The Common Ice Plant (Mesembryanthemum crystallinum L.)–Phytoremediation Potential for Cadmium and Chromate-Contaminated Soils

 , ,
, ,
Abstract
:
1. Introduction
2. Results
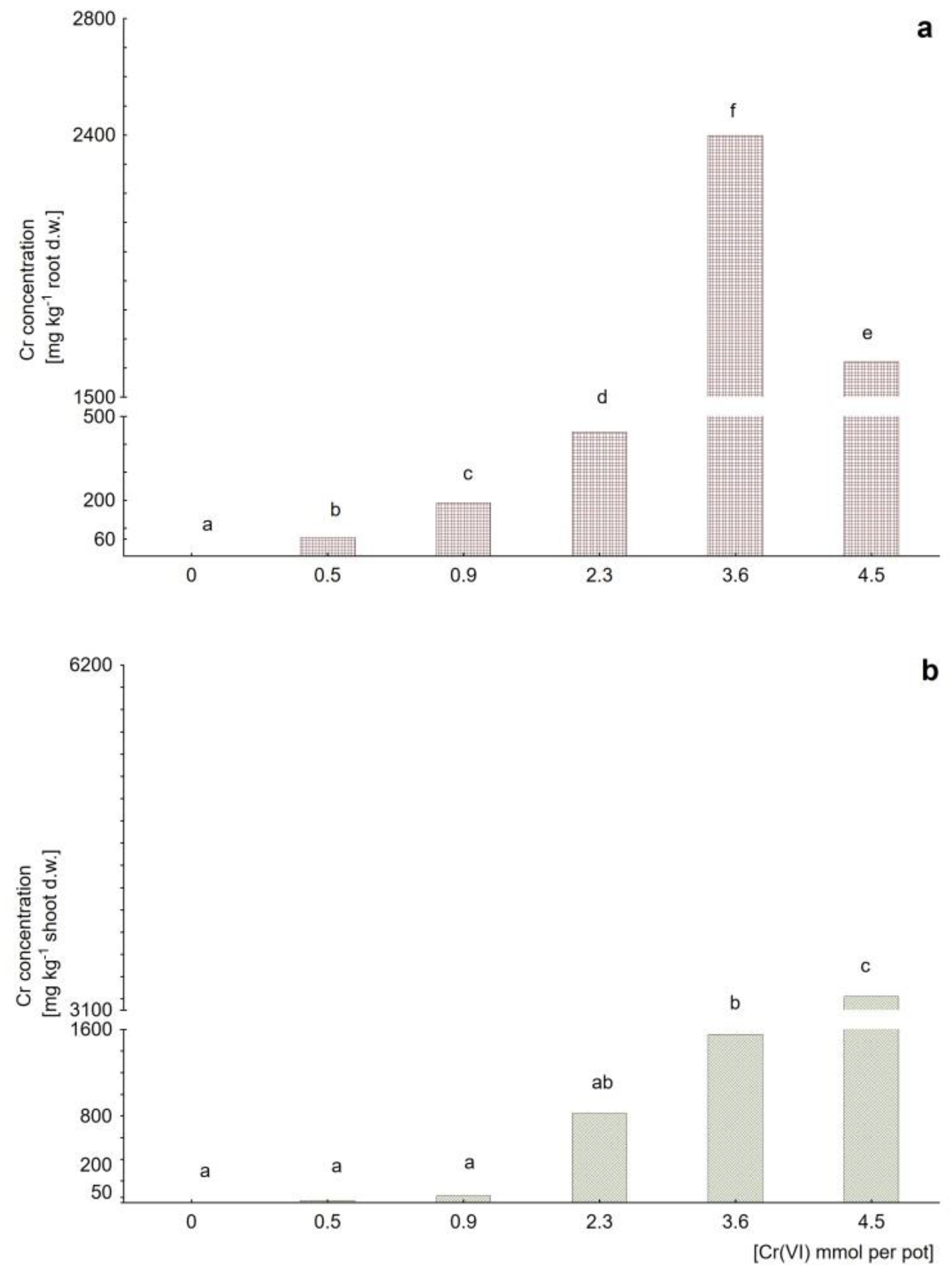
2.1. Evaluation of Cadmium and Chromate Toxicities
2.2. Cd and Cr Accumulation Capacities
3. Discussion
4. Materials and Methods
4.1. Plant Cultivation and Treatment with Heavy Metals
4.2. Biometric Analyses and Determination of Heavy Metals in Plant and Soil Samples
4.3. Evaluation of Heavy Metal Accumulation Capabilities: Bioaccumulation and Translocation Factors
4.4. Statistical Data Analysis
5. Concluding Remarks: Evaluating M. crystallinum Biotechnological Applicability
Author Contributions
Funding
Conflicts of Interest
References
- Karlen, D.L.; Rice, C.W. Soil degradation: Will humankind ever learn? Sustainability 2015, 7, 12490–12501. [Google Scholar] [CrossRef] [Green Version]
- Khan, A.; Khan, S.; Khan, M.A.; Qamar, Z.; Waqas, M. The uptake and bioaccumulation of heavy metals by food plants, their effects on plants nutrients, and associated health risk: A review. Environ. Sci. Pollut. Res. 2015, 22, 13772–13799. [Google Scholar] [CrossRef]
- Amari, T.; Ghnaya, T.; Debez, A.; Taamali, M.; Ben Youssef, N.; Lucchini, G.; Sacchi, G.A.; Abdelly, C. Comparative Ni tolerance and accumulation potentials between Mesembryanthemum crystallinum (halophyte) and Brassica juncea: Metal accumulation, nutrient status and photosynthetic activity. J. Plant Physiol. 2014, 171, 1634–1644. [Google Scholar] [CrossRef]
- Ashraf, M.Y.; Ashraf, M.; Mahmood, K.; Akhter, J.; Hussain, F.; Arshad, M. Phytoremediation of saline soils for sustainable agricultural productivity. In Plant Adaptation and Phytoremediation; Ashraf, M., Ozturk, M., Ahmad, M.S.A., Eds.; Springer Science+Business Media B.V.: Berlin/Heidelberg, Germany, 2010; pp. 335–356. [Google Scholar]
- Liang, L.; Liu, W.; Sun, Y.; Huo, X.; Li, S.; Zhou, Q. Phytoremediation of heavy metal contaminated saline soils using halophytes: Current progress and future perspectives. Environ. Rev. 2017, 25, 269–281. [Google Scholar] [CrossRef] [Green Version]
- Lutts, S.; Lefèvre, I. How can we take advantage of halophyte properties to cope with heavy metal toxicity in salt-affected areas? Ann. Bot. 2015, 115, 509–528. [Google Scholar] [CrossRef]
- Ali, H.; Khan, E.; Sajad, M.A. Phytoremediation of heavy metals—Concepts and applications. Chemosphere 2013, 91, 869–881. [Google Scholar] [CrossRef]
- EPA United States Environmental Protection Agency. Title 40 of the Code of Federal Regulations (CFR) Part 423, Appendix A. Priority Pollutant List, December 2014. Available online: https://www.epa.gov/emergency-response/40-code-federal-regulations-cfr-300900-920 (accessed on 15 July 2020).
- EU Directive. The European Parliament and the Council of the European Union 2008. Directive 2008/105/EC of 16 December 2008 on Environmental Quality Standards in the Field of Water Policy. Official Journal of the European Union, 24.12.2008, L348/84– L348/97. 2008. Available online: https://ec.europa.eu/environment/water/water-dangersub/pri_substances.htm (accessed on 15 July 2020).
- Ordinance of the Minister of the Environment of 1 September 2016 on the Methods of Evaluation of Surface Soil Contamination; Dz. U. (Journal of Laws); 5 September 2016, item 1395; Ministry of the Environment of the Republic of Poland: Warsaw, Poland, 2016.
- Andresen, E.; Küpper, H. Cadmium toxicity in plants. In Cadmium: From Toxicity to Essentiality. Metal Ions in Life Sciences; Sigel, A., Sigel, H., Sigel, R., Eds.; Springer: Dordrecht, The Netherlands, 2013; Volume 11, pp. 395–413. [Google Scholar] [CrossRef]
- He, S.; Yang, X.; He, Z.; Baligar, V.C. Morphological and physiological responses of plants to cadmium toxicity: A review. Pedosphere 2017, 27, 421–438. [Google Scholar] [CrossRef]
- Pavlaki, M.D.; Araújo, M.J.; Cardoso, D.N.; Silva, A.R.R.; Cruz, A.; Mendo, S.; Soares, A.M.V.M.; Calado, R.; Loureiro, S. Ecotoxicity and genotoxicity of cadmium in different marine trophic levels. Environ. Pollut. 2016, 215, 203–212. [Google Scholar] [CrossRef]
- Rizwan, M.; Ali, S.; Adrees, M.; Ibrahim, M.; Tsang, D.C.W.; Zia-ur-Rehman, M.; Zahir, A.Z.; Rinklebe, J.; Tack, F.M.G.; Ok, Y.S. A critical review on effects, tolerance mechanisms and management of cadmium in vegetables. Chemosphere 2017, 182, 90–105. [Google Scholar] [CrossRef]
- Shukla, U.C.; Murthy, R.C.; Kakkar, P. Combined effect of ultraviolet-B radiation and cadmium contamination on nutrient uptake and photosynthetic pigments in Brassica campestris L. seedlings. Environ. Toxicol. 2008, 23, 712–719. [Google Scholar] [CrossRef]
- Kabata-Pendias, A. Trace Elements in Soils and Plants, 4th ed.; CRC Press/Taylor & Francis Group: Boca Raton, FL, USA, 2010. [Google Scholar]
- Kotaś, J.; Stasicka, Z. Chromium occurrence in the environment and methods of its speciation. Environ. Pollut. 2000, 107, 263–283. [Google Scholar] [CrossRef]
- Zayed, A.M.; Terry, N. Chromium in the environment: Factors affecting biological remediation. Plant Soil 2003, 249, 139–156. [Google Scholar] [CrossRef]
- Oliveira, H. Chromium as an environmental pollutant: Insights on induced plant toxicity. J. Botany 2012, 8, 1–8. [Google Scholar] [CrossRef]
- Saha, R.; Nandi, R.; Saha, B. Sources and toxicity of hexavalent chromium. Review. J. Coord. Chem. 2011, 64, 1782–1806. [Google Scholar] [CrossRef]
- Shahid, M.; Shamshad, S.; Rafiq, M.; Khalid, S.; Bibi, I.; Niazi, N.K.; Dumat, C.; Rashid, M.I. Chromium speciation, bioavailability, uptake, toxicity and detoxification in soil-plant system: A review. Chemosphere 2017, 178, 513–533. [Google Scholar] [CrossRef]
- Zhitkovich, A. Chromium in drinking water: Sources, metabolism, and cancer risks. Chem. Res. Toxicol. 2011, 24, 1617–1629. [Google Scholar] [CrossRef]
- Prado, F.E.; Hila, M.; Chocobar-Ponce, S.; Pagano, E.; Rosa, M.; Prado, C. Chromium and the plant: A dangerous affair? In Plant Metal Interaction: Emerging Remediation Techniques; Ahmad, P., Ed.; Elsevier Inc: Amsterdam, The Netherlands, 2016; pp. 149–177. [Google Scholar]
- Singh, H.P.; Mahajan, P.; Kaur, S.; Batish, D.R.; Kohli, R.K. Chromium toxicity and tolerance in plants. Environ. Chem. Lett. 2013, 11, 229–254. [Google Scholar] [CrossRef]
- Shanker, A.K.; Cervantes, C.; Loza-Tavera, H.; Avudainayagam, S. Chromium toxicity in plants. Environ. Int. 2005, 31, 739–753. [Google Scholar] [CrossRef]
- Barceloux, D.G. Chromium. Clin. Toxicol. 1999, 37, 173–194. [Google Scholar] [CrossRef] [PubMed]
- Barnhart, J. Occurrences, uses, and properties of chromium. Regul. Toxicol. Pharmacol. 1997, 26, S3–S7. [Google Scholar] [CrossRef] [PubMed]
- Dominik, J.; Vignati, D.A.L.; Koukal, B.; Pereira de Abreu, M.H.; Kottelat, R.; Szalinska, E.; Baś, B.; Bobrowski, A. Speciation and environmental fate of chromium in rivers contaminated with tannery effluents. Eng. Life Sci. 2007, 7, 155–169. [Google Scholar] [CrossRef]
- Bartlett, R.J.; James, B. Behavior of chromium in soils: III. Oxidation. J. Environ. Qual. 1979, 8, 31–35. [Google Scholar] [CrossRef]
- Bartlett, R.J.; Kimble, J.M. Behavior of chromium in soils: II. Hexavalent forms. J. Environ. Qual. 1976, 5, 383–386. [Google Scholar] [CrossRef]
- Dominik, J.; Baś, B.; Bobrowski, A.; Dworak, T.; Koukal, B.; Niewiara, E.; Pereira de Abreu, M.H.; Rossé, P.; Szalinska, E.; Vignati, D. Partitioning of chromium (VI) and chromium (III) between dissolved and colloidal forms in a stream and reservoir contaminated with tannery waste water. J. de Physique IV 2003, 107, 385–388. [Google Scholar] [CrossRef]
- Ahmad, P. Plant Metal Interaction: Emerging Remediation Techniques; Elsevier Inc: Amsterdam, The Netherlands, 2016. [Google Scholar]
- Anjum, N.; Pereira, M.; Ahmad, I.; Duarte, A.; Umar, S.; Khan, N. Phytotechnologies: Remediation of Environmental Contaminants; CRC Press: Boca Raton, FL, USA, 2013. [Google Scholar]
- Robinson, B.; Green, S.; Mills, T.; Clothier, B.; Van Der Velde, M.; Laplane, R.; Fung, L.; Deurer, M.; Hurst, S.; Thayalakumaran, T.; et al. Phytoremediation: Using plants as biopumps to improve degraded environments. Aust. J. Soil Res. 2003, 41, 599–611. [Google Scholar] [CrossRef]
- Yang, X.; Feng, Y.; He, Z.L.; Stoffella, P.J. Molecular mechanisms of heavy metal hyperaccumulation and phytoremediation. J. Trace Elem. Med. Biol. 2005, 18, 339–353. [Google Scholar] [CrossRef]
- Favas, P.J.C.; Pratas, J.; Varun, M.; D’Souza, R.; Paul, M.S. Phytoremediation of soils contaminated with metals and metalloids at mining areas: Potential of native flora. In Environmental Risk Assessment of Soil Contamination; Hernandez-Soriano, M., Ed.; Intech Open: London, UK, 2014; pp. 486–516. [Google Scholar]
- Pulford, I.D.; Watson, C. Phytoremediation of heavy metal-contaminated land by trees—A review. Environ. Int. 2003, 29, 529–540. [Google Scholar] [CrossRef]
- Adams, P.; Nelson, D.E.; Yamada, S.; Chmara, W.; Jensen, R.G.; Bohnert, H.J.; Griffiths, H. Growth and development of Mesembryanthemum crystallinum (Aizoaceae). New Phytol. 1998, 138, 171–190. [Google Scholar] [CrossRef]
- Kornas, A.; Kuzniak, E.; Ślesak, I.; Miszalski, Z. The key role of the redox status in regulation of metabolism in photosynthesizing organisms. Acta Biochim. Pol. 2010, 57, 143–151. [Google Scholar] [CrossRef] [Green Version]
- Libik-Konieczny, M.; Kuźniak, E.; Surówka, E.; Ślesak, I.; Nosek, M.; Miszalski, Z. Crassulacean acid metabolism and its role in plant acclimatization to abiotic stresses and defence against pathogens. In Progress in Botany; Springer: Berlin/Heidelberg, Germany, 2019; pp. 1–30. [Google Scholar] [CrossRef]
- Kuznetsov, V.; Neto, D.S.; Borisova, N.N.; Dam, Z.B.; Rakitin, V.Y.; Aleksandrova, S.N.; Kholodova, V.P. Stress-induced CAM development and the limit of adaptation potential in Mesembryanthemum crystallinum plants under extreme conditions. Russ. J. Plant Physiol. 2000, 47, 168–175. [Google Scholar]
- Surówka, E.; Dziurka, M.; Kocurek, M.; Goraj, S.; Rapacz, M.; Miszalski, Z. Effects of exogenously applied hydrogen peroxide on antioxidant and osmoprotectant profiles and the C3-CAM shift in the halophyte Mesembryanthemum crystallinum L. J. Plant Physiol. 2016, 200, 102–110. [Google Scholar] [CrossRef] [PubMed]
- Cushman, J.C.; Michalowski, C.; Bohnert, H.J. Developmental control of Crassulacean acid metabolism inducibility by salt stress in the common ice plant. Plant Physiol. 1990, 94, 1137–1142. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kholodova, V.; Volkov, K.; Abdeyeva, A.; Kuznetsov, V. Water status in Mesembryanthemum crystallinum under heavy metal stress. Environ. Exp. Bot. 2011, 71, 382–389. [Google Scholar] [CrossRef]
- Miszalski, Z.; Ślesak, I.; Niewiadomska, E.; Bączek-Kwinta, R.; Lüttge, U.; Ratajczak, R. Subcellular localization and stress responses of superoxide dismutase isoforms from leaves in the C3→CAM intermediate halophyte Mesembryanthemum crystallinum L. Plant Cell Environ. 1998, 21, 169–179. [Google Scholar] [CrossRef]
- Shevyakova, N.I.; Netronina, I.A.; Aronova, E.E.; Kuznetsov, V.V. Compartmentation of cadmium and iron in Mesembryanthemum crystallinum plants during the adaptation to cadmium stress. Russ. J. Plant Physiol. 2003, 50, 678–685. [Google Scholar] [CrossRef]
- Kholodova, V.; Volkov, K.; Kuznetsov, V. Adaptation of the common ice plant to high copper and zinc concentrations and their potential using for phytoremediation. Russ. J. Plant Physiol. 2005, 52, 748–757. [Google Scholar] [CrossRef]
- Nosek, M.; Kaczmarczyk, A.; Śliwa, M.; Jędrzejczyk, R.; Kornaś, A.; Supel, P.; Kaszycki, P.; Miszalski, Z. The response of a model C3/CAM intermediate semi-halophyte Mesembryanthemum crystallinum L. to elevated cadmium concentrations. J. Plant Physiol. 2019, 240, 153005. [Google Scholar] [CrossRef]
- Nosek, M.; Kaczmarczyk, A.; Jędrzejczyk, R.J.; Supel, P.; Kaszycki, P.; Miszalski, Z. Expression of genes involved in heavy metal trafficking in plants exposed to salinity stress and elevated Cd concentrations. Plants 2020, 9, 475. [Google Scholar] [CrossRef] [Green Version]
- Manousaki, E.; Kalogerakis, N. Halophytes—An emerging trend in phytoremediation. Int. J. Phytoremed. 2011, 13, 959–969. [Google Scholar] [CrossRef]
- Tóth, G.; Hermann, T.; Da Silva, M.R.; Montanarella, L. Heavy metals in agricultural soils of the European Union with implications for food safety. Environ. Int. 2016, 88, 299–309. [Google Scholar] [CrossRef]
- Smolders, E.; Degryse, F. Fixation of cadmium and zinc in soils: Implications for risk assessment. In Natural Attenuation of Trace Element Availability in Soils; Hamon, R., McLaughlin, M., Lombi, E., Eds.; Taylor & Francis: Abingdon, UK, 2006; pp. 157–171. [Google Scholar]
- Hassan, W.; Bano, R.; Bashir, S.; Aslam, Z. Cadmium toxicity and soil biological index under potato (Solanum tuberosum L.) cultivation. Soil Res. 2016, 54, 460–468. [Google Scholar] [CrossRef]
- Ghnaya, T.; Nouairib, I.; Slamaa, I.; Messedia, D.; Grignonc, C.; Abdellya, C.; Ghorbel, M. Cadmium effects on growth and mineral nutrition of two halophytes: Sesuvium portulacastrum and Mesembryanthemum crystallinum. J. Plant Physiol. 2005, 162, 1133–1140. [Google Scholar] [CrossRef] [PubMed]
- Kaszycki, P.; Gabryś, H.; Appenroth, K.J.; Jaglarz, A.; Sędziwy, S.; Walczak, T.; Koloczek, H. Exogenously applied sulfate as a tool to investigate transport and reduction of chromate in the duckweed Spirodela polyrhiza. Plant Cell Environ. 2005, 28, 260–268. [Google Scholar] [CrossRef]
- Kaszycki, P.; Dubicka-Lisowska, A.; Augustynowicz, J.; Piwowarczyk, B.; Wesołowski, W. Callitriche cophocarpa (water starwort) proteome under chromate stress: Evidence for induction of a quinone reductase. Environ. Sci. Pollut. Res. 2018, 25, 8928–8942. [Google Scholar] [CrossRef] [Green Version]
- Niewiadomska, E.; Miszalski, Z. Partial characterization and expression of leaf catalase in the CAM-inducible halophyte Mesembryanthemum crystallinum L. Plant Physiol. Biochem. 2008, 46, 421–427. [Google Scholar] [CrossRef]
- Niewiadomska, E.; Bilger, W.; Gruca, M.; Mulisch, M.; Miszalski, Z.; Krupinska, K. CAM-related changes in chloroplastic metabolism of Mesembryanthemum crystallinum L. Planta 2011, 233, 275–285. [Google Scholar] [CrossRef] [Green Version]
- Van der Ent, A.; Baker, A.J.M.; Reeves, R.D.; Pollard, A.J.; Schat, H. Hyperaccumulators of metal and metalloid trace elements: Facts and fiction. Plant Soil 2013, 362, 319–334. [Google Scholar] [CrossRef]
- Shahandeh, H.; Hossner, L.R. Plant screening for chromium phytoremediation. Int. J. Phytoremed. 2000, 2, 31–51. [Google Scholar] [CrossRef]
- Ślesak, I.; Miszalski, Z.; Karpinska, B.; Niewiadomska, E.; Ratajczak, R.; Karpinski, S. Redox control of oxidative stress responses in the C3-CAM intermediate plant Mesembryanthemum crystallinum. Plant Physiol. Biochem. 2002, 40, 669–677. [Google Scholar] [CrossRef]
- Kalve, S.; Sarangi, B.K.; Pandey, R.A.; Chakrabarti, T. Arsenic and chromium hyperaccumulation by an ecotype of Pteris vittate—Prospective for phytoextraction from contaminated water and soil. Curr. Sci. 2011, 100, 888–894. [Google Scholar]
- Subhashini, V.; Swamy, A.V.V.S. Phytoremediation of lead, cadmium and chromium contaminated soils using selected weed plants. Acta Biol. Indica 2015, 4, 205–212. [Google Scholar]
- Gediga, K.; Spiak, Z.; Piszcz, U.; Bielecki, K. Suitability of different soil extractants for determination of available Cu and Mn contents in Polish soils. Commun. Soil Sci. Plant Anal. 2015, 46, 81–93. [Google Scholar] [CrossRef]
- Gembarzewski, H.; Korzeniowska, J. Simultaneous extraction of B, Cu, Fe, Mn, Mo and Zn from mineral soils and an estimation of the results. Agribiol. Res. 1990, 43, 115–127. [Google Scholar]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Treatment | Cr(VI) Dose [mmol per pot] | Plants f.w. [g] | Plants d.w. [g] | Roots d.w. [g/plant] | Shoots d.w. [g/plant] |
|---|---|---|---|---|---|
| Cr(VI) (K2CrO4) | |||||
| 0 | 13.03 ± 1.12 a | 1.16 ± 0.19 a | 0.07 ± 0.00 a | 1.09 ± 0.19 a | |
| 0.45 | 15.30 ± 0.20 a | 1.15 ± 0.06 a | 0.06 ± 0.01 a | 1.09 ± 0.05 a | |
| 0.90 | 12.52 ± 2.62 a | 0.99 ± 0.21 ab | 0.06 ± 0.00 a | 0.94 ± 0.21 a | |
| 2.30 | 11.09 ± 2.96 b | 1.05 ± 0.18 b | 0.07 ± 0.02 a | 0.99 ± 0.18 a | |
| 3.60 | 7.75 ± 3.49 b | 0.81 ± 0.39 b | 0.06 ± 0.02 a | 0.75 ± 0.37 a | |
| 4.50 | 4.06 ± 0.78 c | 0.91 ± 0.04 a | 0.06 ± 0.01 a | 0.85 ± 0.04 a | |
| Cr(VI) (K2Cr2O7) | |||||
| 0 | 13.03 ± 1.12 ab | 1.16 ± 0.19 a | 0.07 ± 0.00 b | 1.09 ± 0.19 a | |
| 0.90 | 15.53 ± 1.28 b | 1.16 ± 0.07 a | 0.05 ± 0.02 a | 1.11 ± 0.04 a | |
| 1.80 | 12.42 ± 2.05 a | 1.03 ± 0.11 ab | 0.06 ± 0.01 ab | 0.97 ± 0.11 ab | |
| 4.50 | 13.43 ± 1.46 ab | 1.18 ± 0.12 a | 0.06 ± 0.01 ab | 1.12 ± 0.12 a | |
| 7.20 | 3.67 ± 0.26 c | 0.88 ± 0.16 b | 0.06 ± 0.00 ab | 0.82 ± 0.16 b | |
| 9.00 | 4.26 ± 1.71 c | 1.01 ± 0.07 ab | 0.05 ± 0.00 a | 0.96 ± 0.08 ab |
| Treatment | Dose [mmol per pot] | Dose [mg kg−1 soil d.w.] | TF (shoot/root) |
|---|---|---|---|
| Cd2+ (CdCl2) | |||
| 0.0008 | 0.82 | 0.33 | |
| 0.008 | 8.20 | 0.33 | |
| 0.080 | 82.0 | 1.00 | |
| 0.800 | 818 | 0.21 | |
| Cr(VI) (K2CrO4) | |||
| 0.45 | 236 | 0.31 | |
| 0.90 | 425 | 0.34 | |
| 2.30 | 1086 | 1.86 | |
| 3.60 | 1699 | 0.86 | |
| 4.50 | 2124 | 1.99 | |
| Cr(VI) (K2Cr2O7) | |||
| 0.90 | 425 | 0.39 | |
| 1.80 | 850 | 0.25 | |
| 4.50 | 2124 | 1.72 | |
| 7.20 | 3398 | 2.05 | |
| 9.00 | 4248 | 2.29 |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Śliwa-Cebula, M.; Kaszycki, P.; Kaczmarczyk, A.; Nosek, M.; Lis-Krzyścin, A.; Miszalski, Z. The Common Ice Plant (Mesembryanthemum crystallinum L.)–Phytoremediation Potential for Cadmium and Chromate-Contaminated Soils. Plants 2020, 9, 1230. https://0-doi-org.brum.beds.ac.uk/10.3390/plants9091230
Śliwa-Cebula M, Kaszycki P, Kaczmarczyk A, Nosek M, Lis-Krzyścin A, Miszalski Z. The Common Ice Plant (Mesembryanthemum crystallinum L.)–Phytoremediation Potential for Cadmium and Chromate-Contaminated Soils. Plants. 2020; 9(9):1230. https://0-doi-org.brum.beds.ac.uk/10.3390/plants9091230
Chicago/Turabian StyleŚliwa-Cebula, Marta, Paweł Kaszycki, Adriana Kaczmarczyk, Michał Nosek, Agnieszka Lis-Krzyścin, and Zbigniew Miszalski. 2020. "The Common Ice Plant (Mesembryanthemum crystallinum L.)–Phytoremediation Potential for Cadmium and Chromate-Contaminated Soils" Plants 9, no. 9: 1230. https://0-doi-org.brum.beds.ac.uk/10.3390/plants9091230