
Microclimate and Vegetation Structure Significantly Affect Butterfly Assemblages in a Tropical Dry Forest
 , , , , and
, , , , and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Study Area and Sampling
2.2. Vegetation Sampling
2.3. Environmental Variable Measurements
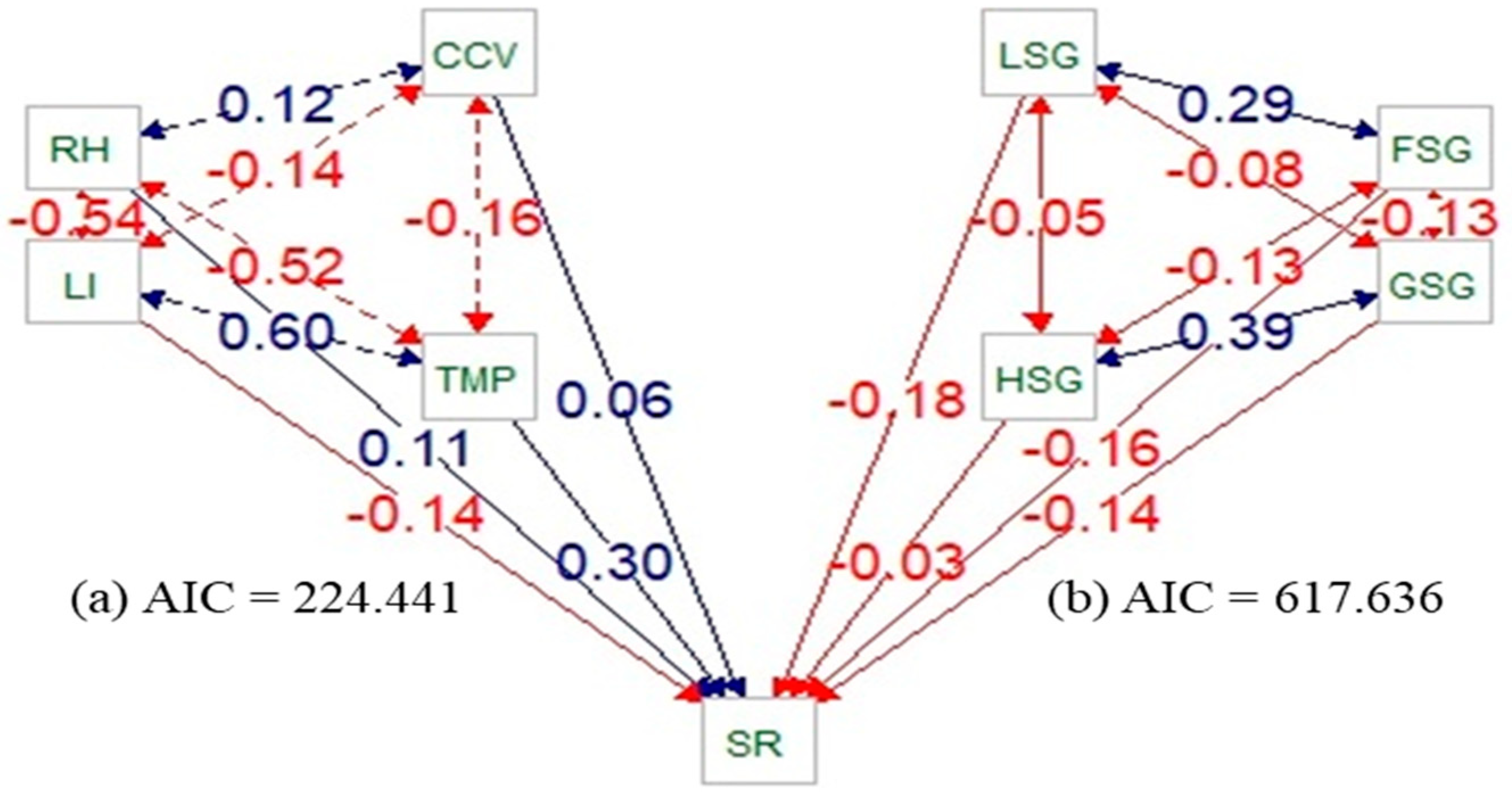
2.4. Statistical Analysis
3. Results
4. Discussion
5. Conservation Prioritization
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- WallisDeVries, M.F.; van Swaay, C.A.M. Global Warming and Excess Nitrogen May Induce Butterfly Decline by Microclimatic Cooling. Glob. Chang. Biol. 2006, 12, 1620–1626. [Google Scholar] [CrossRef]
- Dial, R.J.; Ellwood, M.D.F.; Turner, E.C.; Foster, W.A. Arthropod Abundance, Canopy Structure, and Microclimate in a Bornean Lowland Tropical Rain Forest. Biotropica 2006, 38, 643–652. [Google Scholar] [CrossRef]
- Geiger, R.; Aron, R.H.; Todhunter, P. The Climate near the Ground, 5th ed.; Harvard University Press: Wiesbaden, Germany, 1995; ISBN 978-3-322-86584-7. [Google Scholar]
- Meffe, G.K.; Carroll, C.R. Principles of Conservation Biology; Sinauer Associates, Inc.: Sunderland, MA, USA, 1994. [Google Scholar]
- Hellmann, J.J.; Weiss, S.B.; McLaughlin, J.F.; Ehrlich, P.R.; Murphy, D.D.; Launer, A.E. Structure and Dynamics of Euphydryas Editha Populations. In On the Wings of Checkerspots; Ehrlich, P.R., Hanski, I., Eds.; Oxford University Press: New York, NY, USA, 2004; pp. 34–62. [Google Scholar]
- Checa, M.F.; Rodriguez, J.; Willmott, K.R.; Liger, B. Microclimate Variability Significantly Affects the Composition, Abundance and Phenology of Butterfly Communities in a Highly Threatened Neotropical Dry Forest. Florida Entomol. 2014, 97, 1–13. [Google Scholar] [CrossRef]
- Dolia, J.; Devy, M.S.; Aravind, N.A.; Kumar, A. Adult Butterfly Communities in Coffee Plantations around a Protected Area in the Western Ghats, India. Anim. Conserv. 2008, 11, 26–34. [Google Scholar] [CrossRef]
- Honda, K.; Honda, K. Biology of Butterflies; University of Tokyo Press: Tokyo, Japan, 2005. [Google Scholar]
- Wang, L.; Wang, H.; Zha, Y.; Wei, H.; Chen, F.; Zeng, J. Forest Quality and Available Hostplant Abundance Limit the Canopy Butterfly of Teinopalpus Aureus. Insects 2022, 13, 1082. [Google Scholar] [CrossRef]
- Metzger, J.P. O Código Florestal Tem Base Científica? Nat. Conserv. 2010, 8, 92–99. [Google Scholar] [CrossRef]
- Ohwaki, A.; Maeda, S.; Kitahara, M.; Nakano, T. Associations between Canopy Openness, Butterfly Resources, Butterfly Richness and Abundance along Forest Trails in Planted and Natural Forests. Eur. J. Entomol. 2017, 114, 533–545. [Google Scholar] [CrossRef]
- Clench, H.K. Behavioral Thermoregulation in Butterflies. Ecology 1966, 47, 1021–1034. [Google Scholar] [CrossRef]
- Ehrlich, P.R.; Breedlove, D.E.; Brussard, P.F.; Sharp, M.A. Weather and the “Regulation” of Subalpine Populations. Ecology 1972, 53, 243–247. [Google Scholar] [CrossRef]
- Tsuji, J.S.; Kingsolver, J.G.; Watt, W.B. Thermal Physiological Ecology of Colias Butterflies in Flight. Oecologia 1986, 69, 161–170. [Google Scholar] [CrossRef]
- Murphy, D.D.; Freas, K.E.; Weiss, S.B. An Environment-metapopulation Approach to Population Viability Analysis for a Threatened Invertebrate. Conserv. Biol. 1990, 4, 41–51. [Google Scholar] [CrossRef]
- Pollard, E. Temperature, Rainfall and Butterfly Numbers. J. Appl. Ecol. 1988, 25, 819–828. [Google Scholar]
- Pollard, E.; Yates, T. Monitoring Butterflies for Ecology and Conservation; Springer: Dordrecht, The Netherlands, 1993. [Google Scholar]
- Gillespie, T.W.; Grijalva, A.; Farris, C.N. Diversity, Composition, and Structure of Tropical Dry Forests in Central America. Plant Ecol. 2000, 147, 37–47. [Google Scholar] [CrossRef]
- Dantas, C.; Zacca, T.; Bravo, F. Checklist of Butterflies (Lepidoptera: Papilionoidea) of an Urban Area of Caatinga-Atlantic Forest Ecotone in Bahia, Brazil. EntomoBrasilis 2021, 14, e959. [Google Scholar] [CrossRef]
- Suwarno, I.; Hanum, Y.; Yasmin, S.; Rasnovi, D. Diversity and Abundance of Butterfly (Lepidoptera Rhopalocera) in the City Garden of Banda Aceh, Indonesia. Ecol. Environ. Conserv. 2018, 24, 1009–1017. [Google Scholar]
- Koneri, R.; Nangoy, M.J.; Maabuat, P.V.; Saroyo, S.; Wakhid. Diversity and Composition of Butterflies in Three Habitats around Rayow Waterfall, Minahasa District, North Sulawesi, Indonesia. Biodiversitas 2022, 23, 1091–1098. [Google Scholar] [CrossRef]
- Nelson, S.M.; Andersen, D.C. Butterfly (Papilionoidea and Hesperioidea) Assemblages Associated with Natural, Exotic, and Restored Riparian Habitats along the Lower Colorado River, USA. River Res. Appl. 1999, 15, 485–504. [Google Scholar] [CrossRef]
- Douglas, W. Tallamy Do Alien Plants Reduce Insect Biomass? Conserv. Biol. 2004, 18, 1689–1692. [Google Scholar]
- Legal, L.; Valet, M.; Dorado, O.; de Jesus-Almonte, J.M.; López, K.; Céréghino, R. Lepidoptera Are Relevant Bioindicators of Passive Regeneration in Tropical Dry Forests. Diversity 2020, 12, 231. [Google Scholar] [CrossRef]
- Hilty, J.; Merenlender, A. Faunal Indicator Taxa Selection for Monitoring Ecosystem Health. Biol. Conserv. 2000, 92, 185–197. [Google Scholar] [CrossRef]
- Fleishman, E.; Blair, R.B.; Murphy, D.D. Empirical Validation of a Method for Umbrella Species Selection. Ecol. Appl. 2001, 11, 1489–1501. [Google Scholar] [CrossRef]
- Roberge, J.M.; Angelstam, P. Usefulness of the Umbrella Species Concept as a Conservation Tool. Conserv. Biol. 2004, 18, 76–85. [Google Scholar] [CrossRef]
- Rawat, G.S. Conservation Status of Forests and Wildlife in the Eastern Ghats, India. Environ. Conserv. 1997, 24, 307–315. [Google Scholar] [CrossRef]
- Rao, K.T.; Reddy, N.S.B.; Reddy, C.U. Eastern Ghats Environment Outlook; Greens’ Alliance for Conservation of Eastern Ghats: Hyderabad, India, 2019; ISBN 978-81-943739-0-2. [Google Scholar]
- Ramachandran, R.M.; Roy, P.S.; Chakravarthi, V.; Sanjay, J.; Joshi, P.K. Long-Term Land Use and Land Cover Changes (1920–2015) in Eastern Ghats, India: Pattern of Dynamics and Challenges in Plant Species Conservation. Ecol. Indic. 2018, 85, 21–36. [Google Scholar] [CrossRef]
- Sudhakar Reddy, C.; Ram Mohan Rao, K.; Pattanaik, C.; Joshi, P.K. Assessment of Large-Scale Deforestation of Nawarangpur District, Orissa, India: A Remote Sensing Based Study. Environ. Monit. Assess. 2009, 154, 325–335. [Google Scholar] [CrossRef]
- Adhikary, P.P.; Madhu, M.; Dash, C.J.; Sahoo, D.C.; Jakhar, P.; Naik, B.S.; Hombe Gowda, H.C.; Naik, G.B.; Dash, B. Prioritization of Traditional Tribal Field Crops Based on RWUE in Koraput District of Odisha. Indian J. Tradit. Knowl. 2015, 1, 88–95. [Google Scholar]
- Adhikary, P.P.; Barman, D.; Madhu, M.; Dash, C.J.; Jakhar, P.; Hombegowda, H.C.; Naik, B.S.; Sahoo, D.C.; Beer, K. Land Use and Land Cover Dynamics with Special Emphasis on Shifting Cultivation in Eastern Ghats Highlands of India Using Remote Sensing Data and GIS. Environ. Monit. Assess. 2019, 191, 315. [Google Scholar] [CrossRef]
- Sawford, B. The Butterflies of Hertfordshire; Castlemead Publications: Ware, UK, 1987. [Google Scholar]
- Videvall, E.; Öckinger, E.; Pettersson, L.B. Butterfly Monitoring Using Systematically Placed Transects in Contrasting Climatic Regions—Exploring an Established Spatial Design for Sampling. Nat. Conserv. 2016, 14, 41–62. [Google Scholar] [CrossRef]
- Zografou, K.; Wilson, R.J.; Halley, J.M.; Tzirkalli, E.; Kati, V. How Are Arthopod Communities Structured and Why Are They so Diverse? Answers from Mediterranean Mountains Using Hierarchical Additive Partitioning. Biodivers. Conserv. 2017, 26, 1333–1351. [Google Scholar] [CrossRef]
- Kehimkar, I. The book of Indian Butterflies, 2nd ed.; BNHS and Oxford University Press: Mumbai, India, 2016. [Google Scholar]
- Wynter-Blyth, M.A. Butterflies of the Indian Region; Bombay Nat. Hist. Soc.: Bombay, India, 1957. [Google Scholar]
- Varshney, R.K.; Smetacek, P. A Synoptic Catalogue of the Butterflies of India; Butterfly Research Centre, Bhimtal and Indinov Publishing: New Delhi, India, 2015; ISBN 78-81-929826-4-9. [Google Scholar]
- Indian Meteorological Department. Rainfall and Temperature Trends. Available online: http://www.imdpune.gov.in (accessed on 2 September 2020).
- Bhardwaj, M.; Uniyal, V.P.; Sanyal, A.K.; Singh, A.P. Butterfly Communities along an Elevational Gradient in the Tons Valley, Western Himalayas: Implications of Rapid Assessment for Insect Conservation. J. Asia. Pac. Entomol. 2012, 15, 207–217. [Google Scholar] [CrossRef]
- Forest Survey of India. The Manual of Instructions for Field Inventory; Ministry of Environment and Forests: Dehradun, India, 2002.
- Tichý, L. Field Test of Canopy Cover Estimation by Hemispherical Photographs Taken with a Smartphone. J. Veg. Sci. 2016, 27, 427–435. [Google Scholar] [CrossRef]
- Bianchi, S.; Cahalan, C.; Hale, S.; Gibbons, J.M. Rapid Assessment of Forest Canopy and Light Regime Using Smartphone Hemispherical Photography. Ecol. Evol. 2017, 7, 10556–10566. [Google Scholar] [CrossRef] [PubMed]
- Mahata, A.; Samal, K.T.; Palita, S.K. Butterfly Diversity in Agroforestry Plantations of Eastern Ghats of Southern Odisha, India. Agrofor. Syst. 2019, 93, 1423–1438. [Google Scholar] [CrossRef]
- Novotny, V.; Miller, S.E.; Leps, J.; Basset, Y.; Bito, D.; Janda, M.; Hulcr, J.; Damas, K.; Weiblen, G.D. No Tree an Island: The Plant-Caterpillar Food Web of a Secondary Rain Forest in New Guinea. Ecol. Lett. 2004, 7, 1090–1100. [Google Scholar] [CrossRef]
- Brito, M.M.; Ribeiro, D.B.; Raniero, M.; Hasui, É.; Ramos, F.N.; Arab, A. Functional Composition and Phenology of Fruit-Feeding Butterflies in a Fragmented Landscape: Variation of Seasonality between Habitat Specialists. J. Insect Conserv. 2014, 18, 547–560. [Google Scholar] [CrossRef]
- Itoh, A.; Ohkubo, T.; Nanami, S.; Tan, S.; Yamakura, T. Comparison of Statistical Tests for Habitat Associations in Tropical Forests: A Case Study of Sympatric Dipterocarp Trees in a Bornean Forest. For. Ecol. Manag. 2010, 259, 323–332. [Google Scholar] [CrossRef]
- Noguchi, H.; Itoh, A.; Mizuno, T.; Sri-Ngernyuang, K.; Kanzaki, M.; Teejuntuk, S.; Sungpalee, W.; Hara, M.; Ohkubo, T.; Sahunalu, P.; et al. Habitat Divergence in Sympatric Fagaceae Tree Species of a Tropical Montane Forest in Northern Thailand. J. Trop. Ecol. 2007, 23, 549–558. [Google Scholar] [CrossRef]
- Dufrêne, M.; Legendre, P. Species Assemblages and Indicator Species: The Need for a Flexible Asymmetrical Approach. Ecol. Monogr. 1997, 67, 345–366. [Google Scholar] [CrossRef]
- De Cáceres, M.; Legendre, P.; Moretti, M. Improving Indicator Species Analysis by Combining Groups of Sites. Oikos 2010, 119, 1674–1684. [Google Scholar] [CrossRef]
- De Cáceres, M.; Legendre, P. Associations between Species and Groups of Sites: Indices and Statistical Inference. Ecology 2009, 90, 3566–3574. [Google Scholar] [CrossRef]
- Budka, A.; Łacka, A.; Szoszkiewicz, K. The Use of Rarefaction and Extrapolation as Methods of Estimating the Effects of River Eutrophication on Macrophyte Diversity. Biodivers. Conserv. 2019, 28, 385–400. [Google Scholar] [CrossRef]
- Hill, M.O. Diversity and Evenness: A Unifying Notation and Its Consequences. Ecology 1973, 54, 427–432. [Google Scholar] [CrossRef]
- Chao, A.; Gotelli, N.J.; Hsieh, T.C.; Sander, E.L.; Ma, K.H.; Colwell, R.K.; Ellison, A.M. Rarefaction and Extrapolation with Hill Numbers: A Framework for Sampling and Estimation in Species Diversity Studies. Ecol. Monogr. 2014, 84, 45–67. [Google Scholar] [CrossRef]
- Hsieh, T.C.; Ma, K.H.; Chao, A. INEXT: An R Package for Rarefaction and Extrapolation of Species Diversity (Hill Numbers). Methods Ecol. Evol. 2016, 7, 1451–1456. [Google Scholar] [CrossRef]
- Rosseel, Y. Lavaan: An R Package for Structural Equation Modeling. J. Stat. Softw. 2012, 48, 1–36. [Google Scholar] [CrossRef]
- Epskamp, S.; Stuber, S.; Nak, J.; Veenman, M.; Jorgensen, T.D. Package ‘SemPlot’: Path Diagrams and Visual Analysis of Various SEM Packages’ Output, Version 1.1.6; Github Repository. 2022. Available online: https://github.com/SachaEpskamp/semPlot (accessed on 15 October 2023).
- Ter Braak, C.J.F. The Analysis of Vegetation-Environment Relationships by Canonical Correspondence Analysis. Vegetatio 1987, 69, 69–77. [Google Scholar] [CrossRef]
- Oksanen, R.J.; Simpson, G.L.; Blanchet, F.G.; Solymos, P.; Stevens, M.H.H.; Szoecs, E.; Wagner, H.; Barbour, M.; Bedward, M.; Bolker, B.; et al. Community Ecology Package. Vegan Community Ecol. Packag. 2022. [Google Scholar]
- Oksanen, J.; Kindt, R.; Simpson, G.L.; Murdoch, D. Package ‘Vegan3d’: Static and Dynamic 3D Plots for the “Vegan” Package, Version 1.2-0; Github Repository. 2022. Available online: https://github.com/vegandevs/vegan3d (accessed on 15 October 2023).
- R Core Team. R: A Language and Environment for Statistical Computing; R Core Team: Vienna, Austria, 2022. [Google Scholar]
- De Cáceres, M.; Legendre, P.; Wiser, S.K.; Brotons, L. Using Species Combinations in Indicator Value Analyses. Methods Ecol. Evol. 2012, 3, 973–982. [Google Scholar] [CrossRef]
- Kunte, K. Butterflies of Peninsular India; Universities Press: Hyderabad, India, 2000; ISBN 9788173713545. [Google Scholar]
- Guadalquiver, D.M.; Nuneza, O.M.; Dupo, A.L. Species Diversity of Lepidoptera in Mimbilisan Protected Landscape, Misamis Oriental, Philippines. Entomol. Appl. Sci. Lett. 2019, 6, 33–47. [Google Scholar]
- Borror, D.J.; Triplehorn, C.A.; Johnson, N.F. An Introduction to the Study of Insects; Saunders College Publishing: Philadelphia, PA, USA, 1989; ISBN 0-03-025397-7. [Google Scholar]
- Li, X.; Jia, X.; Xiang, H.; Diao, H.; Yan, Y.; Wang, Y.; Ma, R.; Leskey, T. The Effect of Photoperiods and Light Intensity on Mating Behavior and Reproduction of Grapholita Molesta (Lepidoptera: Tortricidae). Environ. Entomol. 2019, 48, 1035–1041. [Google Scholar] [CrossRef]
- Hill, G.M.; Kawahara, A.Y.; Daniels, J.C.; Bateman, C.C.; Scheffers, B.R. Climate Change Effects on Animal Ecology: Butterflies and Moths as a Case Study. Biol. Rev. 2021, 96, 2113–2126. [Google Scholar] [CrossRef] [PubMed]
- Ghazoul, J. Impact of Logging on the Richness and Diversity of Forest Butterflies in a Tropical Dry Forest in Thailand. Biodivers. Conserv. 2002, 11, 521–541. [Google Scholar] [CrossRef]
- Weiss, A.A. Estimating Nonlinear Dynamic Models Using Least Absolute Error Estimation. Econom. Theory 1991, 7, 46–68. [Google Scholar] [CrossRef]
- Barlow, J.; Overal, W.L.; Araujo, I.S.; Gardner, T.A.; Peres, C.A. The Value of Primary, Secondary and Plantation Forests for Fruit-Feeding Butterflies in the Brazilian Amazon. J. Appl. Ecol. 2007, 44, 1001–1012. [Google Scholar] [CrossRef]
- Das, K.S.A.; Radhamany, D.; Molleman, F. Community Structure in an Isolated Tropical Forest Biome: One Year of Fruit-Feeding Butterfly Trapping in Four Habitats in the Western Ghats, India. Diversity 2023, 15, 36. [Google Scholar] [CrossRef]
- Devries, P.J.; Walla, T.R. Species Diversity and Community Structure in Neotropical Fruit-Feeding Butterflies. Biol. J. Linn. Soc. 2001, 74, 1–15. [Google Scholar] [CrossRef]
- McCain, C.M. The Mid-Domain Effect Applied to Elevational Gradients: Species Richness of Small Mammals in Costa Rica. J. Biogeogr. 2004, 31, 19–31. [Google Scholar] [CrossRef]
- Ren, J.; Li, S.; He, M.; Zhang, Y. Butterfly Community Diversity in the Qinling Mountains. Diversity 2022, 14, 27. [Google Scholar] [CrossRef]
- Gallou, A.; Baillet, Y.; Ficetola, G.F.; Després, L. Elevational Gradient and Human Effects on Butterfly Species Richness in the French Alps. Ecol. Evol. 2017, 7, 3672–3681. [Google Scholar] [CrossRef]
- Maes, D.; Van Dyck, H. Habitat Quality and Biodiversity Indicator Performances of a Threatened Butterfly versus a Multispecies Group for Wet Heathlands in Belgium. Biol. Conserv. 2005, 123, 177–187. [Google Scholar] [CrossRef]
- An, J.-S.; Choi, S.-W. Butterflies as an Indicator Group of Riparian Ecosystem Assessment. J. Asia. Pac. Entomol. 2021, 24, 195–200. [Google Scholar] [CrossRef]
- Brown, K.S., Jr. Insetos Indicadores Da História. In Matas Ciliares: Conservação e Recuperação; Rodrigues, R.R., Leitão-Filho, H.F., Eds.; Editora da Universidade da São Paulo: São Paulo, Brzail, 2000; pp. 223–232. [Google Scholar]
- Spitzer, K.; Jaros, J.; Havelka, J.; Leps, J. Effect of Small-Scale Disturbance on Butterfly Communities of an Indochinese Montane Rainforest. Biol. Conserv. 1997, 80, 9–15. [Google Scholar] [CrossRef]
- Hill, J.; Hamer, K.; Tangah, J.; Dawood, M. Ecology of Tropical Butterflies in Rainforest Gaps. Oecologia 2001, 128, 294–302. [Google Scholar] [CrossRef] [PubMed]
- Blair, R.B.; Launer, A.E. Butterfly Diversity and Human Land Use: Species Assemblages along an Urban Gradient. Biol. Conserv. 1997, 3207, 113–125. [Google Scholar] [CrossRef]
- Van Vu, L.; Vu, C.Q.; Quang Vu, C. Diversity Pattern of Butterfly Communities (Lepidoptera, Papilionoidae) in Different Habitat Types in a Tropical Rain Forest of Southern Vietnam. ISRN Zool. 2011, 2011, 818545. [Google Scholar] [CrossRef]
- Hamer, K.C.; Hill, J.K.; Benedick, S.; Mustaffa, N.; Sherratt, T.N.; Maryati, M.; Chey, V.K. Ecology of Butterflies in Natural and Selectively Logged Forests of Northern Borneo: The Importance of Habitat Heterogeneity. J. Appl. Ecol. 2003, 40, 150–162. [Google Scholar] [CrossRef]
- Reddy, C.S.; Jha, C.S.; Dadhwal, V.K. Assessment and Monitoring of Long-Term Forest Cover Changes in Odisha, India Using Remote Sensing and GIS. Environ. Monit. Assess. 2013, 185, 4399–4415. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Scientific Name | Common Name | Habitat Specificity Index | Habitat Specificity | Relative Abundance | ||
|---|---|---|---|---|---|---|
| Open Forests | Riparian Forests | Dense Forests | ||||
| Family: Hesperiidae | ||||||
| Subfamily: Coeliadinae | ||||||
| Hasora chromus | Common Banded Awl | 0.806 | SHP | 0.24 | 0.99 | 6.46 |
| Subfamily: Hesperiinae | ||||||
| Iambrix salsala | Chestnut Bob | 0.80 | SHP | 0.12 | 0 | 1.03 |
| Subfamily: Pyrginae | ||||||
| Sarangesa purendra | Spotted Small Flat | 1 | RFS | 0 | 0.50 | 0 |
| Sarangesa dasahara | Common Small Flat | 1 | RFS | 0 | 0.25 | 0 |
| Tagiades japetus | Suffused Snow Flat | 1 | RFS | 0 | 0.25 | 0 |
| Tagiades litigiosa | Water Snow Flat | 1 | DFS | 0 | 0 | 0.26 |
| Family: Lycaenidae | ||||||
| Subfamily: Curetinae | ||||||
| Curetis acuta | Angled Sunbeam | 1 | RFS | 0 | 0.25 | 0 |
| Subfamily: Polyommatinae | ||||||
| Acytolepis puspa | Common Hedge Blue | 1 | OFS | 0.12 | 0 | 0 |
| Anthene emolus | Ciliate Blue | 1 | RFS | 0 | 0.25 | 0 |
| Caleta decidia | Angled Pierrot | 0.417 | Generalist | 0.61 | 0.99 | 0.78 |
| Castalius rosimon | Common Pierrot | 0.8 | SHP | 0.12 | 0.99 | 0 |
| Catochryspos strabo | Forget-Me-Not | 1 | OFS | 0.85 | 0 | 0 |
| Chilades pandava | Plains Cupid | 0.444 | Generalist | 0.36 | 0.99 | 0.52 |
| Freyeria trochylus | Grass Jewel | 1 | OFS | 0.49 | 0 | 0 |
| Jamides bochus | Dark Cerulean | 0.768657 | SHP | 2.67 | 2.23 | 26.61 |
| Jamides celeno | Common Cerulean | 0.534884 | SHP | 5.58 | 5.71 | 4.39 |
| Lampides boeticus | Peablue | 1 | OFS | 0.12 | 0 | 0 |
| Neopithecops zalmora | Quaker | 1 | RFS | 0 | 0.25 | 0 |
| Prosotas nora | Common Lineblue | 1 | RFS | 0 | 0.25 | 0 |
| Pseudozizeeria maha | Pale Grass Blue | 0.606 | SHP | 2.43 | 0.99 | 2.33 |
| Zizeeria karsandra | Dark Grass Blue | 0.4 | Generalist | 0.24 | 0.25 | 0.52 |
| Zizula hylax | Tiny Grass Blue | 0.517 | SHP | 1.70 | 3.72 | 0 |
| Subfamily: Theclinae | ||||||
| Amblypodia anita | Purple Leaf Blue | 0.857 | SHP | 0.73 | 0.25 | 0 |
| Arhopalini amantes | Large Oakblue | 0.939 | OFS * | 5.58 | 0.74 | 0 |
| Loxura atymnus | Yam Fly | 0.5 | SHP | 0.24 | 0.74 | 1.29 |
| Spindasis syama | Club Silverline | 0.667 | SHP | 0.49 | 0 | 0.52 |
| Spindasis vulcanus | Common Silverline | 0.75 | SHP | 0.36 | 0 | 0.26 |
| Zeltus amasa | Fluffy Tit | 1 | RFS | 0 | 0.50 | 0 |
| Family: Nymphalidae | ||||||
| Subfamily: Biblidinae | ||||||
| Ariadne ariadne | Angled Castor | 0.667 | SHP | 0.49 | 0.50 | 0 |
| Ariadne merione | Common Castor | 0.9 | OFS * | 1.09 | 0.25 | 0 |
| Subfamily: Charaxinae | ||||||
| Charaxes bernardus | Tawny Rajah | 1 | RFS | 0 | 0.25 | 0 |
| Polyura athamas | Common Nawab | 1 | OFS | 0.24 | 0 | 0 |
| Subfamily: Cyrestinae | ||||||
| Cyrestis thyodamas | Common Map | 0.667 | SHP | 0.24 | 0 | 0.26 |
| Subfamily: Danainae | ||||||
| Danaus chrysippus | Plain Tiger | 0.694 | SHP | 3.03 | 2.23 | 0.52 |
| Danaus genutia | Common/Striped Tiger | 0.458 | Generalist | 1.33 | 1.74 | 1.55 |
| Euploea core | Common Crow | 0.398 | Generalist | 9.22 | 15.14 | 13.95 |
| Parantica aglea | Glassy Tiger | 0.538 | SHP | 0.36 | 1.74 | 0.76 |
| Tirumala limniace | Blue Tiger | 1 | RFS | 0 | 0.50 | 0 |
| Subfamily: Heliconiinae | ||||||
| Acraea violae | Tawny Coster | 0.60 | SHP | 0.24 | 0 | 0.78 |
| Phalanta phalantha | Common Leopard | 0.69 | SHP | 1.33 | 0.99 | 0.26 |
| Vagrans egista | Vagrant | 0.50 | SHP | 0.12 | 0.25 | 0 |
| Subfamily: Limenitidinae | ||||||
| Athyma perius | Common Sergeant | 1 | RFS | 0 | 0.25 | 0 |
| Euthalia aconthea | Common Baron | 1 | OFS | 0.24 | 0 | 0 |
| Euthalia nais | Baronet | 0.46 | Generalist | 1.33 | 2.48 | 0.78 |
| Neptis hylas | Common Sailer | 0.55 | SHP | 1.33 | 1.49 | 0.78 |
| Neptis jumbah | Chestnut-Streake Sailer | 0.75 | SHP | 0.12 | 0 | 0.78 |
| Pantoporia hordonia | Common Lascar | 0.50 | SHP | 0.12 | 0 | 0.26 |
| Tanaecia lepidea | Grey Count | 0.50 | SHP | 0 | 0.25 | 0.26 |
| Subfamily: Nymphalinae | ||||||
| Hypolimnus bolina | Great Eggfly | 0.67 | SHP | 0 | 0.50 | 0.26 |
| Junonia almana | Peacock Pansy | 0.50 | SHP | 0.36 | 0.25 | 0.52 |
| Junonia atlites | Grey Pansy | 1 | RFS | 0 | 1.24 | 0 |
| Junonia iphita | Chocolate Pansy | 0.43 | Generalist | 3.28 | 3.47 | 5.68 |
| Junonia lemonias | Lemon Pansy | 0.74 | SHP | 4.49 | 2.48 | 0.78 |
| Junonia orithiya | Blue Pansy | 1 | OFS | 0.12 | 0 | 0 |
| Kallima inachus | Orange Oakleaf | 0.80 | SHP | 0 | 0.99 | 0.26 |
| Vanessa cardui | Painted lady | 1 | DFS | 0 | 0 | 0.26 |
| Subfamily: Satyrinae | ||||||
| Melanitis leda | Common Evening Brown | 0.50 | SHP | 0.49 | 0.50 | 1.55 |
| Mycalesis perseus | Common Bushbrown | 0.69 | SHP | 0.24 | 2.23 | 0.52 |
| Ypthima huebneri | Common Fourring | 0.52 | SHP | 1.82 | 3.47 | 0 |
| Family: Papilionidae | ||||||
| Subfamily: Papilioninae | ||||||
| Atrophaneura aristolochiae | Common Rose | 0.50 | SHP | 0.12 | 0.25 | 0.52 |
| Graphium agamemnon | Tailed Jay | 0.67 | SHP | 0.24 | 0 | 0.26 |
| Graphium doson | Common Jay | 1 | OFS | 0.49 | 0 | 0 |
| Graphium nomius | Spot Swordtail | 0.78 | SHP | 2.55 | 0.99 | 0.52 |
| Papilio clytia | Common Mime | 0.5 | SHP | 0.36 | 0 | 0.78 |
| Papilio crino | Common Banded Peacock | 1 | OFS | 0.24 | 0 | 0 |
| Papilio demoleus | Lime Butterfly | 0.66 | SHP ** | 14.56 | 12.6 | 2.58 |
| Papilio hector | Crimson Rose | 1 | DFS | 0 | 0 | 0.26 |
| Papilio paris | Paris Peacock | 1 | OFS | 0.24 | 0 | 0 |
| Papilio polymnestor | Blue Mormon | 0.89 | SHP | 0.12 | 1.99 | 0 |
| Papilio polytes | Common Mormon | 0.52 | SHP | 3.40 | 2.48 | 4.13 |
| Family: Pieridae | ||||||
| Subfamily: Coliadinae | ||||||
| Catopsilia pomona | Common Emigrant | 0.51 | SHP | 3.28 | 8.44 | 1.55 |
| Catopsilia pyranthe | Mottled Emigrant | 0.60 | SHP ** | 10.19 | 3.23 | 11.11 |
| Eurema hecabe | Common Grass Yellow | 0.69 | SHP | 4.37 | 3.72 | 0.26 |
| Gandaca harina | Tree Yellow | 0.82 | SHP | 2.18 | 0.74 | 0.26 |
| Subfamily: Pierinae | ||||||
| Cepora nerissa | Common Gull | 0.50 | SHP | 0.24 | 0.25 | 0.26 |
| Delias hyparete | Painted Jezabel | 0.88 | SHP | 2.67 | 0 | 0.78 |
| Ixias pyrene | Yellow Orange Tip | 1 | RFS | 0 | 0.25 | 0 |
| Leptosia nina | Psyche | 1 | RFS | 0 | 0.50 | 0 |
| Family: Riodinidae | ||||||
| Subfamily: Riodininae | ||||||
| Abisara echerius | Plum Judy | 0.8 | SHP | 0 | 0.25 | 1.03 |
| Species Combinations | A | B | Stat | p-Value |
|---|---|---|---|---|
| Riparian Forests (No. of Species = 18) | ||||
| Jamides celeno + Eurema hecabe | 0.713 | 0.8571 | 0.782 | 0.003 ** |
| Eurema hecabe | 0.5268 | 0.8571 | 0.672 | 0.045 * |
| Euploea core + Eurema hecabe | 0.6202 | 0.7143 | 0.666 | 0.023 * |
| Junonia iphita + Eurema hecabe | 0.7004 | 0.5714 | 0.633 | 0.030 * |
| Phalanta phalantha + Catopsilia pomona | 0.8108 | 0.4286 | 0.589 | 0.038 * |
| Junonia iphita + Mycalesis perseus | 0.7921 | 0.4286 | 0.583 | 0.021 * |
| Mycalesis perseus | 0.786 | 0.4286 | 0.58 | 0.049 * |
| Mycalesis perseus + Euploea core | 0.7605 | 0.4286 | 0.571 | 0.029 * |
| Caleta decidia + Eurema hecabe | 1 | 0.2857 | 0.535 | 0.044 * |
| Castalius rosimon + Catopsilia pomona | 1 | 0.2857 | 0.535 | 0.038 * |
| Arhopalini amantes + Mycalesis perseus | 1 | 0.2857 | 0.535 | 0.030 * |
| Arhopalini amantes + Parantica aglea | 1 | 0.2857 | 0.535 | 0.030 * |
| Zizula hylax + Parantica aglea | 1 | 0.2857 | 0.535 | 0.033 * |
| Mycalesis perseus + Ypthima huebneri | 1 | 0.2857 | 0.535 | 0.033 * |
| Phalanta phalantha + Neptis hylas | 1 | 0.2857 | 0.535 | 0.032 * |
| Phalanta phalantha + Graphium nomius | 1 | 0.2857 | 0.535 | 0.033 * |
| Ypthima huebneri + Parantica aglea | 0.9449 | 0.2857 | 0.535 | 0.050 * |
| Dense Forests (No. of Species = 8) | ||||
| Hasora chromus + Catopsilia pyranthe | 0.9756 | 0.5 | 0.698 | 0.006 ** |
| Hasora chromus | 0.8231 | 0.5 | 0.642 | 0.018 * |
| Hasora chromus + Pseudozizeeria maha | 1 | 0.375 | 0.612 | 0.009 ** |
| Pseudozizeeria maha + Melanitis leda | 1 | 0.375 | 0.612 | 0.016 * |
| Melanitis leda + Papilio polytes | 0.9259 | 0.375 | 0.589 | 0.04 |
| Pseudozizeeria maha + Papilio polytes | 0.6863 | 0.5 | 0.586 | 0.043 * |
| Parantica aglea + Papilio polytes | 0.8824 | 0.375 | 0.575 | 0.041 * |
| Pseudozizeeria maha + Danaus genutia | 0.7143 | 0.375 | 0.518 | 0.035 * |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Mahata, A.; Panda, R.M.; Dash, P.; Naik, A.; Naik, A.K.; Palita, S.K. Microclimate and Vegetation Structure Significantly Affect Butterfly Assemblages in a Tropical Dry Forest. Climate 2023, 11, 220. https://0-doi-org.brum.beds.ac.uk/10.3390/cli11110220
Mahata A, Panda RM, Dash P, Naik A, Naik AK, Palita SK. Microclimate and Vegetation Structure Significantly Affect Butterfly Assemblages in a Tropical Dry Forest. Climate. 2023; 11(11):220. https://0-doi-org.brum.beds.ac.uk/10.3390/cli11110220
Chicago/Turabian StyleMahata, Anirban, Rajendra Mohan Panda, Padmanava Dash, Ayusmita Naik, Alok Kumar Naik, and Sharat Kumar Palita. 2023. "Microclimate and Vegetation Structure Significantly Affect Butterfly Assemblages in a Tropical Dry Forest" Climate 11, no. 11: 220. https://0-doi-org.brum.beds.ac.uk/10.3390/cli11110220