Gianinetti, Franca Finocchiaro, Federica Taddei, Laura Gazza, Daniela Sgrulletta, Luigi Cattivelli, and et al. 2019. "GWAS for Starch-Related Parameters in Japonica Rice (Oryza sativa L.)" Plants 8, no. 8: 292. https://0-doi-org.brum.beds.ac.uk/10.3390/plants8080292