
Novel Binding Partners for CCT and PhLP1 Suggest a Common Folding Mechanism for WD40 Proteins with a 7-Bladed Beta-Propeller Structure

{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
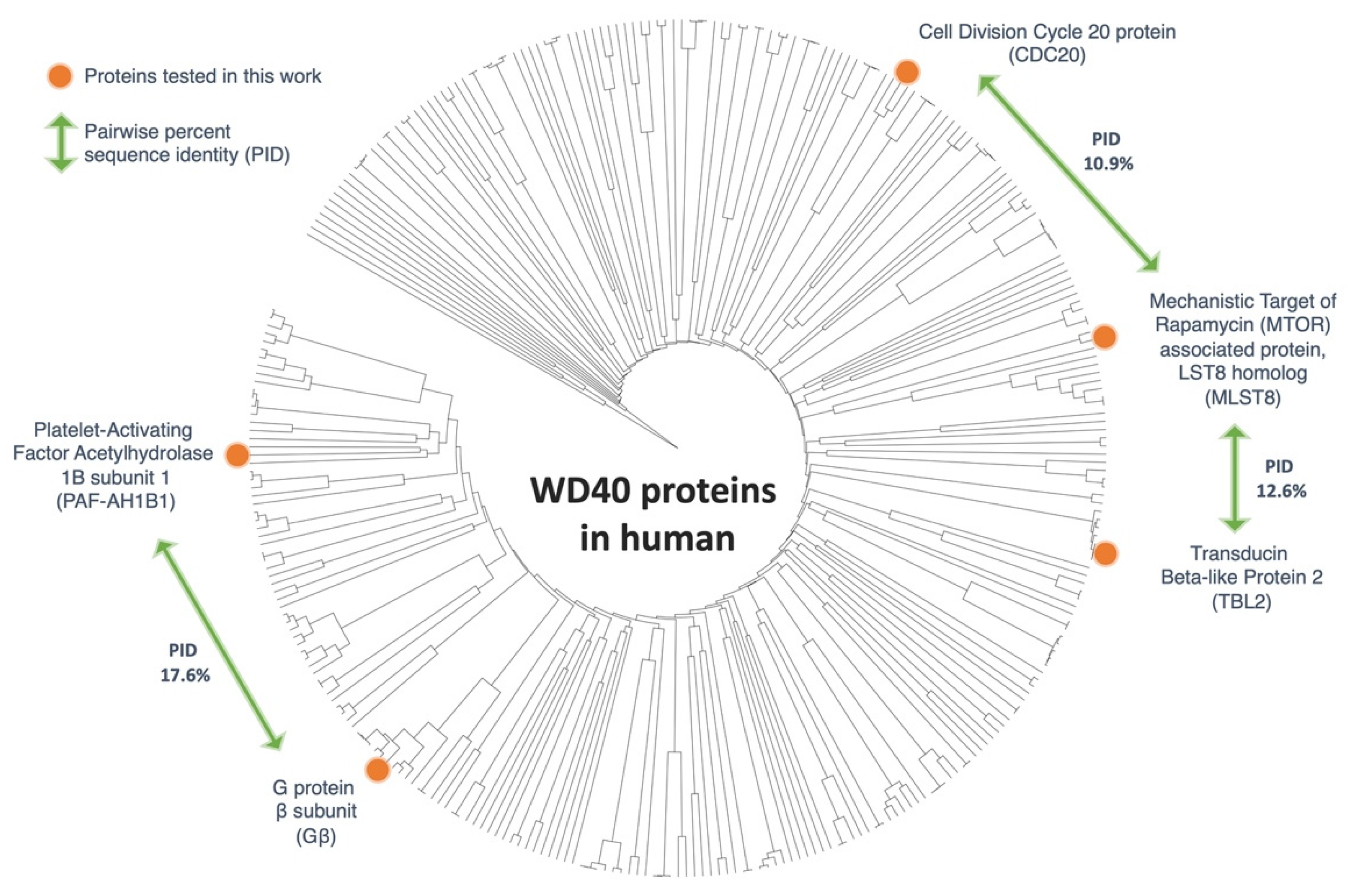
2.1. Phylogenetic Analysis
2.2. Preparation of Expression Constructs
2.3. Cell Culture
2.4. Transient Transfections
2.5. Immunoprecipitations (IP) and Western Immunoblotting
3. Results
3.1. Phylogenetic Analysis of WD40 Proteins
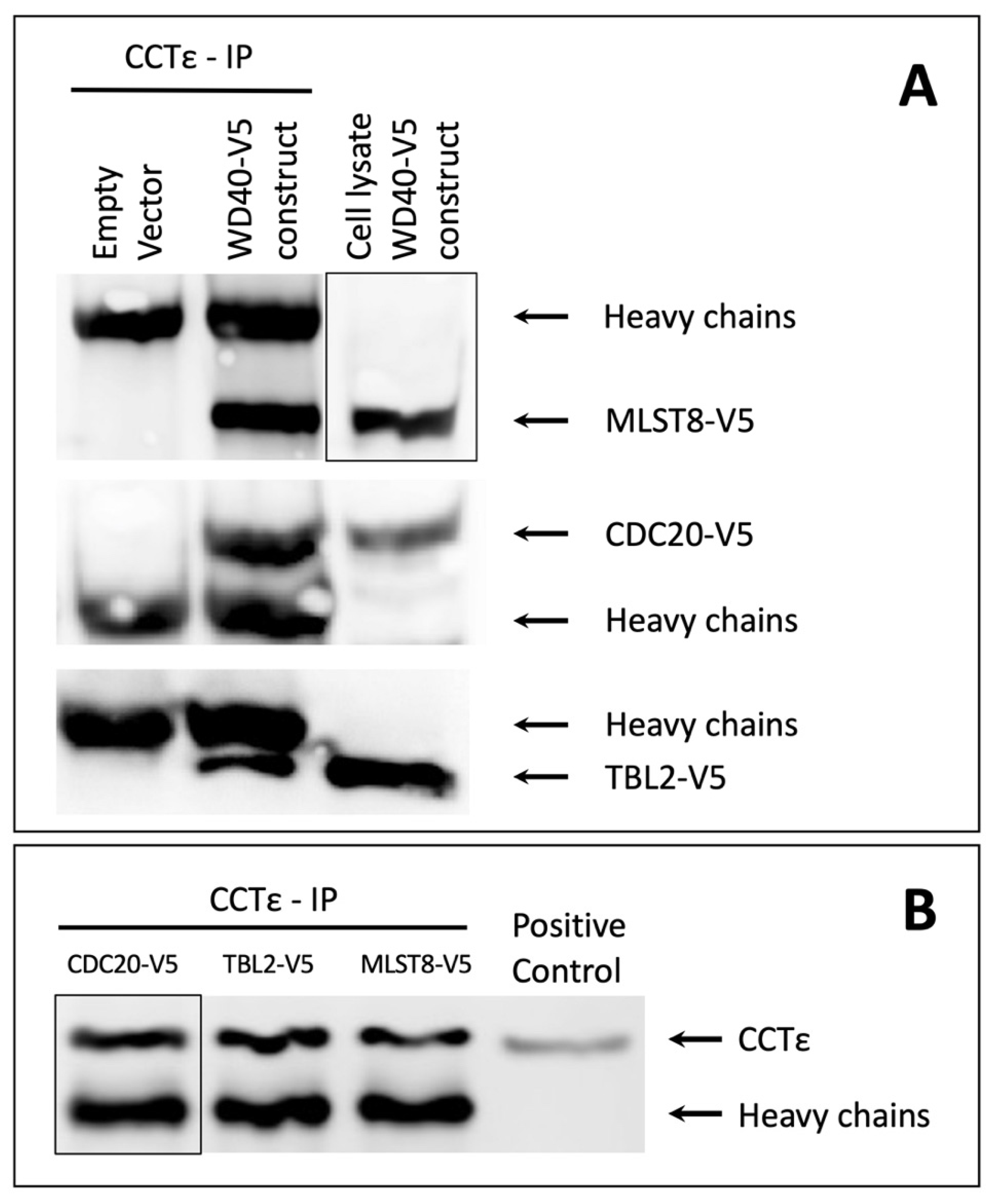
3.2. Binding of 7-Bladed β-Propeller WD40 Proteins to CCT
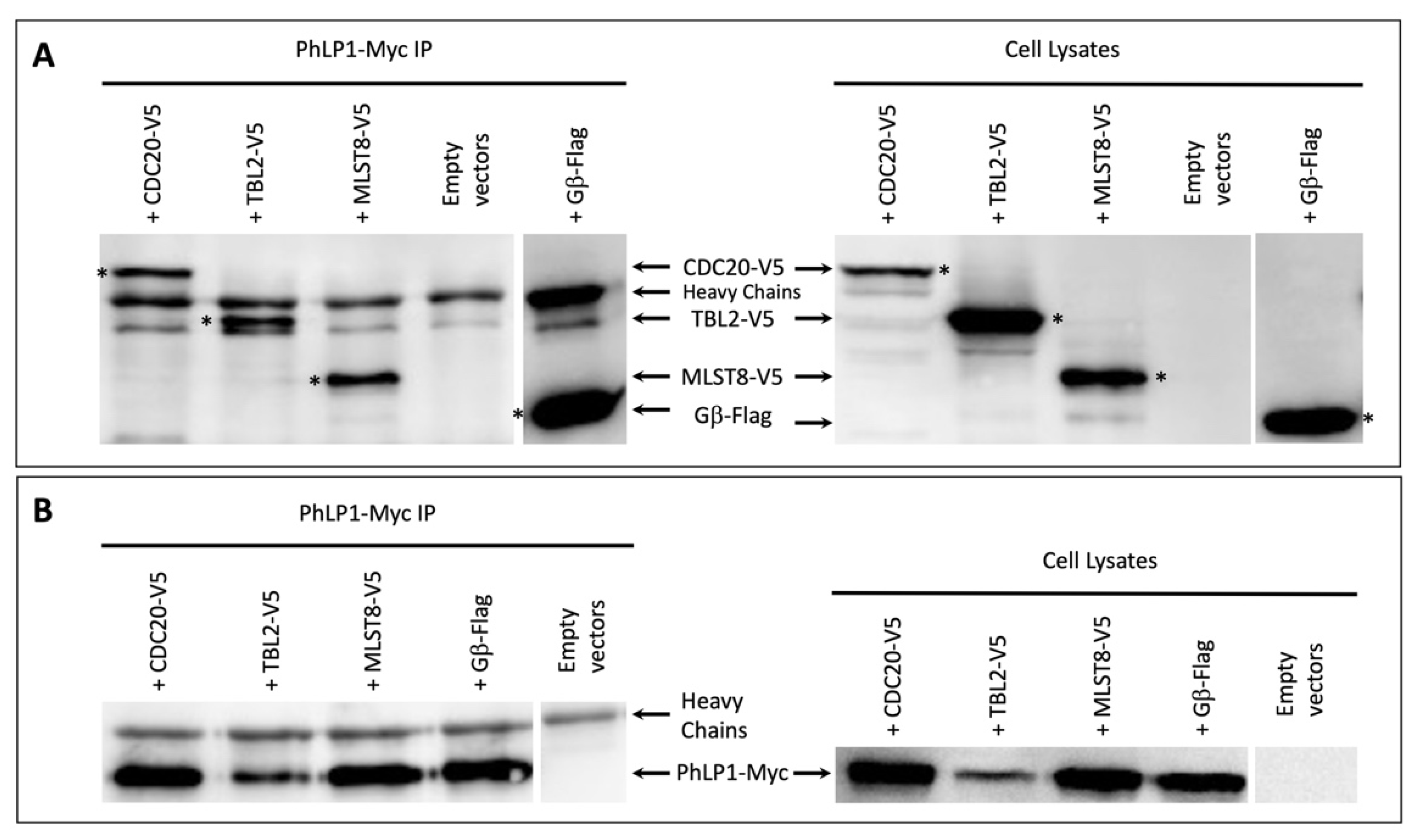
3.3. Binding of 7-Bladed β-Propeller WD40 Proteins to PhLP1
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Yam, A.Y.; Xia, Y.; Lin, H.-T.J.; Burlingame, A.; Gerstein, M.; Frydman, J. Defining the TRiC/CCT interactome links chaperonin function to stabilization of newly made proteins with complex topologies. Nat. Struct. Mol. Biol. 2008, 15, 1255–1262. [Google Scholar] [CrossRef] [PubMed]
- Willison, K.R. The substrate specificity of eukaryotic cytosolic chaperonin CCT. Philos. Trans. R. Soc. B Biol. Sci. 2018, 373, 20170192. [Google Scholar] [CrossRef] [PubMed]
- Llorca, O.; Smyth, M.G.; Carrascosa, J.L.; Willison, K.R.; Radermacher, M.; Steinbacher, S.; Valpuesta, J. 3D reconstruction of the ATP-bound form of CCT reveals the asymmetric folding conformation of a type II chaperonin. Nat. Genet. 1999, 6, 639–642. [Google Scholar] [CrossRef]
- Valpuesta, J.M.; Martín-Benito, J.; Gomez-Puertas, P.; Carrascosa, J.L.; Willison, K.R. Structure and function of a protein folding machine: The eukaryotic cytosolic chaperonin CCT. FEBS Lett. 2002, 529, 11–16. [Google Scholar] [CrossRef] [Green Version]
- Skjaerven, L.; Cuéllar, J.; Martinez, A.; Valpuesta, J. Dynamics, flexibility, and allostery in molecular chaperonins. FEBS Lett. 2015, 589, 2522–2532. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lopez, T.; Dalton, K.; Frydman, J. The Mechanism and Function of Group II Chaperonins. J. Mol. Biol. 2015, 427, 2919–2930. [Google Scholar] [CrossRef] [Green Version]
- Russmann, F.; Stemp, M.J.; Mönkemeyer, L.; Etchells, S.A.; Bracher, A.; Hartl, F.U. Folding of large multidomain proteins by partial encapsulation in the chaperonin TRiC/CCT. Proc. Natl. Acad. Sci. USA 2012, 109, 21208–21215. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ditzel, L.; Löwe, J.; Stock, D.; Stetter, K.O.; Huber, H.; Huber, R.; Steinbacher, S. Crystal Structure of the Thermosome, the Archaeal Chaperonin and Homolog of CCT. Cell 1998, 93, 125–138. [Google Scholar] [CrossRef] [Green Version]
- Willardson, B.M.; Tracy, C.M. Chaperone-Mediated Assembly of G Protein Complexes. GPCR Signal. Complexes–Synth. Assem. Traffick. Specif. 2012, 63, 131–153. [Google Scholar] [CrossRef]
- Arranz, R.; Martín-Benito, J.; Valpuesta, J.M. Structure and Function of the Cochaperone Prefoldin. Prefoldins New Chaperones 2018, 1106, 119–131. [Google Scholar] [CrossRef]
- Lukov, G.L.; Hu, T.; McLaughlin, J.N.; Hamm, H.E.; Willardson, B.M. Phosducin-like protein acts as a molecular chaperone for G protein betagamma dimer assembly. EMBO J. 2005, 24, 1965–1975. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Plimpton, R.L.; Cuellar, J.; Lai, C.W.; Aoba, T.; Makaju, A.; Franklin, S.; Mathis, A.D.; Prince, J.T.; Carrascosa, J.L.; Valpuesta, J.M.; et al. Structures of the Gbeta-CCT and PhLP1-Gbeta-CCT complexes reveal a mechanism for G-protein be-ta-subunit folding and Gbetagamma dimer assembly. Proc. Natl. Acad. Sci. USA 2015, 112, 2413–2418. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sondek, J.; Bohm, A.; Lambright, D.G.; Hamm, H.E.; Sigler, P.B. Crystal structure of a G-protein beta gamma dimer at 2.1A resolution. Nature 1996, 379, 369–374. [Google Scholar] [CrossRef] [PubMed]
- Lambright, D.G.; Sondek, J.; Bohm, A.; Skiba, N.P.; Hamm, H.E.; Sigler, P.B. The 2.0 A crystal structure of a heterotrimeric G protein. Nature 1996, 379, 311–319. [Google Scholar] [CrossRef]
- Stirnimann, C.; Petsalaki, E.; Russell, R.B.; Müller, C. WD40 proteins propel cellular networks. Trends Biochem. Sci. 2010, 35, 565–574. [Google Scholar] [CrossRef]
- Xu, C.; Min, J. Structure and function of WD40 domain proteins. Protein Cell 2011, 2, 202–214. [Google Scholar] [CrossRef]
- Schapira, M.; Tyers, M.; Torrent, M.; Arrowsmith, C.H. WD40 repeat domain proteins: A novel target class? Nat. Rev. Drug Discov. 2017, 16, 773–786. [Google Scholar] [CrossRef]
- Smith, T.F. Diversity of WD-Repeat proteins. Coronin Fam. Proteins 2008, 48, 20–30. [Google Scholar] [CrossRef]
- Dekker, C.; Stirling, P.C.; McCormack, E.A.; Filmore, H.; Paul, A.; Brost, R.L.; Costanzo, M.; Boone, C.; Leroux, M.R.; Willison, K.R. The interaction network of the chaperonin CCT. EMBO J. 2008, 27, 1827–1839. [Google Scholar] [CrossRef] [Green Version]
- Spiess, C.; Meyer, A.S.; Reissmann, S.; Frydman, J. Mechanism of the eukaryotic chaperonin: Protein folding in the chamber of secrets. Trends Cell Biol. 2004, 14, 598–604. [Google Scholar] [CrossRef] [Green Version]
- Willardson, B.M.; Howlett, A.C. Function of phosducin-like proteins in G protein signaling and chaperone-assisted protein folding. Cell. Signal. 2007, 19, 2417–2427. [Google Scholar] [CrossRef] [Green Version]
- Schroder, S.; Lohse, M.J. Quantification of the tissue levels and function of the G-protein regulator phosducin-like protein (PhlP). Naunyn Schmiedebergs Arch. Pharm. 2000, 362, 435–439. [Google Scholar] [CrossRef] [PubMed]
- Miles, M.F.; Barhite, S.; Sganga, M.; Elliott, M. Phosducin-like protein: An ethanol-responsive potential modulator of guanine nucleotide-binding protein function. Proc. Natl. Acad. Sci. USA 1993, 90, 10831–10835. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Willison, K.R. The structure and evolution of eukaryotic chaperonin-containing TCP-1 and its mechanism that folds actin into a protein spring. Biochem. J. 2018, 475, 3009–3034. [Google Scholar] [CrossRef] [PubMed]
- Finn, R.D.; Clements, J.; Arndt, W.; Miller, B.L.; Wheeler, T.J.; Schreiber, F.; Bateman, A.; Eddy, S. HMMER web server: 2015 update. Nucleic Acids Res. 2015, 43, W30–W38. [Google Scholar] [CrossRef]
- UniProt, C. UniProt: The universal protein knowledgebase in 2021. Nucleic Acids Res. 2021, 49, D480–D489. [Google Scholar]
- Fu, L.; Niu, B.; Zhu, Z.; Wu, S.; Li, W. CD-HIT: Accelerated for clustering the next-generation sequencing data. Bioinformatics 2012, 28, 3150–3152. [Google Scholar] [CrossRef] [PubMed]
- Kearse, M.; Moir, R.; Wilson, A.; Stones-Havas, S.; Cheung, M.; Sturrock, S.; Buxton, S.; Cooper, A.; Markowitz, S.; Duran, C.; et al. Geneious Basic: An integrated and extendable desktop software platform for the organization and analysis of sequence data. Bioinformatics 2012, 28, 1647–1649. [Google Scholar] [CrossRef]
- Edgar, R.C. Muscle: Multiple sequence alignment with high accuracy and high throughput. Nucleic Acids Res. 2004, 32, 1792–1797. [Google Scholar] [CrossRef] [Green Version]
- Letunic, I.; Bork, P. Interactive Tree Of Life (iTOL) v5: An online tool for phylogenetic tree display and annotation. Nucleic Acids Res. 2021, 49, W293–W296. [Google Scholar] [CrossRef]
- Ciccarelli, F.; Doerks, T.; von Mering, C.; Creevey, C.; Snel, B.; Bork, P. Toward Automatic Reconstruction of a Highly Resolved Tree of Life. Science 2006, 311, 1283–1287. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Carter, M.D.; Southwick, K.; Lukov, G.; Willardson, B.M.; Thulin, C.D. Identification of Phosphorylation Sites on Phosducin-like Protein by QTOF Mass Spectrometry. J. Biomol. Tech. JBT 2004, 15, 257–264. [Google Scholar] [PubMed]
- Camasses, A.; Bogdanova, A.; Shevchenko, A.; Zachariae, W. The CCT Chaperonin Promotes Activation of the Anaphase-Promoting Complex through the Generation of Functional Cdc20. Mol. Cell 2003, 12, 87–100. [Google Scholar] [CrossRef]
- Kaisari, S.; Sitry-Shevah, D.; Miniowitz-Shemtov, S.; Teichner, A.; Hershko, A. Role of CCT chaperonin in the disassembly of mitotic checkpoint complexes. Proc. Natl. Acad. Sci. USA 2017, 114, 956–961. [Google Scholar] [CrossRef] [Green Version]
- Huttlin, E.L.; Bruckner, R.J.; Paulo, J.A.; Cannon, J.R.; Ting, L.; Baltier, K.; Colby, G.; Gebreab, F.; Gygi, M.P.; Parzen, H.; et al. Architecture of the human interactome defines protein communities and disease networks. Nature 2017, 545, 505–509. [Google Scholar] [CrossRef]
- Kabir, M.A.; Kaminska, J.; Segel, G.B.; Bethlendy, G.; Lin, P.; della Seta, F.; Blegen, C.; Swiderek, K.M.; Żołądek, T.; Arndt, K.T.; et al. Physiological effects of unassembled chaperonin Cct subunits in the yeast Saccharomyces cerevisiae. Yeast 2005, 22, 219–239. [Google Scholar] [CrossRef]
- Cuéllar, J.; Ludlam, W.G.; Tensmeyer, N.C.; Aoba, T.; Dhavale, M.; Santiago, C.; Bueno-Carrasco, M.T.; Mann, M.J.; Plimpton, R.L.; Makaju, A.; et al. Structural and functional analysis of the role of the chaperonin CCT in mTOR complex assembly. Nat. Commun. 2019, 10, 1–14. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lukov, G.L.; Baker, C.M.; Ludtke, P.J.; Hu, T.; Carter, M.D.; Hackett, R.A.; Willardson, B. MMechanism of assembly of G protein betagamma subunits by protein kinase CK2-phosphorylated phosducin-like protein and the cytosolic chaperonin complex. J. Biol. Chem. 2006, 281, 22261–22274. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Howlett, A.C.; Gray, A.J.; Hunter, J.M.; Willardson, B.M. Role of molecular chaperones in G protein beta5/regulator of G protein signaling dimer assembly and G protein betagamma dimer specificity. J. Biol. Chem. 2009, 284, 16386–16399. [Google Scholar] [CrossRef] [Green Version]



Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Mak, W.S.; Tsang, T.M.; Chan, T.Y.; Lukov, G.L. Novel Binding Partners for CCT and PhLP1 Suggest a Common Folding Mechanism for WD40 Proteins with a 7-Bladed Beta-Propeller Structure. Proteomes 2021, 9, 40. https://0-doi-org.brum.beds.ac.uk/10.3390/proteomes9040040
Mak WS, Tsang TM, Chan TY, Lukov GL. Novel Binding Partners for CCT and PhLP1 Suggest a Common Folding Mechanism for WD40 Proteins with a 7-Bladed Beta-Propeller Structure. Proteomes. 2021; 9(4):40. https://0-doi-org.brum.beds.ac.uk/10.3390/proteomes9040040
Chicago/Turabian StyleMak, Wai Shun, Tsz Ming Tsang, Tsz Yin Chan, and Georgi L. Lukov. 2021. "Novel Binding Partners for CCT and PhLP1 Suggest a Common Folding Mechanism for WD40 Proteins with a 7-Bladed Beta-Propeller Structure" Proteomes 9, no. 4: 40. https://0-doi-org.brum.beds.ac.uk/10.3390/proteomes9040040