
Protocol Optimization of Proteomic Analysis of Korean Ginseng (Panax ginseng Meyer)

 , ,
, ,  , and
, and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Chemicals
2.2. Experimental Material
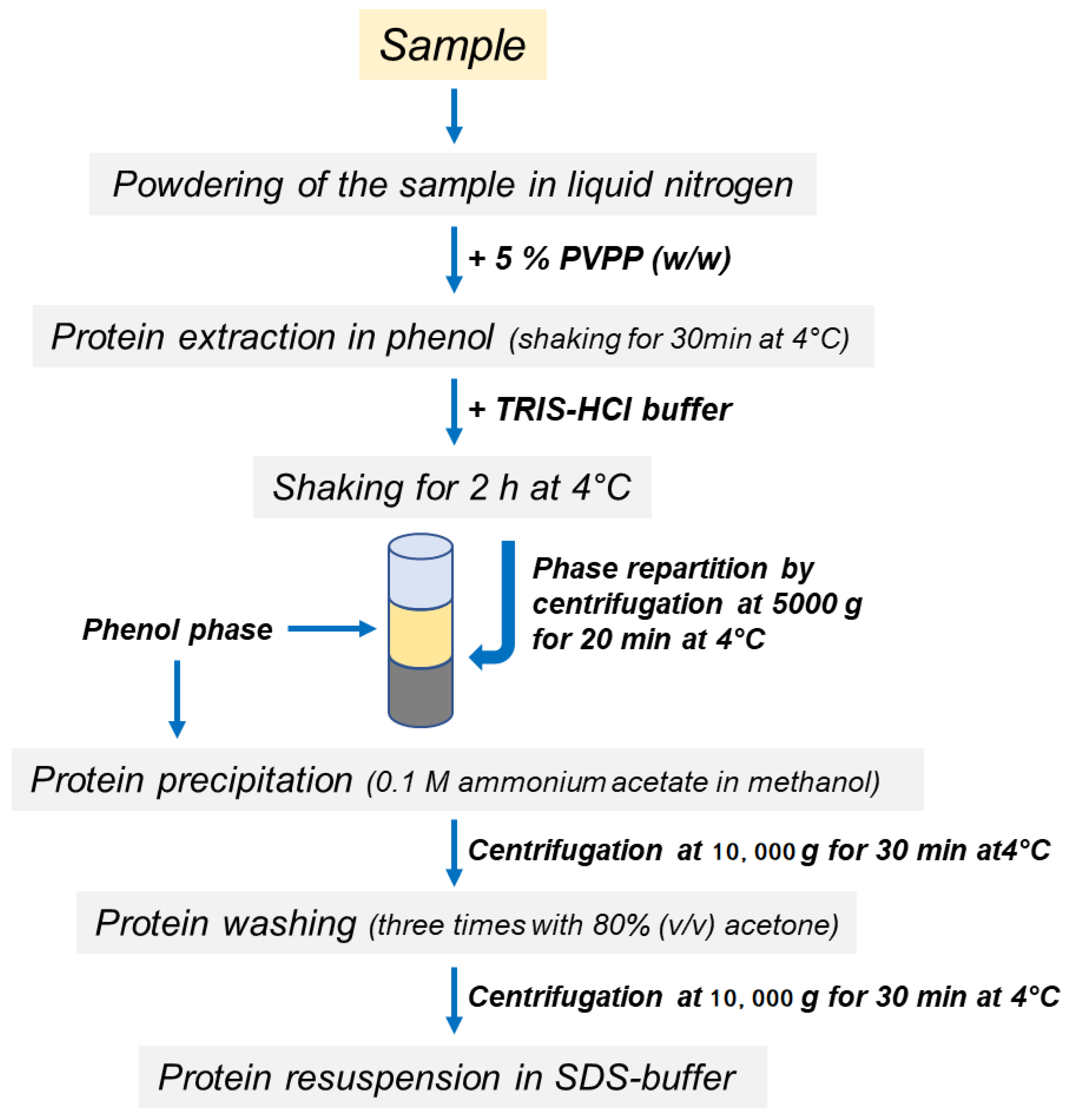
2.3. Protein Extraction
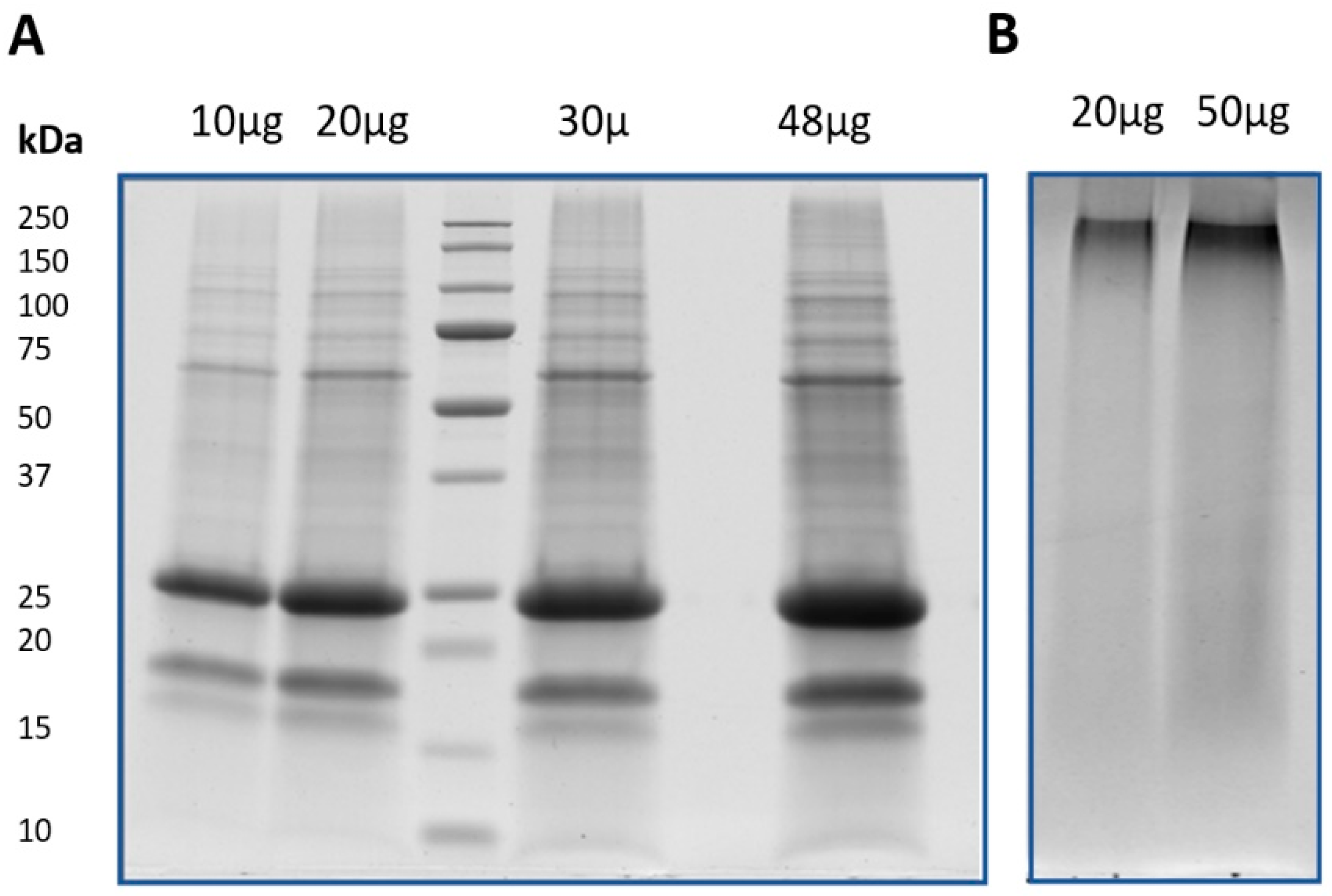
2.4. SDS-PAGE
2.5. Protein Digestion and Peptide Desalting
2.6. NLC-MS/MS Analysis
2.7. Protein Identification
2.8. MapMan Visualization
3. Results and Discussion
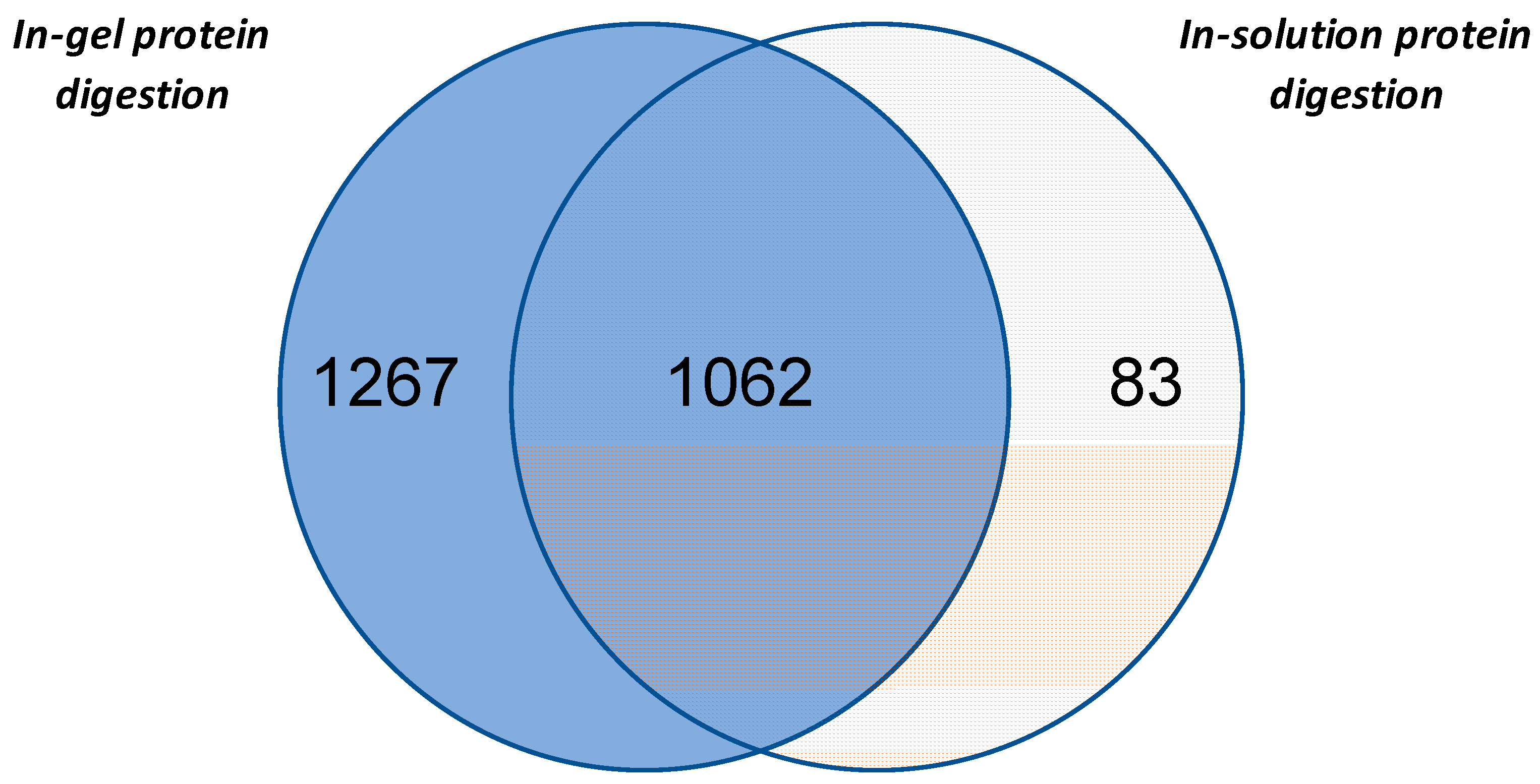
3.1. Evaluation of Different Proteomics Analysis Strategies
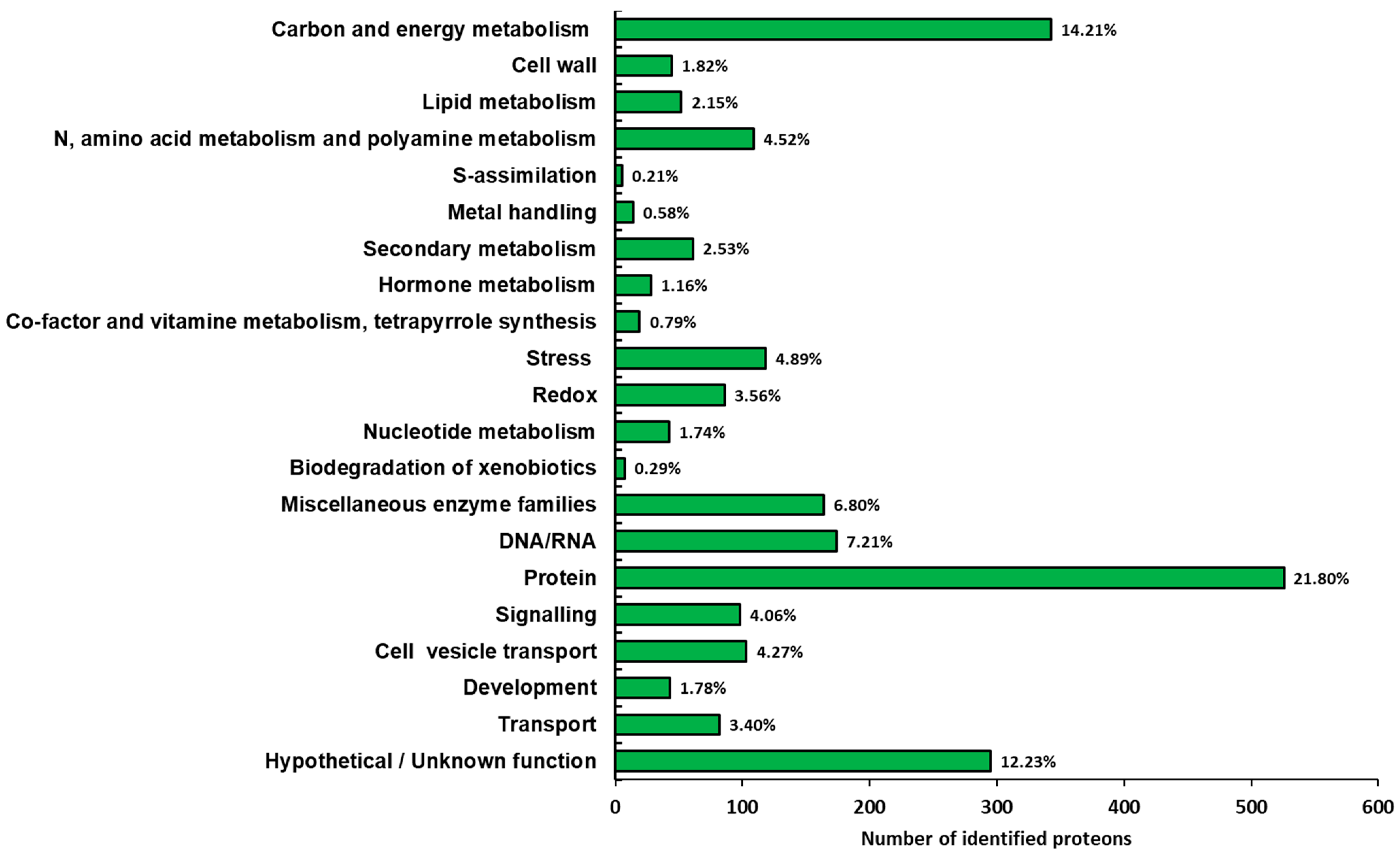
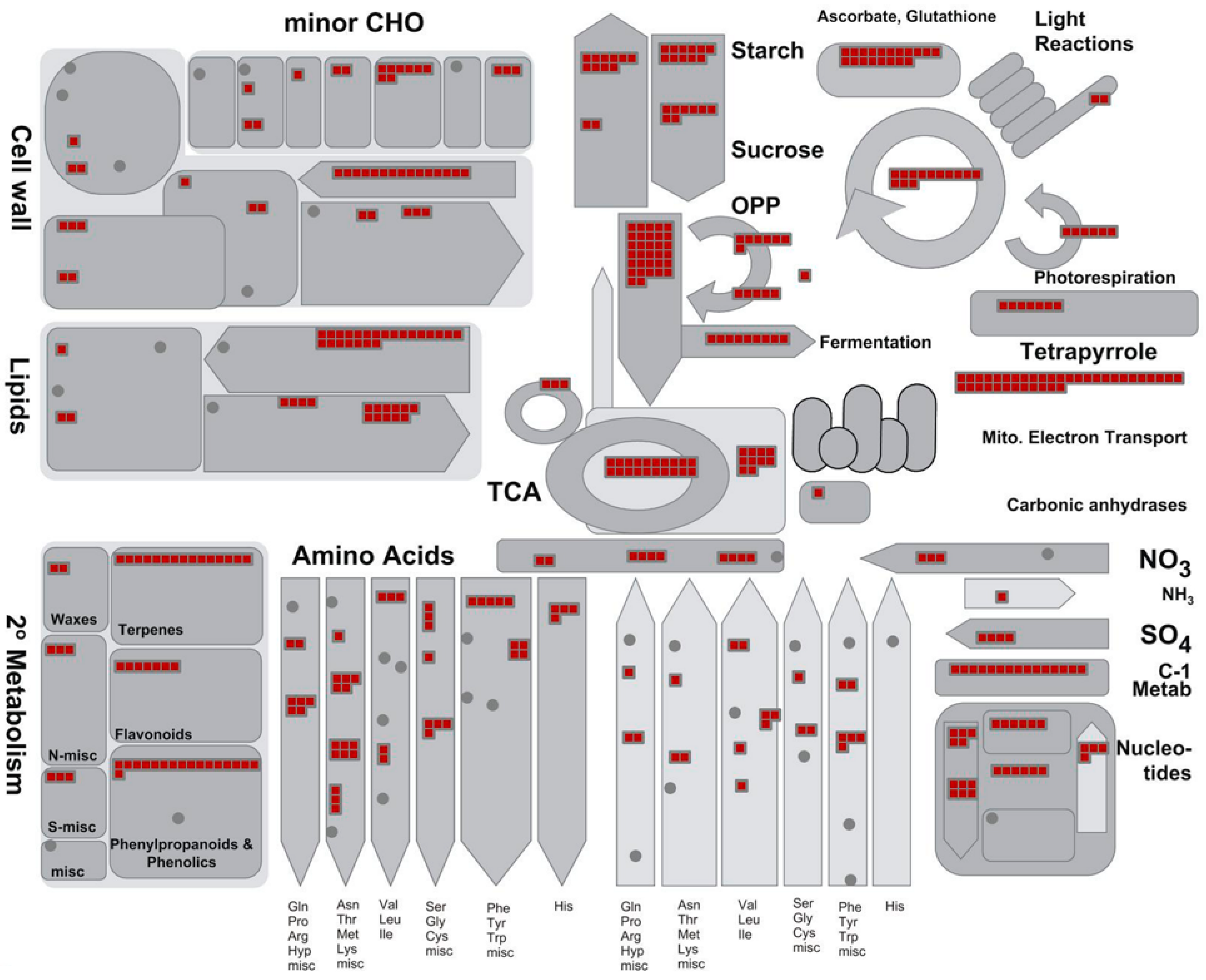
3.2. Functional Distribution of the Identified Proteins
3.3. Main Metabolic Pathways that Emerged from Proteomic Analysis
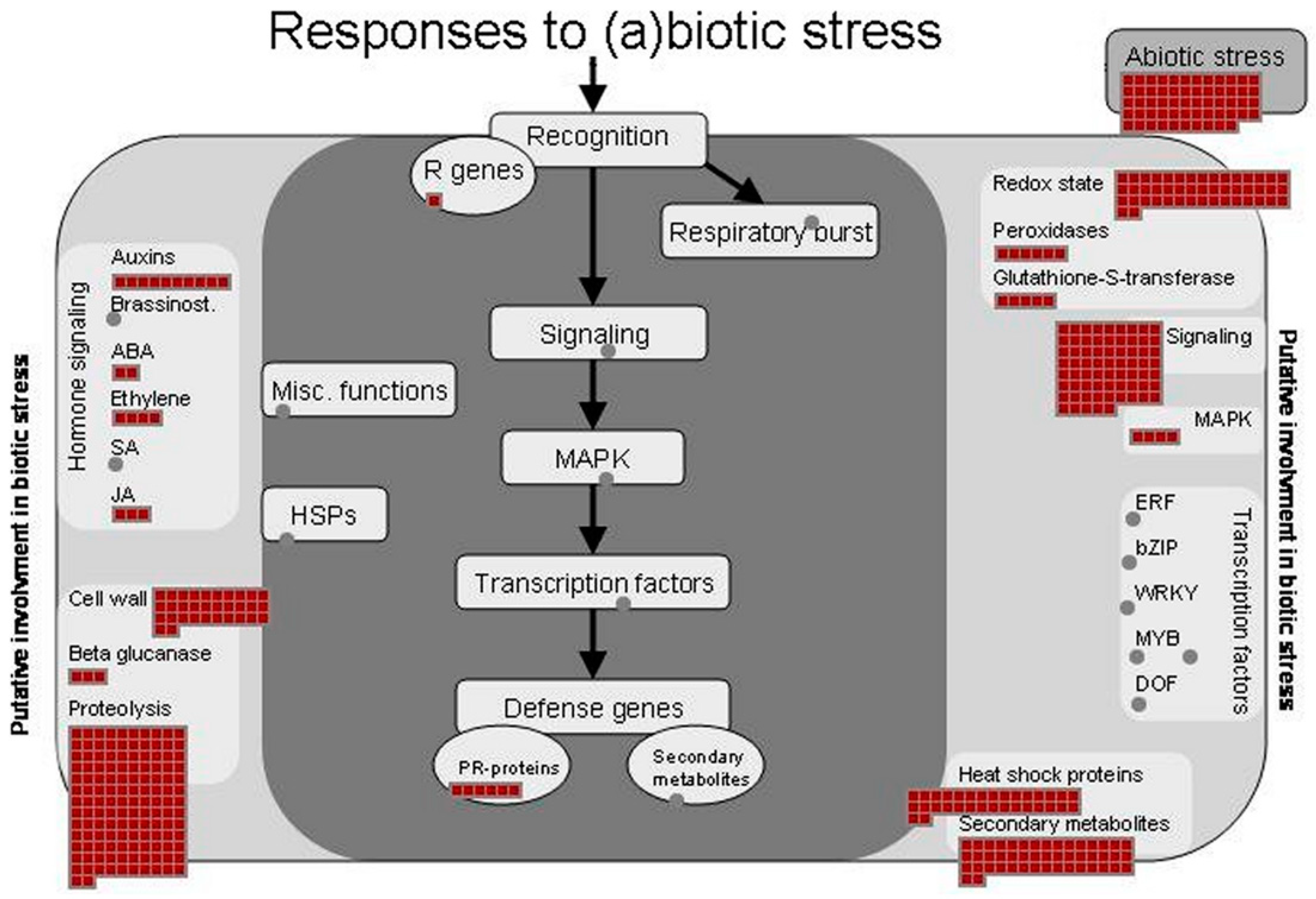
3.4. Identification of Proteins Involved in Stress Responses
3.5. Ginsenoside Biosynthetic Pathways
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Lee, F.C. Facts about Ginseng: The Elixir of Life; Hollym International Corporation: Elizabeth, NJ, USA, 1992; ISBN 10:0930878833. [Google Scholar]
- Petkov, W. Pharmacological Studies of the Drug P. Ginseng C.A. Meyer. Arzneim. Forsch. 1959, 9, 305–311. [Google Scholar]
- Colzani, M.; Altomare, A.; Caliendo, M.; Aldini, G.; Righetti, P.G.; Fasoli, E. The secrets of Oriental panacea: Panax ginseng. J. Proteom. 2016, 130, 150–159. [Google Scholar] [CrossRef]
- Attele, A.S.; Wu, J.A.; Yuan, C.-S. Ginseng pharmacology. Biochem. Pharmacol. 1999, 58, 1685–1693. [Google Scholar] [CrossRef]
- Kim, K.H.; Lee, D.; Lee, H.L.; Kim, C.-E.; Jung, K.; Kang, K.S. Beneficial effects of Panax ginseng for the treatment and prevention of neurodegenerative diseases: Past findings and future directions. J. Ginseng Res. 2018, 42, 239–247. [Google Scholar] [CrossRef]
- Wang, G.-L.; He, Z.-M.; Zhu, H.-Y.; Gao, Y.-G.; Zhao, Y.; Yang, H.; Zhang, L.-X. Involvement of serotonergic, noradrenergic and dopaminergic systems in the antidepressant-like effect of ginsenoside Rb1, a major active ingredient of Panax ginseng C.A. Meyer. J. Ethnopharmacol. 2017, 204, 118–124. [Google Scholar] [CrossRef]
- Lee, S.-Y.; Jeong, J.-J.; Eun, S.-H.; Kim, D.-H. Anti-inflammatory effects of ginsenoside Rg1 and its metabolites ginsenoside Rh1 and 20(S)-protopanaxatriol in mice with TNBS-induced colitis. Eur. J. Pharmacol. 2015, 762, 333–343. [Google Scholar] [CrossRef]
- Kang, J.-H.; Song, K.-H.; Woo, J.-K.; Park, M.H.; Rhee, M.H.; Choi, C.; Oh, S.H. Ginsenoside Rp1 from Panax ginseng Exhibits Anti-cancer Activity by Down-regulation of the IGF-1R/Akt Pathway in Breast Cancer Cells. Plant Foods Hum. Nutr. 2011, 66, 298–305. [Google Scholar] [CrossRef]
- Sharma, J.; Goyal, P.K. Chemoprevention of chemical-induced skin cancer by Panax ginseng root extract. J. Ginseng Res. 2015, 39, 265–273. [Google Scholar] [CrossRef] [Green Version]
- Lee, C.H.; Kim, J.-H. A review on the medicinal potentials of ginseng and ginsenosides on cardiovascular diseases. J. Ginseng Res. 2014, 38, 161–166. [Google Scholar] [CrossRef] [Green Version]
- Qi, H.; Huang, Y.; Yang, Y.; Dou, G.; Wan, F.; Zhang, W.; Yang, H.; Wang, L.; Wu, C.; Li, L. Anti-platelet activity of panaxatriol saponins is mediated by suppression of intracellular calcium mobilization and ERK2/p38 activation. BMC Complement. Altern. Med. 2016, 16, 1–8. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kim, K.; Kim, H.Y. Korean red ginseng stimulates insulin release from isolated rat pancreatic islets. J. Ethnopharmacol. 2008, 120, 190–195. [Google Scholar] [CrossRef]
- Jorrin, J.V.; Maldonado, A.M.; Castillejo, M.A. Plant proteome analysis: A 2006 update. Proteomics 2007, 7, 2947–2962. [Google Scholar] [CrossRef]
- Nam, M.H.; Kim, S.I.; Liu, J.R.; Yang, D.C.; Lim, Y.P.; Kwon, K.-H.; Yoo, J.S.; Park, Y.M. Proteomic analysis of Korean ginseng (Panax ginseng C.A. Meyer). J. Chromatogr. B 2005, 815, 147–155. [Google Scholar] [CrossRef] [PubMed]
- Kim, S.I.; Kweon, S.-M.; Kim, E.A.; Kim, J.Y.; Kim, S.; Yoo, J.S.; Park, Y.M. Characterization of RNase-like major storage protein from the ginseng root by proteomic approach. J. Plant Physiol. 2004, 161, 837–845. [Google Scholar] [CrossRef] [PubMed]
- Lum, J.H.-K.; Fung, K.-L.; Cheung, P.-Y.; Wong, M.-S.; Lee, C.-H.; Kwok, F.S.-L.; Leung, M.C.-P.; Hui, P.-K.; Lo, S.C.-L. Proteome of Oriental ginseng Panax ginseng C. A. Meyer and the potential to use it as an identification tool. Proteomics 2002, 2, 1123–1130. [Google Scholar] [CrossRef]
- Kim, S.I.; Kim, J.Y.; Kim, E.A.; Kwon, K.-H.; Kim, K.-W.; Cho, K.; Lee, J.H.; Nam, M.H.; Yang, D.-C.; Yoo, J.S.; et al. Proteome analysis of hairy root fromPanax ginseng C. A. Meyer using peptide fingerprinting, internal sequencing and expressed sequence tag data. Proteomics 2003, 3, 2379–2392. [Google Scholar] [CrossRef] [PubMed]
- Kim, N.-H.; Jayakodi, M.; Lee, S.-C.; Choi, B.-S.; Jang, W.; Lee, J.; Kim, H.H.; Waminal, N.E.; Lakshmanan, M.; Van Nguyen, B.; et al. Genome and evolution of the shade-requiring medicinal herbPanax ginseng. Plant Biotechnol. J. 2018, 16, 1904–1917. [Google Scholar] [CrossRef] [Green Version]
- Jayakodi, M.; Choi, B.-S.; Lee, S.-C.; Kim, N.-H.; Park, J.Y.; Jang, W.; Lakshmanan, M.; Mohan, S.V.G.; Lee, D.-Y.; Yang, T.-J. Ginseng Genome Database: An open-access platform for genomics of Panax ginseng. BMC Plant Biol. 2018, 18, 62. [Google Scholar] [CrossRef]
- Li, X.; Cheng, X.; Liao, B.; Xu, J.; Han, X.; Zhang, J.; Lin, Z.; Hu, L. Spatial protein expression of Panax ginseng by in-depth proteomic analysis for ginsenoside biosynthesis and transportation. J. Ginseng Res. 2021, 45, 58–65. [Google Scholar] [CrossRef]
- Lee, S.M.; Bae, B.-S.; Park, H.-W.; Ahn, N.-G.; Cho, B.-G.; Cho, Y.-L.; Kwak, Y.-S. Characterization of Korean Red Ginseng (Panax ginseng Meyer): History, preparation method, and chemical composition. J. Ginseng Res. 2015, 39, 384–391. [Google Scholar] [CrossRef] [Green Version]
- So, S.-H.; Lee, J.W.; Kim, Y.-S.; Hyun, S.H.; Han, C.-K. Red ginseng monograph. J. Ginseng Res. 2018, 42, 549–561. [Google Scholar] [CrossRef]
- Rose, J.K.; Bashir, S.; Giovannoni, J.J.; Jahn, M.M.; Saravanan, R.S. Tackling the plant proteome: Practical approaches, hurdles and experimental tools. Plant J. 2004, 39, 715–733. [Google Scholar] [CrossRef]
- Saravanan, R.S.; Rose, J.K.C. A critical evaluation of sample extraction techniques for enhanced proteomic analysis of recalcitrant plant tissues. Proteomics 2004, 4, 2522–2532. [Google Scholar] [CrossRef]
- Carpentier, S.C.; Witters, E.; Laukens, K.; Deckers, P.; Swennen, R.; Panis, B. Preparation of protein extracts from recalcitrant plant tissues: An evaluation of different methods for two-dimensional gel electrophoresis analysis. Proteomics 2005, 5, 2497–2507. [Google Scholar] [CrossRef] [PubMed]
- Prinsi, B.; Negri, A.S.; Failla, O.; Scienza, A.; Espen, L. Root proteomic and metabolic analyses reveal specific responses to drought stress in differently tolerant grapevine rootstocks. BMC Plant Biol. 2018, 18, 1–28. [Google Scholar] [CrossRef]
- Neilson, K.A.; George, I.S.; Emery, S.J.; Muralidharan, S.; Mirzaei, M.; Haynes, P.A. Analysis of Rice Proteins Using SDS-PAGE Shotgun Proteomics. In Advanced Structural Safety Studies; Springer: Berlin, Germany, 2013; Volume 1072, pp. 289–302. [Google Scholar]
- Vadivel, A.K.A. Gel-based proteomics in plants: Time to move on from the tradition. Front. Plant Sci. 2015, 6, 6. [Google Scholar] [CrossRef] [Green Version]
- Prinsi, B.; Negri, A.S.; Quattrocchio, F.M.; Koes, R.E.; Espen, L. Proteomics of red and white corolla limbs in petunia reveals a novel function of the anthocyanin regulator ANTHOCYANIN1 in determining flower longevity. J. Proteom. 2016, 131, 38–47. [Google Scholar] [CrossRef]
- Wilm, M.; Shevchenko, A.; Houthaeve, T.; Breit, S.; Schweigerer, L.; Fotsis, T.; Mann, M. Femtomole sequencing of proteins from polyacrylamide gels by nano-electrospray mass spectrometry. Nat. Cell Biol. 1996, 379, 466–469. [Google Scholar] [CrossRef]
- Thimm, O.; Bläsing, O.; Gibon, Y.; Nagel, A.; Meyer, S.; Krüger, P.; Selbig, J.; Müller, L.A.; Rhee, S.Y.; Stitt, M. mapman: A user-driven tool to display genomics data sets onto diagrams of metabolic pathways and other biological processes. Plant J. 2004, 37, 914–939. [Google Scholar] [CrossRef]
- Mine, Y.; Li-Chan, E.C.; Jiang, B. Biologically Active Food Proteins and Peptides in Health: An Overview. In Bioactive Proteins and Peptides as Functional Foods and Nutraceuticals; Wiley and Sons: Amsterdam, The Netherlands, 2010; pp. 3–11. [Google Scholar]
- Hurkman, W.J.; Tanaka, C.K. Solubilization of Plant Membrane Proteins for Analysis by Two-Dimensional Gel Electrophoresis. Plant Physiol. 1986, 81, 802–806. [Google Scholar] [CrossRef] [Green Version]
- Faurobert, M.; Pelpoir, E.; Chaïb, J.; Valerie, M.; Catherine, D.; Michel, Z.; Hervé, T. Phenol Extraction of Proteins for Proteomic Studies of Recalcitrant Plant Tissues. In Plant Proteomics; Springer: Berlin, Germany, 2007; Volume 355, pp. 9–14. [Google Scholar]
- Wu, Y.V.; Inglett, G.E. Denaturation of plant proteins related to functionality and food applications: A review. J. Food Sci. 1974, 39, 218–225. [Google Scholar] [CrossRef]
- Sánchez, A.; Vázquez, A. Bioactive peptides: A review. Food Qual. Saf. 2017, 1, 29–46. [Google Scholar] [CrossRef]
- Patil, S.P.; Goswami, A.; Kalia, K.; Kate, A.S. Plant-Derived Bioactive Peptides: A Treatment to Cure Diabetes. Int. J. Pept. Res. Ther. 2020, 26, 955–968. [Google Scholar] [CrossRef]
- Jayakodi, M.; Lee, S.-C.; Lee, Y.S.; Park, H.-S.; Kim, N.-H.; Jang, W.; Lee, H.O.; Joh, H.J.; Yang, T.-J. Comprehensive analysis of Panax ginseng root transcriptomes. BMC Plant Biol. 2015, 15, 138. [Google Scholar] [CrossRef] [Green Version]
- Kim, Y.-J.; Joo, S.C.; Shi, J.; Hu, C.; Quan, S.; Hu, J.; Sukweenadhi, J.; Mohanan, P.; Yang, D.-C.; Zhang, D. Metabolic dynamics and physiological adaptation of Panax ginseng during development. Plant Cell Rep. 2017, 37, 393–410. [Google Scholar] [CrossRef]
- Ruan, Y.-L. Sucrose Metabolism: Gateway to Diverse Carbon Use and Sugar Signaling. Annu. Rev. Plant Biol. 2014, 65, 33–67. [Google Scholar] [CrossRef]
- Ma, R.; Sun, L.; Chen, X.; Mei, B.; Chang, G.; Wang, M.; Zhao, D. Proteomic Analyses Provide Novel Insights into Plant Growth and Ginsenoside Biosynthesis in Forest Cultivated Panax ginseng (F. Ginseng). Front. Plant Sci. 2016, 7, 7. [Google Scholar] [CrossRef] [Green Version]
- Zhen, G.; Zhang, L.; Du, Y.; Yu, R.; Liu, X.; Cao, F.; Chang, Q.; Deng, X.; Xia, M.; He, H. De novo assembly and comparative analysis of root transcriptomes from different varieties of Panax ginseng C. A. Meyer grown in different environments. Sci. China Life Sci. 2015, 58, 1099–1110. [Google Scholar] [CrossRef] [Green Version]
- Demidchik, V. Mechanisms of oxidative stress in plants: From classical chemistry to cell biology. Environ. Exp. Bot. 2015, 109, 212–228. [Google Scholar] [CrossRef]
- Al-Whaibi, M.H. Plant heat-shock proteins: A mini review. J. King Saud Univ. Sci. 2011, 23, 139–150. [Google Scholar] [CrossRef] [Green Version]
- Haq, S.U.; Khan, A.; Ali, M.; Khattak, A.M.; Gai, W.-X.; Zhang, H.-X.; Wei, A.-M.; Gong, Z.-H. Heat Shock Proteins: Dynamic Biomolecules to Counter Plant Biotic and Abiotic Stresses. Int. J. Mol. Sci. 2019, 20, 5321. [Google Scholar] [CrossRef] [Green Version]
- Jung, J.H.; Kim, H.-Y.; Kim, H.S.; Jung, S.H. Transcriptome analysis of Panax ginseng response to high light stress. J. Ginseng Res. 2020, 44, 312–320. [Google Scholar] [CrossRef] [PubMed]
- Musidlak, O.; Bałdysz, S.; Krakowiak, M.; Nawrot, R. Plant latex proteins and their functions. In Advances in Botanical Research; Elsevier BV: Amsterdam, The Netherlands, 2020; Volume 93, pp. 55–97. [Google Scholar]
- Yu, Z.; Wang, X.; Zhang, L. Structural and Functional Dynamics of Dehydrins: A Plant Protector Protein under Abiotic Stress. Int. J. Mol. Sci. 2018, 19, 3420. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chen, J.-Y.; Dai, X.-F. Cloning and characterization of the Gossypium hirsutum major latex protein gene and functional analysis in Arabidopsis thaliana. Planta 2010, 231, 861–873. [Google Scholar] [CrossRef] [PubMed]
- Dunwell, J.M.; Gibbings, J.G.; Mahmood, T.; Naqvi, S.M.S. Germin and Germin-like Proteins: Evolution, Structure, and Function. Crit. Rev. Plant Sci. 2008, 27, 342–375. [Google Scholar] [CrossRef]
- Lee, Y.S.; Park, H.-S.; Lee, D.-K.; Jayakodi, M.; Kim, N.-H.; Koo, H.J.; Lee, S.-C.; Kim, Y.J.; Kwon, S.W.; Yang, T.-J. Integrated Transcriptomic and Metabolomic Analysis of Five Panax ginseng Cultivars Reveals the Dynamics of Ginsenoside Biosynthesis. Front. Plant Sci. 2017, 8, 8. [Google Scholar] [CrossRef] [PubMed] [Green Version]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Enxyme | Accession | Pathway 1 |
|---|---|---|
| acetyl-CoA C-acetyltransferase [EC:2.3.1.9] | Pg_S6240.3 | MVA |
| acetyl-CoA C-acetyltransferase [EC:2.3.1.9] | Pg_S0022.46 | |
| phosphomevalonate kinase [EC:2.7.4.2] | Pg_S3321.6 | |
| phosphomevalonate kinase [EC:2.7.4.2] | Pg_S5751.1 | |
| diphosphomevalonate decarboxylase [EC:4.1.1.33] | Pg_S1430.1 | |
| diphosphomevalonate decarboxylase [EC:4.1.1.33] | Pg_S3074.4 | |
| isopentenyl-diphosphate Delta-isomerase [EC:5.3.3.2] | Pg_S1168.2 | |
| 1-deoxy-D-xylulose-5-phosphate reductoisomerase [EC:1.1.1.267] | Pg_S0166.7 | MEP |
| 4-diphosphocytidyl-2-C-methyl-D-erythritol kinase [EC:2.7.1.148] | Pg_S0550.8 | |
| farnesyl diphosphate synthase [EC:2.5.1.1 2.5.1.10] | Pg_S0304.36 | Downstream pathway |
| farnesyl diphosphate synthase [EC:2.5.1.1 2.5.1.10] | Pg_S8325.1 | |
| Panax ginseng cytochrome P450 CYP716A53v2 | Pg_S1770.12 | |
| UDP-glycosyltransferases | Pg_S6708.3 | UGT |
| UDP-glycosyltransferases | Pg_S4157.4 | |
| UDP-glycosyltransferases | Pg_S2351.7 | |
| UDP-glycosyltransferases | Pg_S4493.1 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Braccia, C.; Prinsi, B.; Colzani, M.; Altomare, A.A.; Espen, L.; Lee, Y.-M.; Aldini, G.; Yeum, K.-J. Protocol Optimization of Proteomic Analysis of Korean Ginseng (Panax ginseng Meyer). Separations 2021, 8, 53. https://0-doi-org.brum.beds.ac.uk/10.3390/separations8040053
Braccia C, Prinsi B, Colzani M, Altomare AA, Espen L, Lee Y-M, Aldini G, Yeum K-J. Protocol Optimization of Proteomic Analysis of Korean Ginseng (Panax ginseng Meyer). Separations. 2021; 8(4):53. https://0-doi-org.brum.beds.ac.uk/10.3390/separations8040053
Chicago/Turabian StyleBraccia, Clarissa, Bhakti Prinsi, Mara Colzani, Alessandra A. Altomare, Luca Espen, Yoon-Mi Lee, Giancarlo Aldini, and Kyung-Jin Yeum. 2021. "Protocol Optimization of Proteomic Analysis of Korean Ginseng (Panax ginseng Meyer)" Separations 8, no. 4: 53. https://0-doi-org.brum.beds.ac.uk/10.3390/separations8040053