
Analysis of Differentially Expressed Proteins and Modifications Induced by Formaldehyde Using LC-MS/MS

 ,
, 
{kind=link}
{kind=link}
{kind=link}
{kind=link}
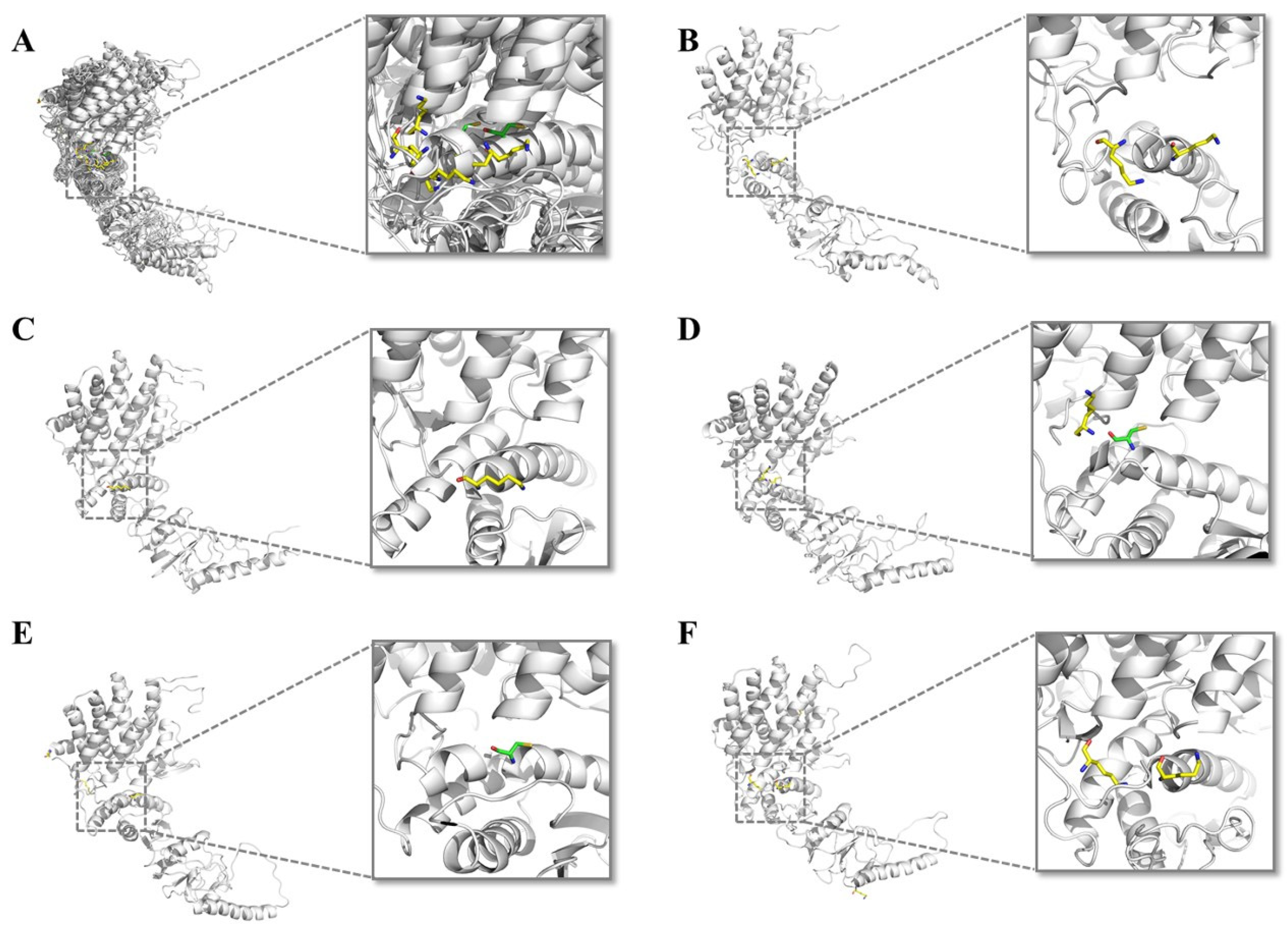{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Chemicals and Reagents
2.2. Cell Culture, Protein Extraction and Digestion
2.3. LC-MS/MS Analysis
2.4. Database Search
2.5. Bioinformatic Analysis
3. Results and Discussion
3.1. The Design of Analysis Method
3.2. Evaluation of Analysis Data Quality
3.3. Analysis of Proteins Differentially Induced by FA
3.4. The Analysis of Protein Modifications Induced by FA
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Bernardini, L.; Barbosa, E.; Charao, M.F.; Brucker, N. Formaldehyde toxicity reports fromin vitro and in vivo studies: A review and updated data. Drug Chem. Toxicol. 2020, 45, 972–984. [Google Scholar] [CrossRef] [PubMed]
- Dingler, F.A.; Wang, M.; Mu, A.; Millington, C.L.; Oberbeck, N.; Watcham, S.; Pontel, L.B.; Kamimae-Lanning, A.N.; Langevin, F.; Nadler, C.; et al. Two Aldehyde Clearance Systems Are Essential to Prevent Lethal Formaldehyde Accumulation in Mice and Humans. Mol. Cell 2020, 80, 996–1012.e9. [Google Scholar] [CrossRef] [PubMed]
- Wang, F.; Chen, D.; Wu, P.; Klein, C.; Jin, C. Formaldehyde, Epigenetics, and Alzheimer’s Disease. Chem. Res. Toxicol. 2019, 32, 820–830. [Google Scholar] [CrossRef] [PubMed]
- Szende, B.; Tyihak, E. Effect of formaldehyde on cell proliferation and death. Cell Biol. Int. 2010, 34, 1273–1282. [Google Scholar] [CrossRef]
- Marsh, G.M.; Youk, A.O.; Buchanich, J.M.; Erdal, S.; Esmen, N.A. Work in the metal industry and nasopharyngeal cancer mortality among formaldehyde-exposed workers. Regul. Toxicol. Pharmacol. 2007, 48, 308–319. [Google Scholar] [CrossRef]
- Nishikawa, A.; Nagano, K.; Kojima, H.; Ogawa, K. A comprehensive review of mechanistic insights into formaldehyde-induced nasal cavity carcinogenicity. Regul. Toxicol. Pharmacol. 2021, 123, 104937. [Google Scholar] [CrossRef]
- Bachand, A.M.; Mundt, K.A.; Mundt, D.J.; Montgomery, R.R. Epidemiological studies of formaldehyde exposure and risk of leukemia and nasopharyngeal cancer: A meta-analysis. Crit. Rev. Toxicol. 2010, 40, 85–100. [Google Scholar] [CrossRef]
- Checkoway, H.; Boffetta, P.; Mundt, D.J.; Mundt, K.A. Critical review and synthesis of the epidemiologic evidence on formaldehyde exposure and risk of leukemia and other lymphohematopoietic malignancies. Cancer Causes Control 2012, 23, 1747–1766. [Google Scholar] [CrossRef] [Green Version]
- Kwon, S.C.; Kim, I.; Song, J.; Park, J. Does formaldehyde have a causal association with nasopharyngeal cancer and leukaemia? Ann. Occup. Environ. Med. 2018, 30, 5. [Google Scholar] [CrossRef] [Green Version]
- Allegra, A.; Spatari, G.; Mattioli, S.; Curti, S.; Innao, V.; Ettari, R.; Allegra, A.G.; Giorgianni, C.; Gangemi, S.; Musolino, C. Formaldehyde Exposure and Acute Myeloid Leukemia: A Review of the Literature. Medicina 2019, 55, 638. [Google Scholar] [CrossRef] [Green Version]
- Pira, E.; Romano, C.; Vecchia, C.; Boffetta, P. Hematologic and cytogenetic biomarkers of leukemia risk from formaldehyde exposure. Carcinogenesis 2017, 38, 1251–1252. [Google Scholar] [CrossRef] [PubMed]
- Kwak, K.; Paek, D.; Park, J.T. Occupational exposure to formaldehyde and risk of lung cancer: A systematic review and meta-analysis. Am. J. Ind. Med. 2020, 63, 312–327. [Google Scholar] [CrossRef] [PubMed]
- Mahboubi, A.; Koushik, A.; Siemiatycki, J.; Lavoue, J.; Rousseau, M.C. Assessment of the effect of occupational exposure to formaldehyde on the risk of lung cancer in two Canadian population-based case-control studies. Scand. J. Work Environ. Health 2013, 39, 401–410. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hu, Y.; Wu, L.; Yang, S.Q.; Wei, H.J.; Wang, C.Y.; Kang, X.; Jiang, J.M.; Zhang, P.; Tang, X.Q. Formaldehyde induces ferritinophagy to damage hippocampal neuronal cells. Toxicol. Ind. Health 2021, 37, 685–694. [Google Scholar] [CrossRef]
- Park, J.; Yang, H.S.; Song, M.K.; Kim, D.I.; Lee, K. Formaldehyde exposure induces regulatory T cell-mediated immunosuppression via calcineurin-NFAT signalling pathway. Sci. Rep. 2020, 10, 17023. [Google Scholar] [CrossRef] [PubMed]
- Tayri-Wilk, T.; Slavin, M.; Zamel, J.; Blass, A.; Cohen, S.; Motzik, A.; Sun, X.; Shalev, D.E.; Ram, O.; Kalisman, N. Mass spectrometry reveals the chemistry of formaldehyde cross-linking in structured proteins. Nat. Commun. 2020, 11, 3128. [Google Scholar] [CrossRef]
- Xie, Z.; Ge, J.; Zhang, H.; Bai, T.; He, S.; Ling, J.; Sun, H.; Zhu, Q. A highly selective two-photon fluorogenic probe for formaldehyde and its bioimaging application in cells and zebrafish. Sens. Actuators B Chem. 2017, 241, 1050–1056. [Google Scholar] [CrossRef]
- He, L.; Yang, X.; Ren, M.; Kong, X.; Liu, Y.; Lin, W. An ultra-fast illuminating fluorescent probe for monitoring formaldehyde in living cells, shiitake mushrooms, and indoors. Chem. Commun. 2016, 52, 9582–9585. [Google Scholar] [CrossRef] [Green Version]
- Im, H.; Oh, E.; Mun, J.; Khim, J.-Y.; Lee, E.; Kang, H.-S.; Kim, E.; Kim, H.; Won, N.-H.; Kim, Y.-H.; et al. Evaluation of Toxicological Monitoring Markers Using Proteomic Analysis in Rats Exposed to Formaldehyde. J. Proteome Res. 2006, 5, 1354–1366. [Google Scholar] [CrossRef]
- Roth, A.; Li, H.; Anorma, C.; Chan, J. A Reaction-Based Fluorescent Probe for Imaging of Formaldehyde in Living Cells. J. Am. Chem. Soc. 2015, 137, 10890–10893. [Google Scholar] [CrossRef]
- Zhang, Y.; Du, Y.; Li, M.; Zhang, D.; Xiang, Z.; Peng, T. Activity-Based Genetically Encoded Fluorescent and Luminescent Probes for Detecting Formaldehyde in Living Cells. Angew. Chem. Int. Ed. Engl. 2020, 59, 16352–16356. [Google Scholar] [CrossRef] [PubMed]
- Shen, L.; Dong, H.; Guo, Z.; Zhai, G.; Zhang, K. Identification of Abnormal Proteins in Plasma from Gout Patients by LC-MS/MS. Separations 2021, 8, 85. [Google Scholar] [CrossRef]
- Dong, H.Y.; Zhao, Y.J.; Bi, C.F.; Han, Y.; Zhang, J.J.; Bai, X.; Zhai, G.J.; Zhang, H.; Tian, S.S.; Hu, D.Q.; et al. TmcA functions as a lysine 2-hydroxyisobutyryltransferase to regulate transcription. Nat. Chem. Biol. 2022, 18, 142–151. [Google Scholar] [CrossRef]
- Mohanty, B.P.; Mahanty, A.; Mitra, T.; Mohanty, S.; Naik, A.K.; Parija, S.C. Proteomic and transcriptomic changes in rat liver following oral feeding of formaldehyde. Chemosphere 2020, 245, 125599. [Google Scholar] [CrossRef]
- Emirbayer, P.E.; Sinha, A.; Ignatchenko, V.; Hoyer, S.; Dorrie, J.; Schaft, N.; Pischetsrieder, M.; Kislinger, T. Proteomic Response of Human Umbilical Vein Endothelial Cells to Histamine Stimulation. Proteomics 2017, 17, 1700116. [Google Scholar] [CrossRef]
- Ahn, K.H.; Kim, S.K.; Lee, J.M.; Jeon, H.J.; Lee, D.H.; Kim, D.K. Proteomic Analysis of Bronchoalveolar Lavage Fluid Obtained from Rats Exposed to Formaldehyde. J. Health Sci. 2010, 56, 287–295. [Google Scholar] [CrossRef] [Green Version]
- Jeon, Y.-M.; Ryu, J.-C.; Lee, M.-Y. Proteomic Analysis of Differentially Expressed Proteins in Human Lung Cells Following Formaldehyde Treatment. Mol. Cell. 2007, 3, 238–245. [Google Scholar]
- Zhu, R.; Zhang, G.; Jing, M.; Han, Y.; Li, J.; Zhao, J.; Li, Y.; Chen, P.R. Genetically encoded formaldehyde sensors inspired by a protein intra-helical crosslinking reaction. Nat. Commun. 2021, 12, 581. [Google Scholar] [CrossRef]
- Nocentini, S.; Moreno, G.; Coppey, J. Survival, DNA synthesis and ribosomal RNA transcription in monkey kidney cells treated by formaldehyde. Mutat. Res. 1980, 70, 231–234. [Google Scholar] [CrossRef]
- Metz, B.; Michiels, T.; Uittenbogaard, J.; Danial, M.; Tilstra, W.; Meiring, H.D.; Hennink, W.E.; Crommelin, D.J.A.; Kersten, G.F.A.; Jiskoot, W. Identification of Formaldehyde-Induced Modifications in Diphtheria Toxin. J. Pharm. Sci. 2020, 109, 543–557. [Google Scholar] [CrossRef] [Green Version]
- Liu, J.; Chan, K.K.; Chan, W. Identification of Protein Thiazolidination as a Novel Molecular Signature for Oxidative Stress and Formaldehyde Exposure. Chem. Res. Toxicol. 2016, 29, 1865–1871. [Google Scholar] [CrossRef] [PubMed]






Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Liu, R.; Han, Y.; Wu, Z.; Zhang, J.; Zang, Y.; Shen, L.; Tian, S.; Zhang, K. Analysis of Differentially Expressed Proteins and Modifications Induced by Formaldehyde Using LC-MS/MS. Separations 2022, 9, 112. https://0-doi-org.brum.beds.ac.uk/10.3390/separations9050112
Liu R, Han Y, Wu Z, Zhang J, Zang Y, Shen L, Tian S, Zhang K. Analysis of Differentially Expressed Proteins and Modifications Induced by Formaldehyde Using LC-MS/MS. Separations. 2022; 9(5):112. https://0-doi-org.brum.beds.ac.uk/10.3390/separations9050112
Chicago/Turabian StyleLiu, Ranran, Yue Han, Zhiyue Wu, Jianji Zhang, Yong Zang, Lijin Shen, Shanshan Tian, and Kai Zhang. 2022. "Analysis of Differentially Expressed Proteins and Modifications Induced by Formaldehyde Using LC-MS/MS" Separations 9, no. 5: 112. https://0-doi-org.brum.beds.ac.uk/10.3390/separations9050112