Ecotoxicological Effects of Ibuprofen on Plant Growth of Vigna unguiculata L.

 ,
, 
Abstract
:1. Introduction
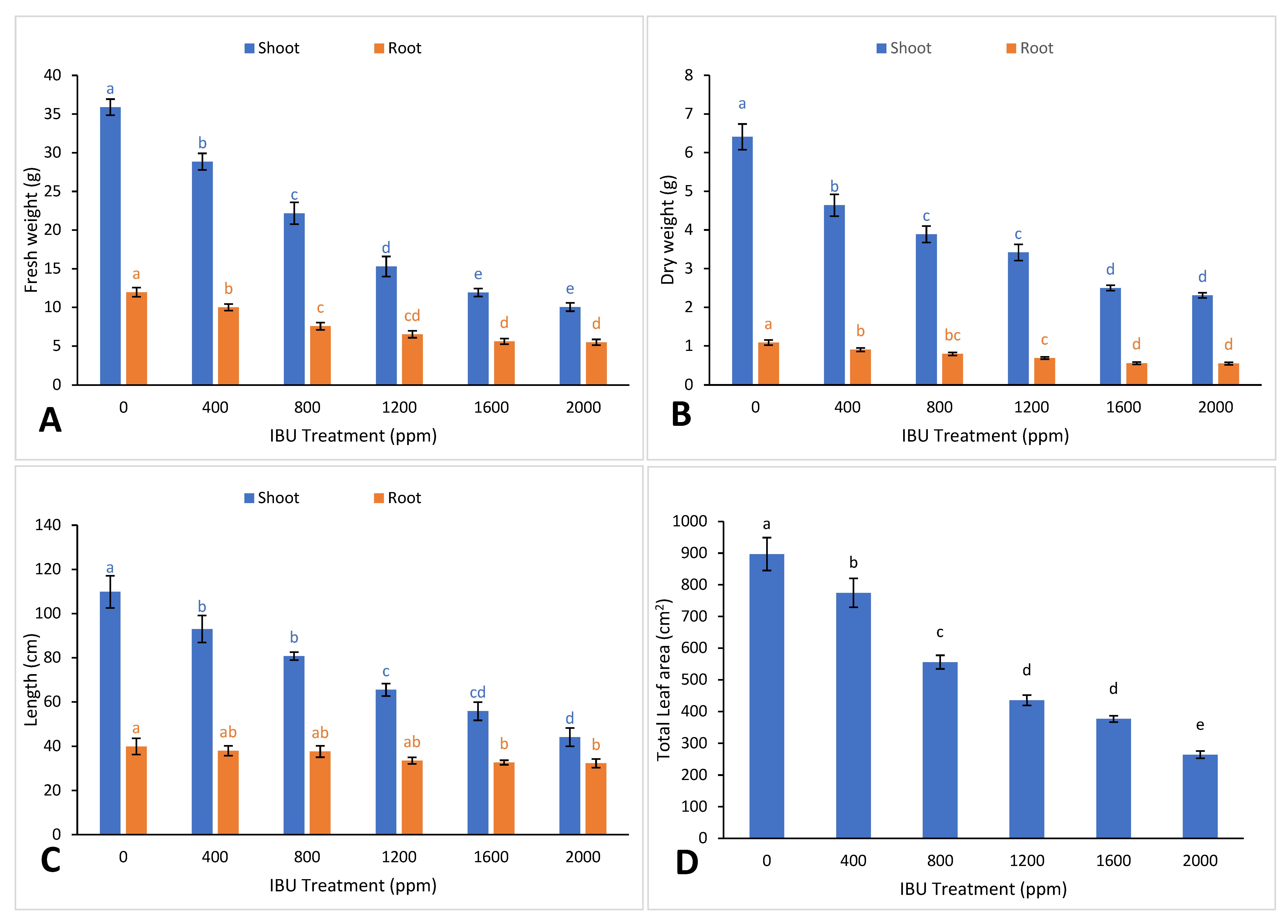
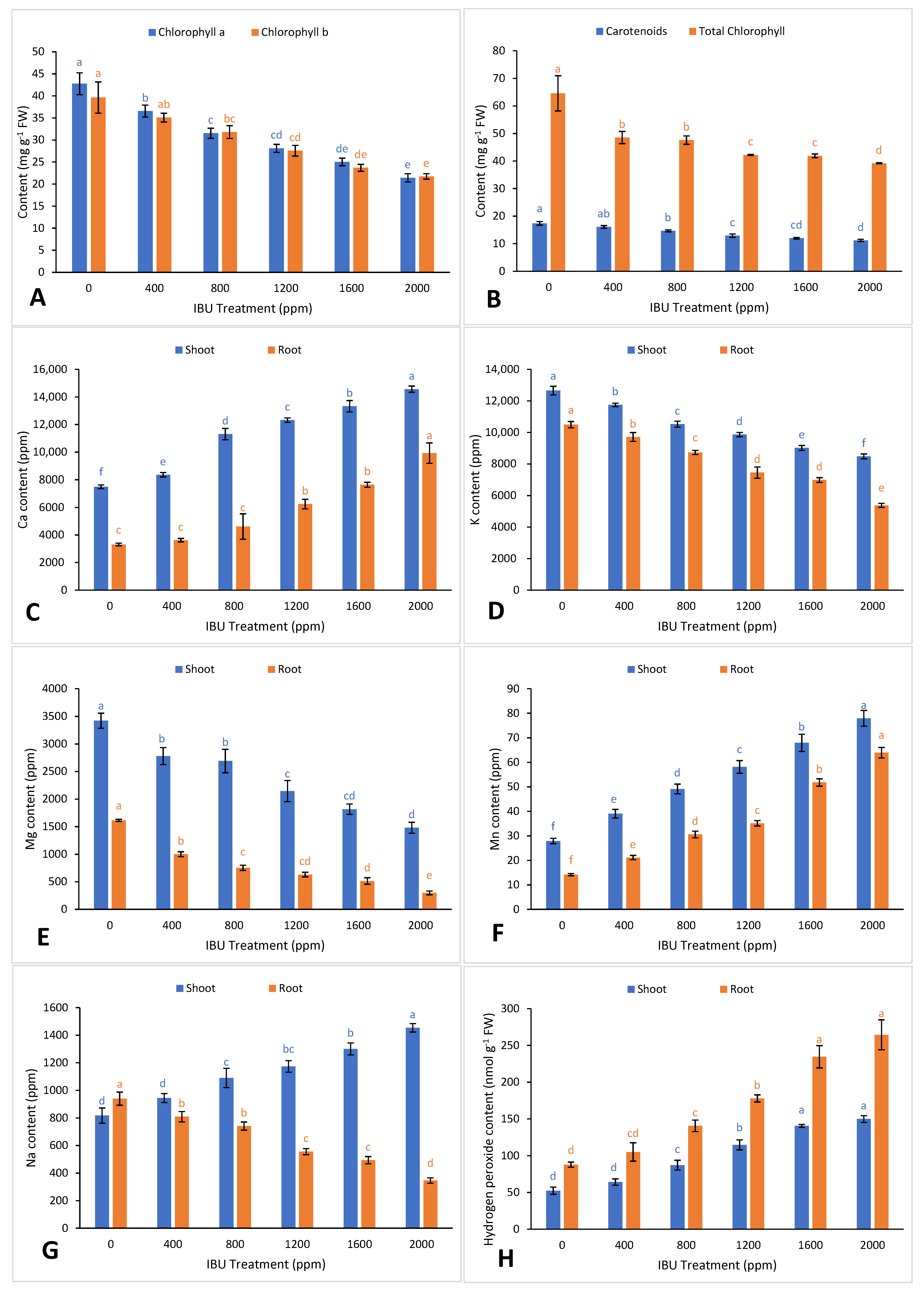
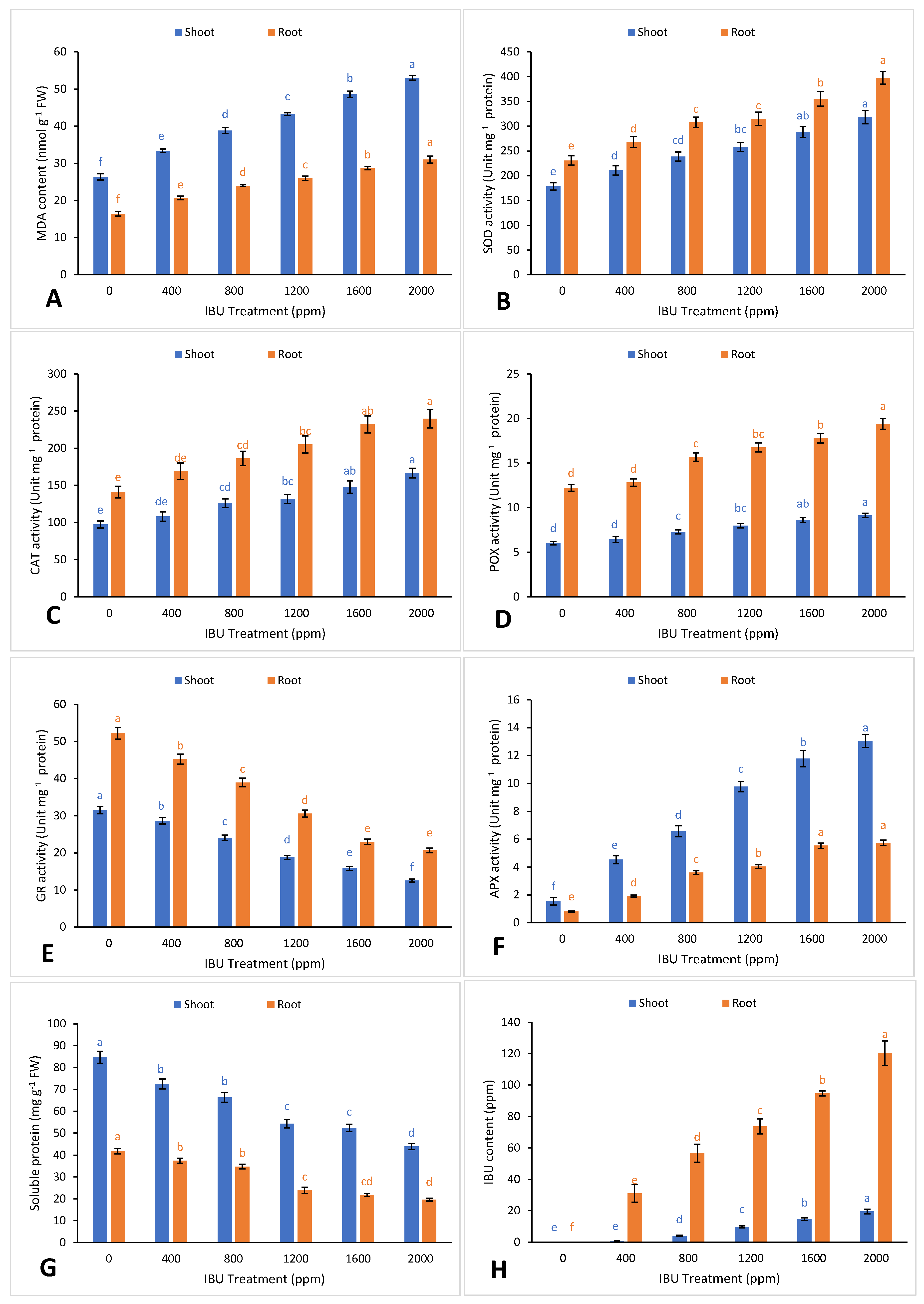
2. Results and Discussion
3. Materials and Methods
3.1. Plant Growth Experiment
3.2. Physicochemical Analysis
3.2.1. Chlorophyll and Carotenoid
3.2.2. Ca, K, Mg, Mn, and Na Content
3.2.3. Hydrogen Peroxide Content
3.2.4. Malondialdehyde (MDA) Assay
3.2.5. Enzyme Activity and Soluble Protein
3.2.6. Ibuprofen Analysis
3.3. Statistical Analysis
4. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Torchilin, V.P. Drug targeting. Eur. J. Pharm. Sci. 2000, 11, S81–S91. [Google Scholar] [CrossRef]
- Ellis, J.B. Pharmaceutical and personal care products (PPCPs) in urban receiving waters. Environ. Pollut. 2006, 144, 184–189. [Google Scholar] [CrossRef] [PubMed]
- Vatovec, C.; Van Wagoner, E.; Evans, C. Investigating sources of pharmaceutical pollution: Survey of over-the-counter and prescription medication purchasing, use, and disposal practices among university students. J. Environ. Manag. 2017, 198, 348–352. [Google Scholar] [CrossRef] [PubMed]
- Li, W.C. Occurrence, sources, and fate of pharmaceuticals in aquatic environment and soil. Environ. Pollut. 2014, 187, 193–201. [Google Scholar] [CrossRef]
- Tiwari, B.; Sellamuthu, B.; Ouarda, Y.; Drogui, P.; Tyagi, R.D.; Buelna, G. Review on fate and mechanism of removal of pharmaceutical pollutants from wastewater using biological approach. Bioresour. Technol. 2017, 224, 1–12. [Google Scholar] [CrossRef] [Green Version]
- Shaaban, H. Pharmaceutical pollution: Perception and management. IJCST 2017, 105. [Google Scholar] [CrossRef]
- Beroe-Inc. Ibuprofen Market Intelligence; Beroe Inc.: Raleigh, NC, USA, 2019. [Google Scholar]
- González-Naranjo, V.; Boltes, K.; Biel, M. Mobility of ibuprofen, a persistent active drug, in soils irrigated with reclaimed water. Plant Soil Environ. 2013, 59, 68–73. [Google Scholar] [CrossRef] [Green Version]
- Holling, C.S.; Bailey, J.L.; Heuvel, B.V.; Kinney, C.A. Uptake of human pharmaceuticals and personal care products by cabbage (Brassica campestris) from fortified and biosolids-amended soils. J. Environ. Monit. 2012, 14, 3029–3036. [Google Scholar] [CrossRef]
- Moro, I.; Matozzo, V.; Piovan, A.; Moschin, E.; Vecchia, F. Morpho-physiological effects of ibuprofen on Scenedesmus rubescens. Environ. Toxicol. Pharmacol. 2014, 38, 379–387. [Google Scholar] [CrossRef]
- Pomati, F.; Netting, A.G.; Calamari, D.; Neilan, B.A. Effects of erythromycin, tetracycline and ibuprofen on the growth of Synechocystis sp. and Lemna minor. Aquatic Toxicol. 2004, 67, 387–396. [Google Scholar] [CrossRef]
- Maamar, M.B.; Lesné, L.; Hennig, K.; Desdoits-Lethimonier, C.; Kilcoyne, K.R.; Coiffec, I.; Rolland, A.D.; Chevrier, C.; Kristensen, D.M.; Lavoué, V. Ibuprofen results in alterations of human fetal testis development. Sci. Rep. 2017, 7, 44184. [Google Scholar] [CrossRef]
- Xia, L.; Zheng, L.; Zhou, J.L. Effects of ibuprofen, diclofenac and paracetamol on hatch and motor behavior in developing zebrafish (Danio rerio). Chemosphere 2017, 182, 416–425. [Google Scholar] [CrossRef]
- Březinova, T.D.; Vymazal, J.; Koželuh, M.; Kule, L. Occurrence and removal of ibuprofen and its metabolites in full-scale constructed wetlands treating municipal wastewater. Ecol. Eng. 2018, 120, 1–5. [Google Scholar] [CrossRef]
- Picó, Y.; Alvarez-Ruiz, R.; Alfarhan, A.H.; El-Sheikh, M.A.; Alshahrani, H.O.; Barceló, D. Pharmaceuticals, pesticides, personal care products and microplastics contamination assessment of Al-Hassa irrigation network (Saudi Arabia) and its shallow lakes. Sci. Total Environ. 2020, 701, 135021. [Google Scholar] [CrossRef] [PubMed]
- Alshahrani, H. Evaluation of Ecological Pollution by Pesticides, Pharmaceuticals and Microplastics in Al-Hassa Lakes, Saudi Arabia; King Saud University: Riyadh, Saudi Arabia, 2020. [Google Scholar]
- Ehlers, J.D.; Hall, A.E. Cowpea (Vigna unguiculata L. Walp.). Field Crops Res. 1997, 53, 187–204. [Google Scholar] [CrossRef]
- Timko, M.P.; Ehlers, J.D.; Roberts, P.A. Cowpea. In Pulses, Sugar and Tuber Crops; Springer: Berlin/Heidelberg, Germany, 2007; pp. 49–67. [Google Scholar]
- Atkinson, N.J.; Urwin, P.E. The interaction of plant biotic and abiotic stresses: From genes to the field. J. Exp. Bot. 2012, 63, 3523–3543. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kumar, V.L.; Singhal, A. Germinating seeds of the mung bean, Vigna radiata (Fabaceae), as a model for the preliminary evaluation of cytotoxic effects of drugs. Biocell 2009, 33, 19–24. [Google Scholar] [CrossRef]
- Millaleo, R.; Reyes-Díaz, M.; Ivanov, A.; Mora, M.; Alberdi, M. Manganese as essential and toxic element for plants: Transport, accumulation and resistance mechanisms. J. Soil Sci. Plant Nutr. 2010, 10, 470–481. [Google Scholar] [CrossRef] [Green Version]
- Iori, V.; Zacchini, M.; Pietrini, F. Growth, physiological response and phytoremoval capability of two willow clones exposed to ibuprofen under hydroponic culture. J. Hazard. Mater. 2013, 262, 796–804. [Google Scholar] [CrossRef]
- Zhang, L.; Lv, T.; Zhang, Y.; Stein, O.R.; Arias, C.A.; Brix, H.; Carvalho, P.N. Effects of constructed wetland design on ibuprofen removal—A mesocosm scale study. Sci. Total Environ. 2017, 609, 38–45. [Google Scholar] [CrossRef]
- Kotyza, J.; Soudek, P.; Kafka, Z.; Vaněk, T. Phytoremediation of Pharmaceuticals—Preliminary Study. Int. J. Phytoremediat. 2010, 12, 306–316. [Google Scholar] [CrossRef]
- Mohamed, H.I.; Akladious, S.A. Changes in antioxidants potential, secondary metabolites and plant hormones induced by different fungicides treatment in cotton plants. Pestic. Biochem. Physiol. 2017, 142, 117–122. [Google Scholar] [CrossRef]
- Picó, Y.; Alvarez-Ruiz, R.; Wijaya, L.; Alfarhan, A.; Alyemeni, M.; Barceló, D. Analysis of ibuprofen and its main metabolites in roots, shoots, and seeds of cowpea (Vigna unguiculata L. Walp) using liquid chromatography-quadrupole time-of-flight mass spectrometry: Uptake, metabolism, and translocation. Anal. Bioanal. Chem. 2018, 410, 1163–1176. [Google Scholar] [CrossRef] [PubMed]
- Wrede, J. Pharmaceutically Active Compounds Might Bioaccumulate and Cause Effects on the Free–Floating Macrophyte; Humboldt-Universität zu Berlin: Berlin, Germany, 2015. [Google Scholar]
- Liu, Y.-F.; Han, X.-R.; Zhan, X.-M.; Yang, J.-F.; Wang, Y.-Z.; Song, Q.-B.; Chen, X. Regulation of calcium on peanut photosynthesis under low night temperature stress. J. Integr. Agric. 2013, 12, 2172–2178. [Google Scholar] [CrossRef]
- Maathuis, F.J.; Diatloff, E. Roles and functions of plant mineral nutrients. In Plant Mineral Nutrients; Springer: Berlin/Heidelberg, Germany, 2013; pp. 1–21. [Google Scholar]
- Ramalho, J.C.; Rebelo, M.; Santos, M.E.; Antunes, M.L.; Nunes, M.A. Effects of calcium deficiency on Coffea arabica. Nutrient changes and correlation of calcium levels with some photosynthetic parameters. Plant Soil 1995, 172, 87–96. [Google Scholar] [CrossRef]
- Hochmal, A.K.; Schulze, S.; Trompelt, K.; Hippler, M. Calcium-dependent regulation of photosynthesis. Biochim. Biophys. Acta (BBA) Bioenerg. 2015, 1847, 993–1003. [Google Scholar] [CrossRef] [Green Version]
- Mahouachi, J.; Socorro, A.R.; Talon, M. Responses of papaya seedlings (Carica papaya L.) to water stress and re-hydration: Growth, photosynthesis and mineral nutrient imbalance. Plant Soil 2006, 281, 137–146. [Google Scholar] [CrossRef]
- Zhao, H.; Zhou, Q.; Zhou, M.; Li, C.; Gong, X.; Liu, C.; Qu, C.; Wang, L.; Si, W.; Hong, F. Magnesium deficiency results in damage of nitrogen and carbon cross-talk of maize and improvement by cerium addition. Biol. Trace Elem. Res. 2012, 148, 102–109. [Google Scholar] [CrossRef]
- Liu, M.; Liu, X.X.; He, X.L.; Liu, L.J.; Wu, H.; Tang, C.X.; Zhang, Y.S.; Jin, C.W. Ethylene and nitric oxide interact to regulate the magnesium deficiency-induced root hair development in Arabidopsis. New Phytol. 2017, 213, 1242–1256. [Google Scholar] [CrossRef]
- Munns, R. Comparative physiology of salt and water stress. Plant Cell Environ. 2002, 25, 239–250. [Google Scholar] [CrossRef]
- Ahmad, P.; Sarwat, M.; Sharma, S. Reactive oxygen species, antioxidants and signaling in plants. J. Plant Biol. 2007, 51, 167–173. [Google Scholar] [CrossRef]
- Del Rio, D.; Stewart, A.J.; Pellegrini, N. A review of recent studies on malondialdehyde as toxic molecule and biological marker of oxidative stress. Nutr. Metab. Cardiovasc. Dis. 2005, 15, 316–328. [Google Scholar] [CrossRef] [PubMed]
- Almohisen, I.A. Effect of paracetamol on antioxidant system and osmoprotectants of mung bean (Vigna radiata). Appl. Ecol. Environ. Res. 2019, 17, 4297–4303. [Google Scholar] [CrossRef]
- Kummerová, M.; Zezulka, Š.; Babula, P.; Tříska, J. Possible ecological risk of two pharmaceuticals diclofenac and paracetamol demonstrated on a model plant Lemna minor. J. Hazard. Mater. 2016, 302, 351–361. [Google Scholar] [CrossRef]
- Ahmad, P.; Jaleel, C.A.; Salem, M.A.; Nabi, G.; Sharma, S. Roles of enzymatic and nonenzymatic antioxidants in plants during abiotic stress. Crit. Rev. Biotechnol. 2010, 30, 161–175. [Google Scholar] [CrossRef] [PubMed]
- Bowler, C.; Van Camp, W.; Van Montagu, M.; Inze, D.; Asada, K. Superoxide dismutase in plants. Crit. Rev. Plant Sci. 1994, 13, 199–218. [Google Scholar] [CrossRef]
- Carvalho, L.C.; Vidigal, P.; Amancio, S. Oxidative stress homeostasis in grapevine (Vitis vinifera L.). Front. Environ. Sci. 2015, 3. [Google Scholar] [CrossRef] [Green Version]
- Bartha, B.; Huber, C.; Schröder, P. Uptake and metabolism of diclofenac in Typha latifolia—How plants cope with human pharmaceutical pollution. Plant Sci. 2014, 227, 12–20. [Google Scholar] [CrossRef]
- Pierattini, E.C.; Francini, A.; Huber, C.; Sebastiani, L.; Schröder, P. Poplar and diclofenac pollution: A focus on physiology, oxidative stress and uptake in plant organs. Sci. Total Environ. 2018, 636, 944–952. [Google Scholar] [CrossRef]
- Tripathi, A.; Gautam, M. Biochemical parameters of plants as indicators of air pollution. J. Environ. Biol. 2007, 28, 127. [Google Scholar]
- Hasan, M.K.; Cheng, Y.; Kanwar, M.K.; Chu, X.-Y.; Ahammed, G.J.; Qi, Z.-Y. Responses of Plant Proteins to Heavy Metal Stress—A Review. Front. Plant Sci. 2017, 8. [Google Scholar] [CrossRef] [Green Version]
- Di Baccio, D.; Pietrini, F.; Bertolotto, P.; Pérez, S.; Barcelò, D.; Zacchini, M.; Donati, E. Response of Lemna gibba L. to high and environmentally relevant concentrations of ibuprofen: Removal, metabolism and morpho-physiological traits for biomonitoring of emerging contaminants. Sci. Total Environ. 2017, 584–585, 363–373. [Google Scholar] [CrossRef]
- Calderón-Preciado, D.; Jiménez-Cartagena, C.; Matamoros, V.; Bayona, J. Screening of 47 organic microcontaminants in agricultural irrigation waters and their soil loading. Water Res. 2011, 45, 221–231. [Google Scholar] [CrossRef]
- Pietrini, F.; Di Baccio, D.; Aceña, J.; Pérez, S.; Barceló, D.; Zacchini, M. Ibuprofen exposure in Lemna gibba L.: Evaluation of growth and phytotoxic indicators, detection of ibuprofen and identification of its metabolites in plant and in the medium. J. Hazard. Mater. 2015, 300, 189–193. [Google Scholar] [CrossRef] [PubMed]
- He, Y.; Langenhoff, A.A.; Sutton, N.B.; Rijnaarts, H.H.; Blokland, M.H.; Chen, F.; Huber, C.; Schröder, P. Metabolism of ibuprofen by Phragmites australis: Uptake and phytodegradation. Environ. Sci. Technol. 2017, 51, 4576–4584. [Google Scholar] [CrossRef]
- Hawkins, T.S.; Gardiner, E.S.; Comer, G.S. Modeling the relationship between extractable chlorophyll and SPAD-502 readings for endangered plant species research. J. Nat. Conserv. 2009, 17, 123–127. [Google Scholar] [CrossRef]
- Lichtenthaler, H.K.; Welburn, A.R. Determination of total carotenoids and chlorophylls a and b of leaf extracts in different solvents. Biochem. Soc. Trans. 1983, 11, 591–592. [Google Scholar] [CrossRef] [Green Version]
- Porra, R.J. The chequered history of the development and use of simultaneous equations for the accurate determination of chlorophylls a and b. Photosynth. Res. 2002, 73, 149–156. [Google Scholar] [CrossRef]
- Price, W.J. Analytical Atomic Absorption Spectrometry; Heyden and Son: London, UK, 1974.
- Katz, S.A.; Jennis, S.W. Regulatory Compliance Monitoring by Atomic Absorption Spectroscopy; Verlag Chemie International: Tampa, FL, USA, 1983. [Google Scholar]
- Velikova, V.; Yordanov, I.; Edreva, A. Oxidative stress and some antioxidant systems in acid rain-treated bean plants. Plant Sci. 2000, 151, 59–66. [Google Scholar] [CrossRef]
- Heath, R.L.; Packer, L. Photoperoxidation in isolated chloroplasts: I. Kinetics and stoichiometry of fatty acid peroxidation. Arch. Biochem. Biophys. 1968, 125, 189–198. [Google Scholar] [CrossRef]
- Beauchamp, C.; Fridovich, I. Superoxide dismutase: Improved assays and an assay applicable to acrylamide gels. Anal. Biochem. 1971, 44, 276–287. [Google Scholar] [CrossRef]
- Aebi, H. Catalase in vitro. Methods Enzymol. 1984, 105, 121–126. [Google Scholar] [CrossRef]
- Tiedemann, A.V. Evidence for a primary role of active oxygen species in induction of host cell death during infection of bean leaves with Botrytis cinerea. Physiol. Mol. Plant Pathol. 1997, 50, 151–166. [Google Scholar] [CrossRef]
- Foster, J.G.; Hess, J.L. Responses of superoxide dismutase and glutathione reductase activities in cotton leaf tissue exposed to an atmosphere enriched in oxygen. Plant Physiol. 1980, 66, 482–487. [Google Scholar] [CrossRef] [PubMed]
- Nakano, Y.; Asada, K. Hydrogen peroxide is scavenged by ascorbate-specific peroxidase in spinach chloroplasts. Plant Cell Physiol. 1981, 22, 867–880. [Google Scholar]
- Bradford, M.M. A rapid and sensitive method for the quantitation of microgram quantities of protein utilizing the principle of protein-dye binding. Anal. Biochem. 1976, 72, 248–254. [Google Scholar] [CrossRef]
- Andreotti, F.; Mucha, A.P.; Caetano, C.; Rodrigues, P.; Gomes, C.R.; Almeida, C.M.R. Interactions between salt marsh plants and Cu nanoparticles–effects on metal uptake and phytoremediation processes. Ecotoxicol. Environ. Saf. 2015, 120, 303–309. [Google Scholar] [CrossRef]
- Wang, W.-X. Bioaccumulation and biomonitoring. In Marine Ecotoxicology; Elsevier: Amsterdam, The Netherlands, 2016; pp. 99–119. [Google Scholar]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Bioconcentration Factor (BCF) | Bioaccumulation Factor (BAF) | Translocation Factor (TF) | ||
|---|---|---|---|---|
| IBU Treatments (ppm) | Roots | Shoots | Whole Plants | From Roots to Shoots |
| 400 | 0.078 | 0.002 | 0.08 | 0.029 |
| 800 | 0.071 | 0.005 | 0.076 | 0.072 |
| 1200 | 0.061 | 0.008 | 0.07 | 0.132 |
| 1600 | 0.059 | 0.009 | 0.068 | 0.155 |
| 2000 | 0.06 | 0.01 | 0.07 | 0.162 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Wijaya, L.; Alyemeni, M.; Ahmad, P.; Alfarhan, A.; Barcelo, D.; El-Sheikh, M.A.; Pico, Y. Ecotoxicological Effects of Ibuprofen on Plant Growth of Vigna unguiculata L. Plants 2020, 9, 1473. https://0-doi-org.brum.beds.ac.uk/10.3390/plants9111473
Wijaya L, Alyemeni M, Ahmad P, Alfarhan A, Barcelo D, El-Sheikh MA, Pico Y. Ecotoxicological Effects of Ibuprofen on Plant Growth of Vigna unguiculata L. Plants. 2020; 9(11):1473. https://0-doi-org.brum.beds.ac.uk/10.3390/plants9111473
Chicago/Turabian StyleWijaya, Leonard, Mohammed Alyemeni, Parvaiz Ahmad, Ahmed Alfarhan, Damia Barcelo, Mohamed A. El-Sheikh, and Yolanda Pico. 2020. "Ecotoxicological Effects of Ibuprofen on Plant Growth of Vigna unguiculata L." Plants 9, no. 11: 1473. https://0-doi-org.brum.beds.ac.uk/10.3390/plants9111473