Serum Glycoproteomic Alterations in Patients with Diabetic Retinopathy

Abstract
:1. Introduction
2. Materials and Methods
2.1. Human Subjects and Sample Collection
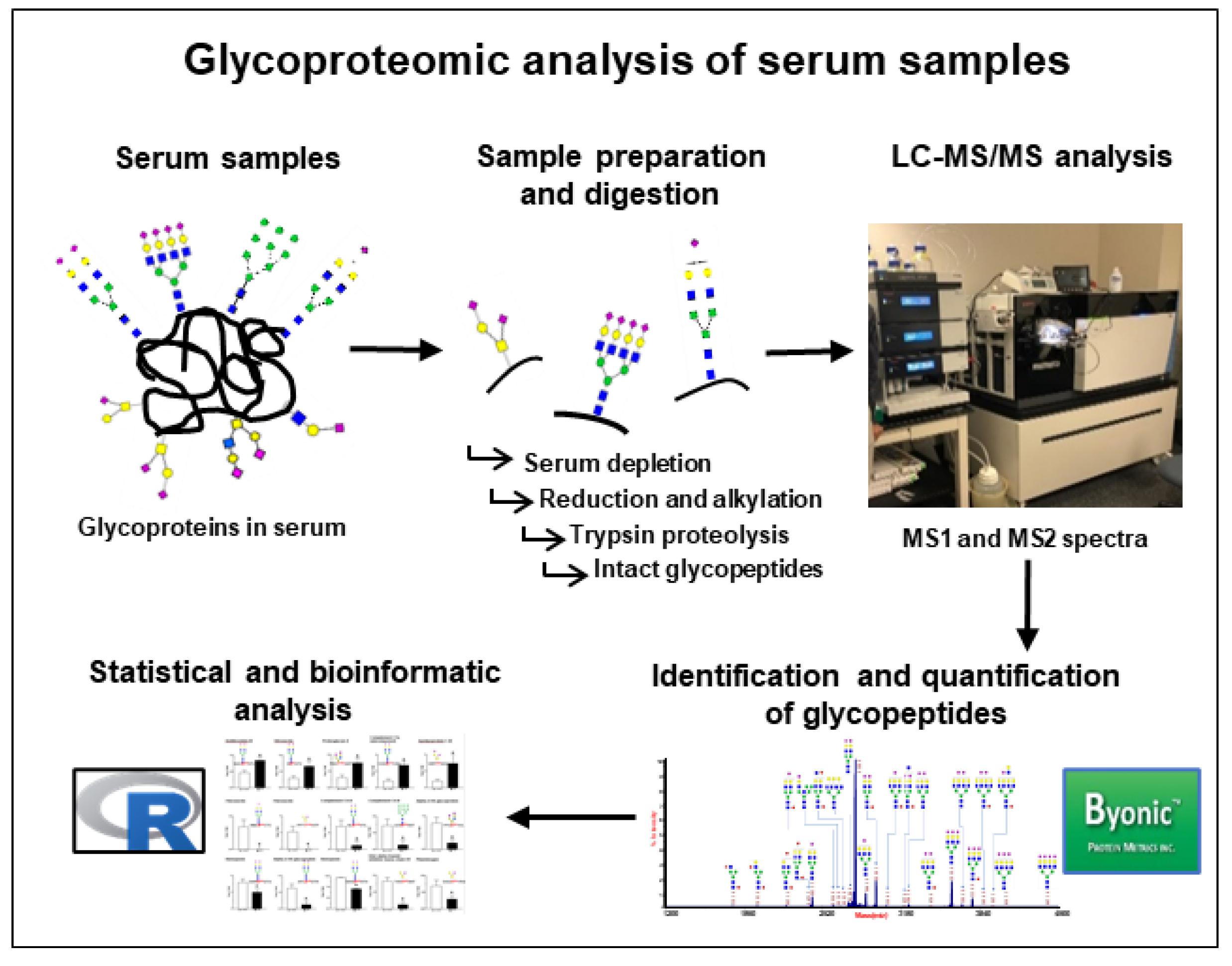
2.2. Sample Preparation and Digestion
2.3. LC–MS/MS Analysis
2.4. Protein Identification and Quantification
2.5. Statistical and Bioinformatics Analysis
3. Results
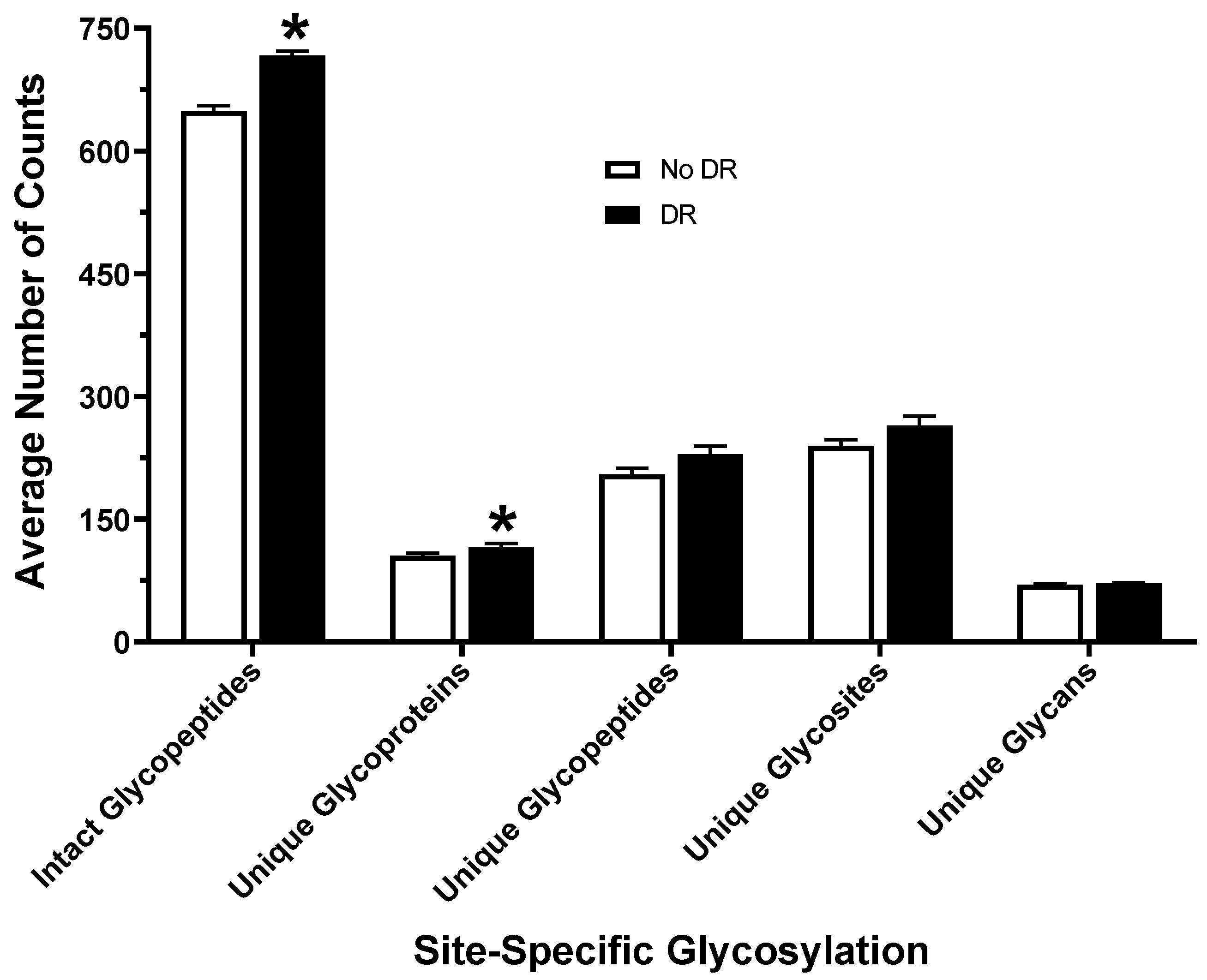
3.1. Glycosylation Profiling of Human Serum
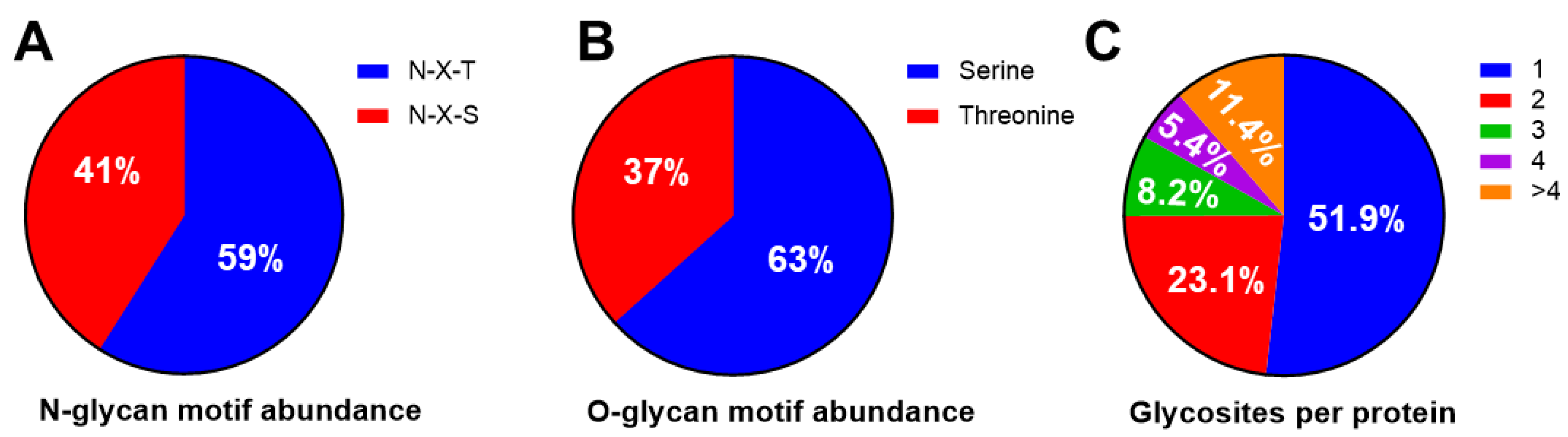
3.2. N-Glycans versus O-Glycans
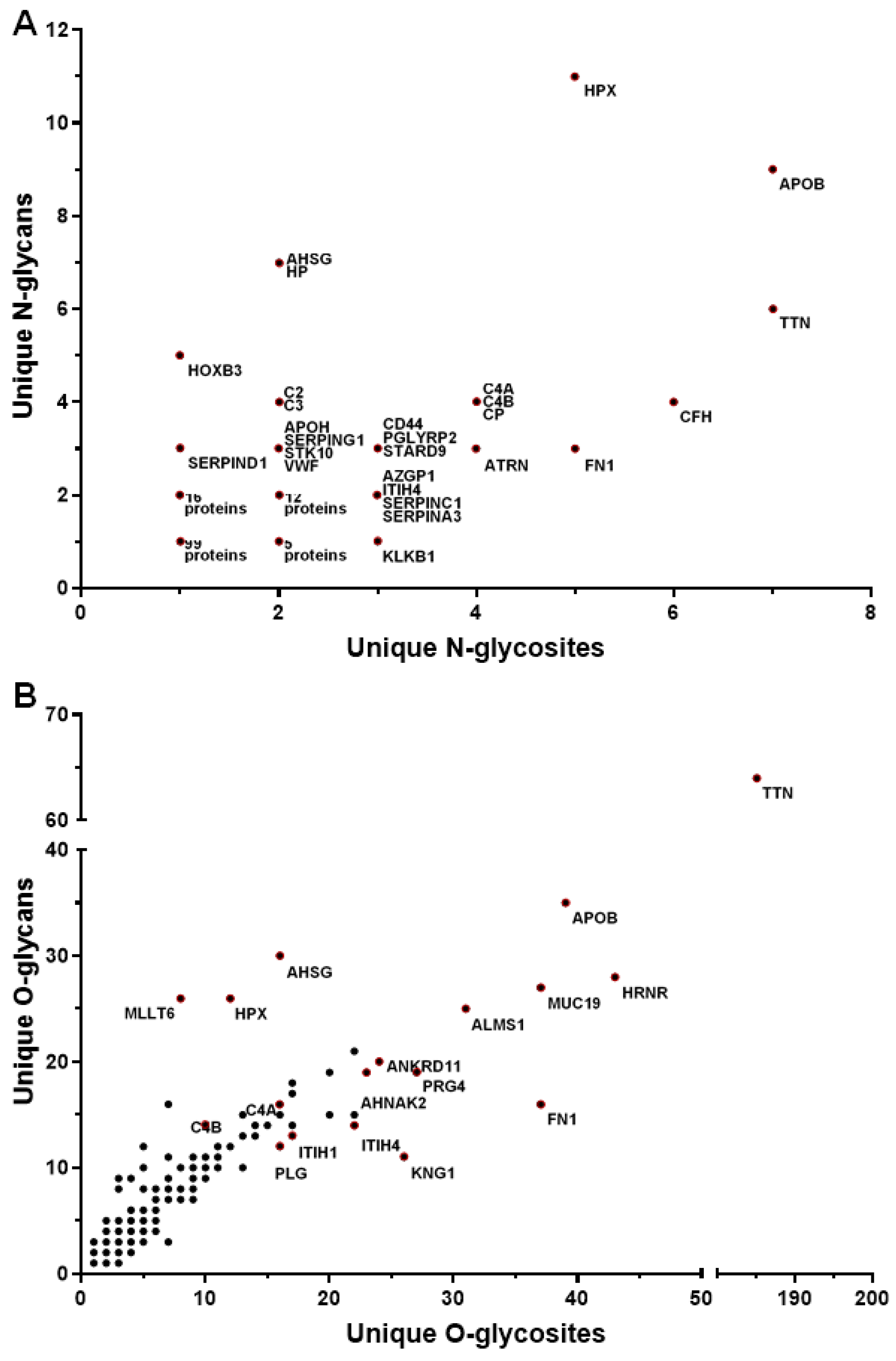
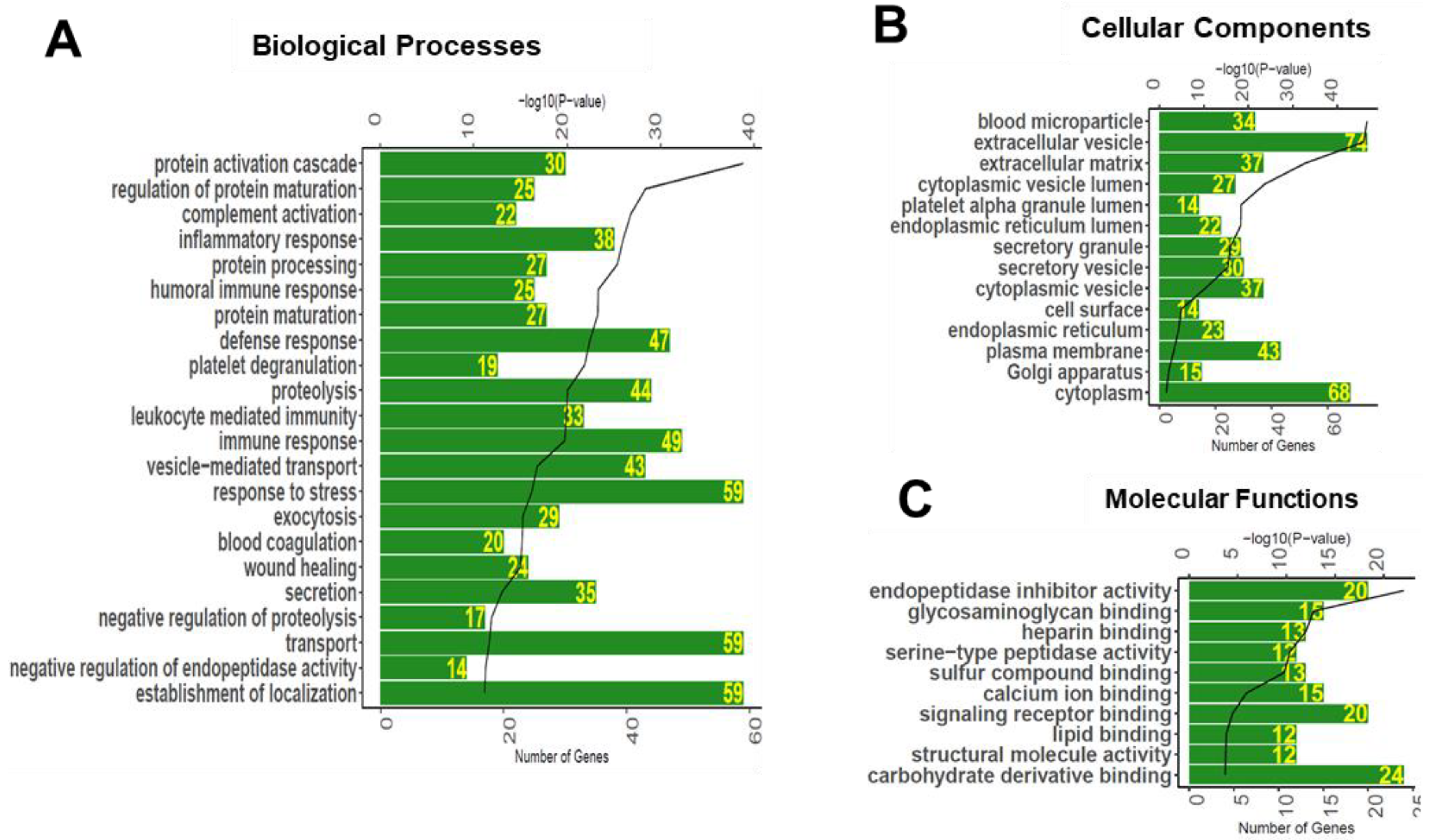
3.3. Gene Ontology Enrichment Analyses of Glycoproteins Detected in Serum
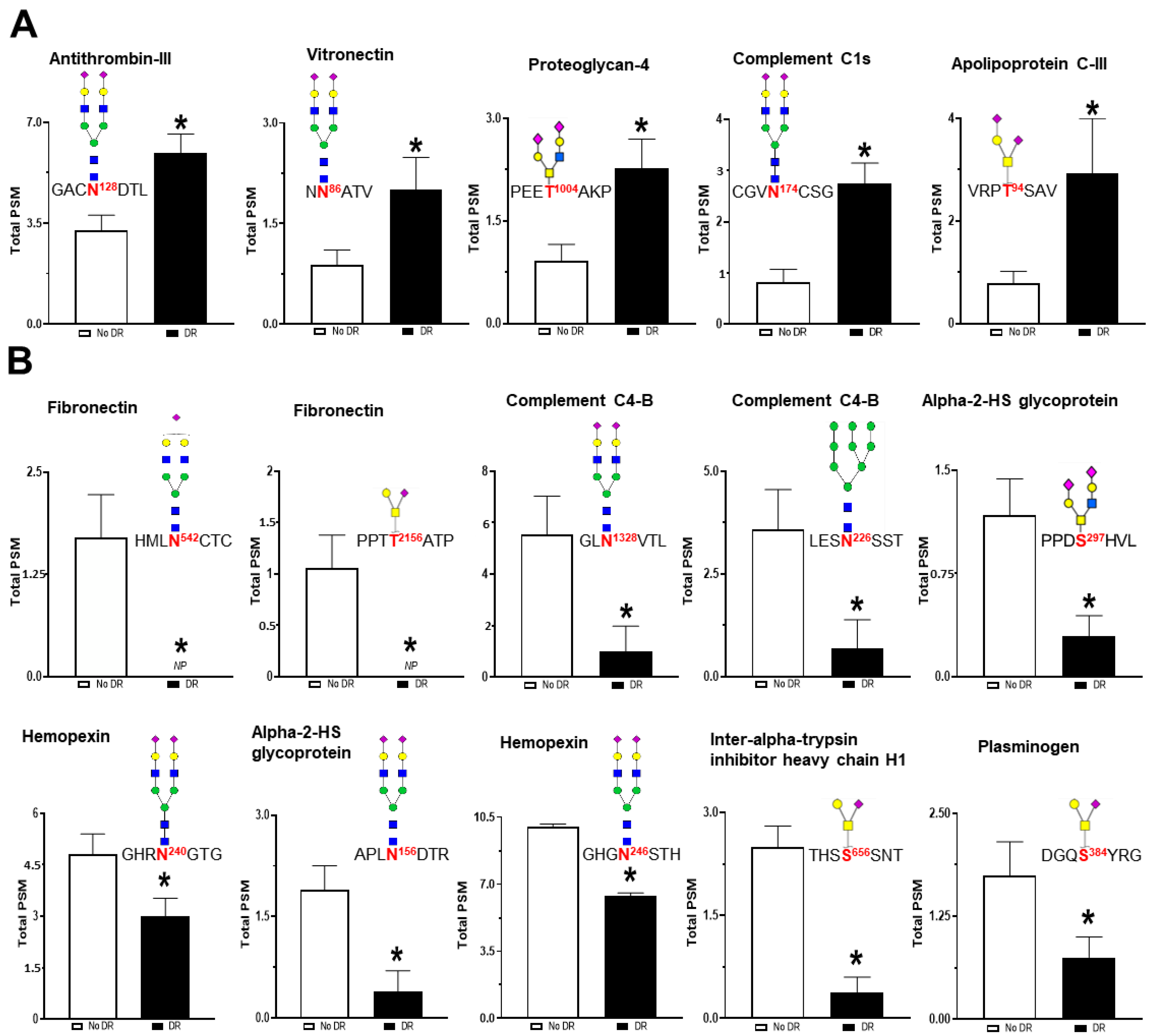
3.4. Serum Glycoproteomic Alterations Associated with Diabetic Retinopathy
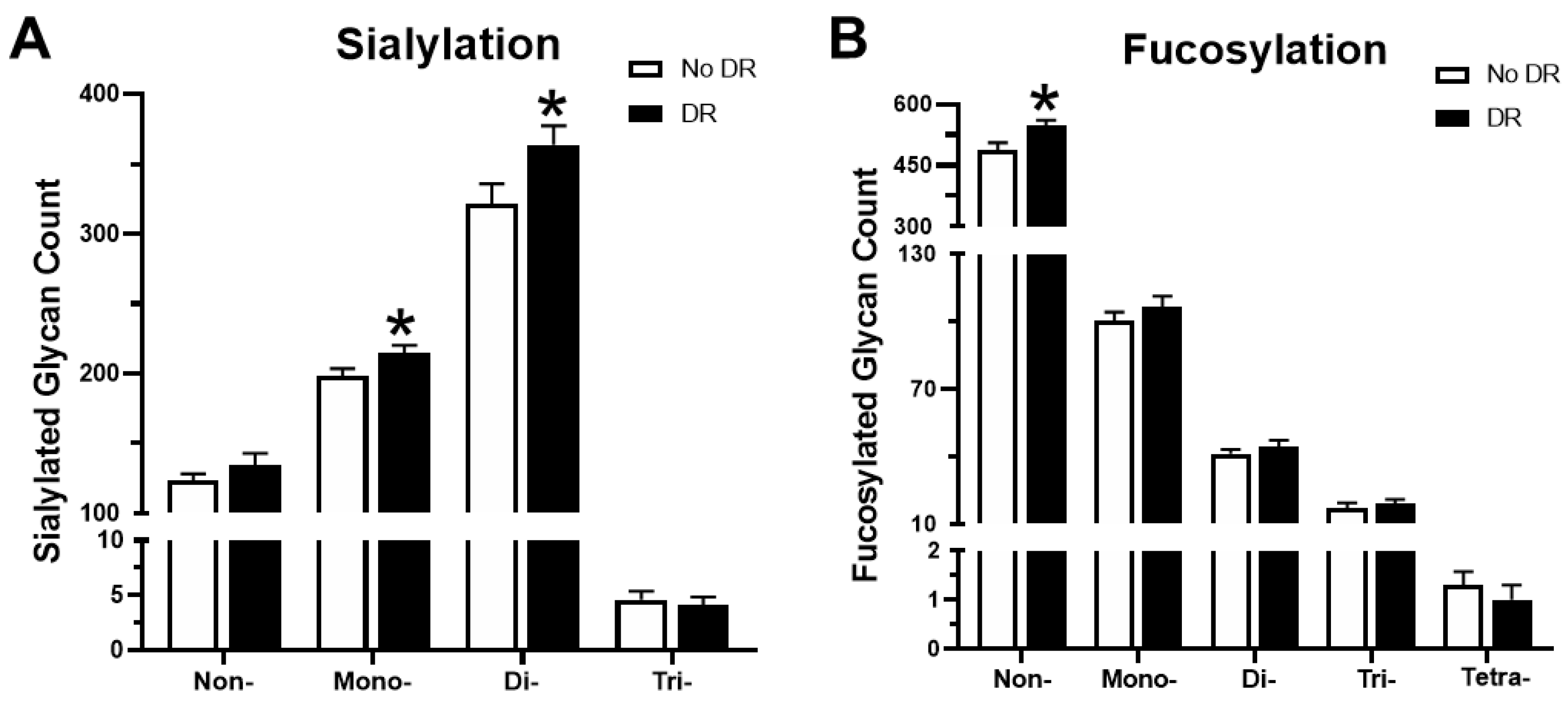
3.5. Trends in Sialylation and Fucosylation of Glycans
4. Discussion
5. Conclusions
Author Contributions
Funding
Conflicts of Interest
References
- Kempen, J.H.; O’Colmain, B.J.; Leske, M.C.; Haffner, S.M.; Klein, R.; Moss, S.E.; Taylor, H.R.; Hamman, R.F.; West, S.K.; Wang, J.J. The prevalence of diabetic retinopathy among adults in the United States. Arch. Ophthalmol. 2004, 122, 552–563. [Google Scholar]
- Guariguata, L.; Whiting, D.R.; Hambleton, I.; Beagley, J.; Linnenkamp, U.; Shaw, J.E. Global estimates of diabetes prevalence for 2013 and projections for 2035. Diabetes Res. Clin. Pract. 2014, 103, 137–149. [Google Scholar] [CrossRef] [PubMed]
- El-Asrar, A.M.A. Role of inflammation in the pathogenesis of diabetic retinopathy. Middle East Afr. J. Ophthalmol. 2012, 19, 70. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gardner, T.W.; Sundstrom, J.M. A proposal for early and personalized treatment of diabetic retinopathy based on clinical pathophysiology and molecular phenotyping. Vis. Res. 2017, 139, 153–160. [Google Scholar] [CrossRef] [PubMed]
- Ohtsubo, K.; Marth, J.D. Glycosylation in cellular mechanisms of health and disease. Cell 2006, 126, 855–867. [Google Scholar] [CrossRef] [Green Version]
- Saldova, R.; Wormald, M.R.; Dwek, R.A.; Rudd, P.M. Glycosylation changes on serum glycoproteins in ovarian cancer may contribute to disease pathogenesis. Dis. Markers 2008, 25, 219–232. [Google Scholar] [CrossRef] [PubMed]
- Li, X.; Ding, L.; Li, X.; Zhu, H.; Gashash, E.A.; Li, Z.; Wang, P.G.; Ma, C. An integrated proteomic and glycoproteomic study for differences on glycosylation occupancy in rheumatoid arthritis. Anal. Bioanal. Chem. 2019, 411, 1331–1338. [Google Scholar] [CrossRef]
- Shah, P.; Wang, X.; Yang, W.; Eshghi, S.T.; Sun, S.; Hoti, N.; Chen, L.; Yang, S.; Pasay, J.; Rubin, A. Integrated proteomic and glycoproteomic analyses of prostate cancer cells reveal glycoprotein alteration in protein abundance and glycosylation. Mol. Cell. Proteom. 2015, 14, 2753–2763. [Google Scholar] [CrossRef] [Green Version]
- Ang, I.L.; Poon, T.C.; Lai, P.B.; Chan, A.T.; Ngai, S.-M.; Hui, A.Y.; Johnson, P.J.; Sung, J.J. Study of serum haptoglobin and its glycoforms in the diagnosis of hepatocellular carcinoma: A glycoproteomic approach. J. Proteome Res. 2006, 5, 2691–2700. [Google Scholar] [CrossRef]
- Dube, D.H.; Bertozzi, C.R. Glycans in cancer and inflammation—Potential for therapeutics and diagnostics. Nat. Rev. Drug Discov. 2005, 4, 477–488. [Google Scholar] [CrossRef]
- Mariño, K.; Saldova, R.; Adamczyk, B.; Rudd, P.M.; Rauter, A. Changes in serum N-glycosylation profiles: Functional significance and potential for diagnostics. Carbohydr. Chem. 2012, 37, 57–93. [Google Scholar]
- Scott, D.W.; Patel, R.P. Endothelial heterogeneity and adhesion molecules N-glycosylation: Implications in leukocyte trafficking in inflammation. Glycobiology 2013, 23, 622–633. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Robinson, M.D.; McCarthy, D.J.; Smyth, G.K. edgeR: A Bioconductor package for differential expression analysis of digital gene expression data. Bioinformatics 2010, 26, 139–140. [Google Scholar] [CrossRef] [Green Version]
- Ritchie, M.E.; Phipson, B.; Wu, D.; Hu, Y.; Law, C.W.; Shi, W.; Smyth, G.K. limma powers differential expression analyses for RNA-sequencing and microarray studies. Nucleic Acids Res. 2015, 43, e47. [Google Scholar] [CrossRef] [PubMed]
- Cao, L.; Qu, Y.; Zhang, Z.; Wang, Z.; Prytkova, I.; Wu, S. Intact glycopeptide characterization using mass spectrometry. Expert. Rev. Proteom. 2016, 13, 513–522. [Google Scholar] [CrossRef] [Green Version]
- Chen, Z.; Huang, J.; Li, L. Recent advances in mass spectrometry (MS)-based glycoproteomics in complex biological samples. Trac. Trends. Anal. Chem. 2019, 118, 880–892. [Google Scholar] [CrossRef]
- Schumacher, K.N.; Dodds, E.D. A case for protein-level and site-level specificity in glycoproteomic studies of disease. Glycoconj. J. 2016, 33, 377–385. [Google Scholar] [CrossRef]
- Lemańska-Perek, A.; Adamik, B. Fibronectin and its soluble EDA-FN isoform as biomarkers for inflammation and sepsis. Adv. Clin. Exp. Med. 2019. [Google Scholar] [CrossRef] [Green Version]
- Miller, C.G.; Budoff, G.; Prenner, J.L.; Schwarzbauer, J.E. Minireview: Fibronectin in retinal disease. Exp. Biol. Med. 2017, 242, 1–7. [Google Scholar] [CrossRef] [Green Version]
- Kanters, S.D.; Banga, J.-D.; Algra, A.; Frijns, R.C.; Beutler, J.J.; Fijnheer, R. Plasma levels of cellular fibronectin in diabetes. Diabetes Care 2001, 24, 323–327. [Google Scholar] [CrossRef] [Green Version]
- Jones, G.E.; Arumugham, R.G.; Tanzer, M.L. Fibronectin glycosylation modulates fibroblast adhesion and spreading. J. Cell Biol. 1986, 103, 1663–1670. [Google Scholar] [CrossRef]
- Hsiao, C.-T.; Cheng, H.-W.; Huang, C.-M.; Li, H.-R.; Ou, M.-H.; Huang, J.-R.; Khoo, K.-H.; Yu, H.W.; Chen, Y.-Q.; Wang, Y.-K. Fibronectin in cell adhesion and migration via N-glycosylation. Oncotarget 2017, 8, 70653. [Google Scholar] [CrossRef] [Green Version]
- Samsom, M.L.; Morrison, S.; Masala, N.; Sullivan, B.D.; Sullivan, D.A.; Sheardown, H.; Schmidt, T.A. Characterization of full-length recombinant human Proteoglycan 4 as an ocular surface boundary lubricant. Exp. Eye Res. 2014, 127, 14–19. [Google Scholar] [CrossRef] [PubMed]
- Schmidt, T.A.; Sullivan, D.A.; Knop, E.; Richards, S.M.; Knop, N.; Liu, S.; Sahin, A.; Darabad, R.R.; Morrison, S.; Kam, W.R. Transcription, translation, and function of lubricin, a boundary lubricant, at the ocular surface. Jama Ophthalmol. 2013, 131, 766–776. [Google Scholar] [CrossRef]
- Wang, L.; Kikuchi, S.; Schmidt, T.A.; Hoofnagle, M.; Wight, T.N.; Azuma, N.; Tang, G.L.; Sobel, M.; Velamoor, G.R.; Mokadam, N.A. Inhibitory Effects of PRG4 on Migration and Proliferation of Human Venous Cells. J. Surg. Res. 2020, 253, 53–62. [Google Scholar] [CrossRef]
- Steele, B.; Alvarez-Veronesi, M.; Schmidt, T. Molecular weight characterization of PRG4 proteins using multi-angle laser light scattering (MALLS). Osteoarthr. Cartil. 2013, 21, 498–504. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mortensen, S.; Kidmose, R.T.; Petersen, S.V.; Szilágyi, Á.; Prohászka, Z.; Andersen, G.R. Structural Basis for the Function of Complement Component C4 within the Classical and Lectin Pathways of Complement. J. Immunol. 2015, 194, 5488–5496. [Google Scholar] [CrossRef] [Green Version]
- Garcia-Ramirez, M.; Canals, F.; Hernández, C.; Colome, N.; Ferrer, C.; Carrasco, E.; Garcia-Arumi, J.; Simo, R. Proteomic analysis of human vitreous fluid by fluorescence-based difference gel electrophoresis (DIGE): A new strategy for identifying potential candidates in the pathogenesis of proliferative diabetic retinopathy. Diabetologia 2007, 50, 1294–1303. [Google Scholar] [CrossRef] [Green Version]
- Gaboriaud, C.; Rossi, V.; Bally, I.; Arlaud, G.J.; Fontecilla-Camps, J.C. Crystal structure of the catalytic domain of human complement C1s: A serine protease with a handle. Embo J. 2000, 19, 1755–1765. [Google Scholar] [CrossRef] [Green Version]
- Østergaard, J.; Hansen, T.K.; Thiel, S.; Flyvbjerg, A. Complement activation and diabetic vascular complications. Clin. Chim. Acta 2005, 361, 10–19. [Google Scholar] [CrossRef]
- Lu, Z.; Wang, F.; Liang, M. SerpinC1/Antithrombin III in kidney-related diseases. Clin. Sci. 2017, 131, 823–831. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mori, K.; Emoto, M.; Inaba, M. Fetuin-A: A multifunctional protein. Recent Pat. Endocr. Metab. Immune Drug Discov. 2011, 5, 124–146. [Google Scholar] [CrossRef]
- Zhuo, L.; Hascall, V.C.; Kimata, K. Inter-α-trypsin inhibitor, a covalent protein-glycosaminoglycan-protein complex. J. Biol. Chem. 2004, 279, 38079–38082. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bakker, W.W.; Borghuis, T.; Harmsen, M.C.; Van den Berg, A.; Kema, I.; Niezen, K.E.; Kapojos, J.J. Protease activity of plasma hemopexin. Kidney Int. 2005, 68, 603–610. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Preissner, K.T.; de Boer, H.; Pannekoek, H.; de Groot, P.G. Thrombin regulation by physiological inhibitors: The role of vitronectin. In Seminars in Thrombosis and Hemostasis; Thieme Medical Publishers Inc.: Leipzig, Germany, 1996; pp. 165–172. [Google Scholar]
- Mcauley, A. Biomarkers in Diabetic Retinopathy: Genetic and Proteomic Profiling. Ph.D. Thesis, The University of Melbourne, Melbourne, Australia, 2014. [Google Scholar]
- Kim, T.H.; Koo, J.H.; Heo, M.J.; Han, C.Y.; Kim, Y.-I.; Park, S.-Y.; Cho, I.J.; Lee, C.H.; Choi, C.S.; Lee, J.W. Overproduction of inter-α-trypsin inhibitor heavy chain 1 after loss of Gα13 in liver exacerbates systemic insulin resistance in mice. Sci. Transl. Med. 2019, 11, eaan4735. [Google Scholar] [CrossRef] [PubMed]
- Zhou, Z.-W.; Ju, H.-X.; Sun, M.-Z.; Fu, Q.-P.; Chen, H.-M.; Ji, H.-B.; Ji, F.; Jiang, D.-M. Serum fetuin-A levels are independently correlated with vascular endothelial growth factor and C-reactive protein concentrations in type 2 diabetic patients with diabetic retinopathy. Clin. Chim. Acta 2016, 455, 113–117. [Google Scholar] [CrossRef] [PubMed]
- Levy, J.H.; Sniecinski, R.M.; Welsby, I.J.; Levi, M. Antithrombin: Anti-inflammatory properties and clinical applications. Thromb. Haemost. 2016, 115, 712–728. [Google Scholar]
- Ceriello, A.; Giugliano, D.; Russo, P.D.; Tirelli, A.; Passariello, N.; Sgambato, S. Metabolic control may alter antithrombin III activity but not its plasma concentration in diabetes: A possible role for nonenzymatic glycosylation. Diabetes Care 1986, 9, 32–35. [Google Scholar] [CrossRef] [PubMed]
- Mitsi, M.; Forsten-Williams, K.; Gopalakrishnan, M.; Nugent, M.A. A catalytic role of heparin within the extracellular matrix. J. Biol. Chem. 2008, 283, 34796–34807. [Google Scholar] [CrossRef] [Green Version]
- Chillakuri, C.R.; Jones, C.; Mardon, H.J. Heparin binding domain in vitronectin is required for oligomerization and thus enhances integrin mediated cell adhesion and spreading. Febs. Lett. 2010, 584, 3287–3291. [Google Scholar] [CrossRef] [Green Version]
- Preissner, K.; Kanse, S.; Hammes, H.-P. Integrin chatter and vascular function in diabetic retinopathy. Horm. Metab. Res. 1997, 29, 643–645. [Google Scholar] [CrossRef]
- Esser, P.; Bresgen, M.; Weller, M.; Heimann, K.; Wiedemann, P. The significance of vitronectin in proliferative diabetic retinopathy. Graefe’s Arch. Clin. Exp. Ophthalmol. 1994, 232, 477–481. [Google Scholar] [CrossRef]
- Taketani, S.; Immenschuh, S.; Go, S.; Sinclair, P.R.; Stockert, R.J.; Liem, H.H.; Eberhard, U.M. Hemopexin from four species inhibits the association of heme with cultured hepatoma cells or primary rat hepatocytes exhibiting a small number of species specific hemopexin receptors. Hepatology 1998, 27, 808–814. [Google Scholar] [CrossRef] [PubMed]
- Santos, I.C.; Silbiger, V.N.; Higuchi, D.A.; Gomes, M.A.; Barcelos, L.S.; Teixeira, M.M.; Lopes, M.T.P.; Cardoso, V.N.; Lima, M.P.; Araujo, R.C.; et al. Angiostatic activity of human plasminogen fragments is highly dependent on glycosylation. Cancer Sci. 2010, 101, 453–459. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Collen, D. The plasminogen (fibrinolytic) system. Thromb. Haemost. 1999, 82, 259–270. [Google Scholar] [CrossRef] [PubMed]
- Kawakami, A.; Aikawa, M.; Libby, P.; Alcaide, P.; Luscinskas, F.W.; Sacks, F.M. Apolipoprotein CIII in apolipoprotein B lipoproteins enhances the adhesion of human monocytic cells to endothelial cells. Circulation 2006, 113, 691–700. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Klein, R.L.; McHenry, M.B.; Lok, K.H.; Hunter, S.J.; Le, N.-A.; Jenkins, A.J.; Zheng, D.; Semler, A.; Page, G.; Brown, W.V. Apolipoprotein C-III protein concentrations and gene polymorphisms in Type 1 diabetes: Associations with microvascular disease complications in the DCCT/EDIC cohort. J. Diabetes Complicat. 2005, 19, 18–25. [Google Scholar] [CrossRef] [PubMed]
- Zhang, P.; Gao, J.; Pu, C.; Zhang, Y. Apolipoprotein status in type 2 diabetes mellitus and its complications. Mol. Med. Rep. 2017, 16, 9279–9286. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, Q.; Hu, J.; Hu, Y.; Ding, Y.; Zhu, J.; Zhuang, C. Relationship between serum apolipoproteins levels and retinopathy risk in subjects with type 2 diabetes mellitus. Acta. Diabetol. 2018, 55, 681–689. [Google Scholar] [CrossRef] [PubMed]
- Kawakami, A.; Aikawa, M.; Alcaide, P.; Luscinskas, F.W.; Libby, P.; Sacks, F.M. Apolipoprotein CIII induces expression of vascular cell adhesion molecule-1 (VCAM-1) in vascular endothelial cells and increases adhesion of monocytic cells. Am. Heart Assoc. 2006. [Google Scholar] [CrossRef]
- Kawakami, A.; Aikawa, M.; Nitta, N.; Yoshida, M.; Libby, P.; Sacks, F.M. Apolipoprotein CIII-induced THP-1 cell adhesion to endothelial cells involves pertussis toxin-sensitive G protein-and protein kinase Cα-mediated nuclear factor-κB activation. Arterioscler. Thromb. Vasc. Biol. 2007, 27, 219–225. [Google Scholar] [CrossRef] [PubMed]
- Koska, J.; Yassine, H.; Trenchevska, O.; Sinari, S.; Schwenke, D.C.; Yen, F.T.; Billheimer, D.; Nelson, R.W.; Nedelkov, D.; Reaven, P.D. Disialylated apolipoprotein C-III proteoform is associated with improved lipids in prediabetes and type 2 diabetes. J. Lipid Res. 2016, 57, 894–905. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Akbri, M.Z.A.; Sheikh, A.S.; Bhatti, M.S.; Hussnain, M.; Chaudhry, Z.A. Serum sialic acid level in diabetic retinopathy. J. Ayub Med. Coll. Abbottabad 2001, 13, 29–30. [Google Scholar] [PubMed]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Patient Characteristics | Patients Without DR | Patients With DR |
|---|---|---|
| Subjects | N = 13 | N = 10 |
| Gender (F/M) | 8/5 | 7/3 |
| Race (B/W) | 5/8 | 6/4 |
| Age (years) | 69.25 ± 11.17 | 71.04 ± 12.15 |
| Duration of Disease (years) | 8.53 ± 2.83 | 12.81 ± 3.96 |
| Stage of DR (NPDR/PDR) | - | 4/6 |
| Hypertension (Y/N) | 6/7 | 7/3 |
| Accession ID | Gene Symbol | Protein Name | Glycopeptide | Glycan Structure | Average PSM |
|---|---|---|---|---|---|
| P02790 | HPX | Hemopexin | SWPAVGN187CSSALRW | 1.1.0.1 | 17.27 |
| P02765 | AHSG | Alpha-2-HS-glycoprotein | TVVQPS346VGAAAGPVVPPCPGRI | 1.1.0.1 | 16.85 |
| P02790 | HPX | Hemopexin | K.ALPQPQN453VTSLLGCTH- | 4.5.0.2 | 15.60 |
| P02765 | AHSG | Alpha-2-HS-glycoprotein | K.VCQDCPLLAPLN156DTRV | 4.5.0.2 | 10.38 |
| P02790 | HPX | Hemopexin | R.GHGHRNGTGHGN246STHHGPEYMRC | 4.5.0.2 | 10.02 |
| P19827 | ITIH1 | Inter-alpha-trypsin inhibitor heavy chain H1 | R.TFVLSALQPSPT653HSSSNTQRL | 1.1.0.1 | 8.18 |
| P02765 | AHSG | Alpha-2-HS-glycoprotein | K.AALAAFNAQNN176GSNFQLEEISRA | 4.5.0.2 | 7.03 |
| P08603 | CFH | Complement factor H | K.IPCSQPPQIEHGTIN882SSRS | 4.5.0.2 | 6.55 |
| P02790 | HPX | Hemopexin | R.GHGHRN240GTGHGNSTHHGPEYMRC | 4.5.0.2 | 6.39 |
| P00450 | CP | Ceruloplasmin | AGLQAFFQVQECN358KS | 4.5.0.2 | 6.14 |
| P02790 | HPX | Hemopexin | SWPAVGN187CSSALRW | 4.5.1.2 | 5.75 |
| P00738 | HP | Haptoglobin | VVLHPN241YSQVDIGLIKL | 4.5.0.2 | 5.70 |
| Q14624 | ITIH4 | Inter-alpha-trypsin inhibitor heavy chain H4 | LAILPASAPPATSNPDPAVS709RV | 3.3.2.2 | 5.59 |
| P0C0L5 | C4B | Complement C4-B | GLN1328VTLSSTGRN | 4.5.0.2 | 5.55 |
| P00450 | CP | Ceruloplasmin | EN397LTAPGSDSAVFFEQGTTRI | 4.5.0.2 | 5.39 |
| P00734 | F2 | Prothrombin | YPHKPEIN143STTHPGADLQENFCRN | 4.5.0.2 | 5.35 |
| P19827 | ITIH1 | Inter-alpha-trypsin inhibitor heavy chain H1 | RTFVLSALQPS651PTHSSSNTQRL | 1.1.0.1 | 5.07 |
| P02765 | AHSG | Alpha-2-HS-glycoprotein | TVVQPS346VGAAAGPVVPPCPGRI | 1.1.0.2 | 4.99 |
| P02765 | AHSG | Alpha-2-HS-glycoprotein | KVCQDCPLLAPLN156DTRV | 4.5.0.2 | 4.81 |
| P05155 | SERPING1 | Plasma protease C1 inhibitor | VGQLQLSHN352LSLVILVPQNLKH | 4.5.0.2 | 4.57 |
| P00450 | CP | Ceruloplasmin | EHEGAIYPDN138TTDFQRA | 4.5.0.2 | 4.54 |
| P02749 | APOH | Beta-2-glycoprotein 1 | VYKPSAGN162NSLYRD | 4.5.0.2 | 4.54 |
| P05546 | SERPIND1 | Heparin cofactor 2 | N49LSMPLLPADFHKE | 4.5.0.2 | 4.49 |
| P02790 | HPX | Hemopexin | SWPAVGN187CSSALRW | 4.5.0.2 | 4.49 |
| P02790 | HPX | Hemopexin | ALPQPQN453VTSLLGCTH- | 4.5.0.1 | 4.47 |
| P01008 | SERPINC1 | Antithrombin-III | SLTFN187ETYQDISELVYGAKL | 4.5.0.2 | 4.21 |
| P00734 | F2 | Prothrombin | SRYPHKPEIN143STTHPGADLQENFCRN | 4.5.0.2 | 3.88 |
| P0C0L5 | C4B | Complement C4-B | FSDGLESN226SSTQFEVKK | 2.9.0.0 | 3.59 |
| P02790 | HPX | Hemopexin | CSDGWSFDATTLDDN64GTMLFFKG | 4.5.0.2 | 3.33 |
| P01008 | SERPINC1 | Antithrombin-III | LGACN128DTLQQLMEVFKF | 4.5.0.2 | 3.25 |
| P02790 | HPX | Hemopexin | ALPQPQN453VTSLLGCTH- | 4.5.0.2 | 3.17 |
| P02790 | HPX | Hemopexin | GHGHRNGTGHGN246STHHGPEYMRC | 4.5.0.2 | 3.15 |
| P00450 | CP | Ceruloplasmin | EHEGAIYPDN138TTDFQRA | 4.5.1.2 | 3.10 |
| P00450 | CP | Ceruloplasmin | ELHHLQEQN762VSNAFLDKG | 4.5.0.2 | 3.10 |
| P01019 | AGT | Angiotensinogen | VYIHPFHLVIHN47ESTCEQLAKA | 4.5.0.2 | 2.99 |
| P01042 | KNG1 | Kininogen-1 | FSVATQTCQITPAEGPVVT137AQYDCLGCVHPISTQSPDLEPILRH | 1.1.0.1 | 2.98 |
| P00747 | PLG | Plasminogen | GNVAVTVSGHTCQHWSAQTPHTHN308RT | 4.5.0.2 | 2.84 |
| P19827 | ITIH1 | Inter-alpha-trypsin inhibitor heavy chain H1 | TFVLSALQPSPTHSS656SNTQRL | 1.1.0.1 | 2.49 |
| P02656 | APOC3 | Apolipoprotein C-III | DKFSEFWDLDPEVRPT94SAVAA- | 1.1.0.1 | 2.46 |
| P00450 | CP | Ceruloplasmin | ELHHLQEQN762VSNAFLDKGEFYIGSKY | 4.5.0.2 | 2.44 |
| P04196 | HRG | Histidine-rich glycoprotein | R.VIDFN125CTTSSVSSALANTKD | 4.5.0.1 | 2.35 |
| P04114 | APOB | Apolipoprotein B-100 | FVEGSHN3411STVSLTTKN | 4.5.0.1 | 2.34 |
| P19823 | ITIH2 | Inter-alpha-trypsin inhibitor heavy chain H2 | GAFISN118FSMTVDGKT | 4.5.0.2 | 2.24 |
| P02656 | APOC3 | Apolipoprotein C-III | FSEFWDLDPEVRPT94SAVAA- | 1.1.0.1 | 2.22 |
| P02749 | APOH | Beta-2-glycoprotein 1 | LGN253WSAMPSCKA | 4.5.0.2 | 2.09 |
| P04114 | APOB | Apolipoprotein B-100 | FVEGSHN3411STVSLTTKN | 3.5.0.1 | 2.02 |
| P00748 | F12 | Coagulation factor XII | RN433HSCEPCQTLAVRS | 4.5.0.2 | 2.00 |
| Q5JWR5 | DOPEY1 | Protein dopey-1 | KALET86YEIIFKI | 1.1.0.1 | 1.92 |
| P02765 | AHSG | Alpha-2-HS-glycoprotein | TVVQPS346VGAAAGPVVPPCPGRI | 1.2.1.0 | 1.89 |
| Accession ID | Symbol | Protein Name | Glycopeptide(s) | Glycan | Glycan Structure | Glycan Mass | Fold Change DR | p-Value |
|---|---|---|---|---|---|---|---|---|
| Upregulated | ||||||||
| P02656 | APOC3 | Apolipoprotein C-III | FSEFWDLDPEVRPT94SAVAA. | HexNAc(1)Hex(1)NeuAc(2) |  | 947.323 | 3.210 | 0.0002 |
| P09871 | C1S | Complement C1s subcomponent | NCGVN174CSGDVFTALIGEIASPNYPKPYPENSR.C | HexNAc(4)Hex(5)NeuAc(2) |  | 2204.772 | 2.858 | 0.0007 |
| Q92954 | PRG4 | Proteoglycan 4 | TITTTEIMNKPEET1004AKPK.D | HexNAc(2)Hex(2)NeuAc(2) |  | 1312.455 | 2.210 | 0.0133 |
| P04004 | VTN | Vitronectin | NN86ATVHEQVGGPSLTSDLQAQSK.G | HexNAc(4)Hex(5)NeuAc(2) |  | 2204.772 | 2.104 | 0.0482 |
| P01008 | SERPINC1 | Antithrombin-III | LGACN128DTLQQLMEVFK.F | HexNAc(4)Hex(5)NeuAc(2) |  | 2204.772 | 1.789 | 0.0026 |
| Downregulated | ||||||||
| P02751 | FN1 | Fibronectin | RHEEGHMLN542CTCFGQGR.G | HexNAc(4)Hex(5)NeuAc(1) |  | 1913.677 | 0.069 | 7.43 × 10−6 |
| P02751 | FN1 | Fibronectin | TTPPTT2156ATPIR.H | HexNAc(1)Hex(1)NeuAc(1) |  | 656.228 | 0.107 | 0.0007 |
| P0C0L5 | C4B | Complement C4-B | GLN1328VTLSSTGR.N | HexNAc(4)Hex(5)NeuAc(2) |  | 2204.772 | 0.185 | 7.25 × 10−10 |
| P0C0L5 | C4B | Complement C4-B | FSDGLESN226SSTQFEVK.K | HexNAc(2)Hex(9) |  | 1864.634 | 0.207 | 1.53 × 10−6 |
| P02765 | AHSG | Alpha-2-HS-glycoprotein | LGGAEVAVTCTVFQTQPVTSQPQPEGANEAVPTPVVDPDAPPSPPLGAPGLPPAGSPPDS297HVLLAAPPGHQLHR.A | HexNAc(2)Hex(2)NeuAc(2) |  | 1312.455 | 0.328 | 0.0331 |
| P02765 | AHSG | Alpha-2-HS-glycoprotein | KVCQDCPLLAPLN156DTR.V | HexNAc(4)Hex(5)NeuAc(2) |  | 2204.772 | 0.641 | 0.0445 |
| P02790 | HPX | Hemopexin | GHGHRN240GTGHGNSTHHGPEYMR.C | HexNAc(4)Hex(5)NeuAc(2) |  | 2204.772 | 0.256 | 0.0021 |
| P02790 | HPX | Hemopexin | GHGHRNGTGHGN246STHHGPEYMR.C | HexNAc(4)Hex(5)NeuAc(2) |  | 2204.772 | 0.639 | 0.0046 |
| P19827 | ITIH1 | Inter-alpha-trypsin inhibitor heavy chain H1 | TFVLSALQPSPTHSS656SNTQR.L | HexNAc(1)Hex(1)NeuAc(1) |  | 656.228 | 0.193 | 5.83 × 10−5 |
| P00747 | PLG | Plasminogen | IPSCDSSPVSTEQLAPTAPPELTPVVQDCYHGDGQS384YR.G | HexNAc(1)Hex(1)NeuAc(1) |  | 656.228 | 0.468 | 0.0374 |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Sharma, A.; Cox, J.; Glass, J.; Lee, T.J.; Kodeboyina, S.K.; Zhi, W.; Ulrich, L.; Lukowski, Z.; Sharma, S. Serum Glycoproteomic Alterations in Patients with Diabetic Retinopathy. Proteomes 2020, 8, 25. https://0-doi-org.brum.beds.ac.uk/10.3390/proteomes8030025
Sharma A, Cox J, Glass J, Lee TJ, Kodeboyina SK, Zhi W, Ulrich L, Lukowski Z, Sharma S. Serum Glycoproteomic Alterations in Patients with Diabetic Retinopathy. Proteomes. 2020; 8(3):25. https://0-doi-org.brum.beds.ac.uk/10.3390/proteomes8030025
Chicago/Turabian StyleSharma, Ashok, James Cox, Joshua Glass, Tae Jin Lee, Sai Karthik Kodeboyina, Wenbo Zhi, Lane Ulrich, Zachary Lukowski, and Shruti Sharma. 2020. "Serum Glycoproteomic Alterations in Patients with Diabetic Retinopathy" Proteomes 8, no. 3: 25. https://0-doi-org.brum.beds.ac.uk/10.3390/proteomes8030025