
Effect of Low and High Doses of Two Selective Serotonin Reuptake Inhibitors on Pregnancy Outcomes and Neonatal Mortality

 , , , ,
, , , ,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Animals
2.2. Experimental Design
2.3. Blood Collection and Fluoxetine Assay
2.4. Statistical Analysis
3. Results
3.1. Systemic Fluoxetine Concentrations
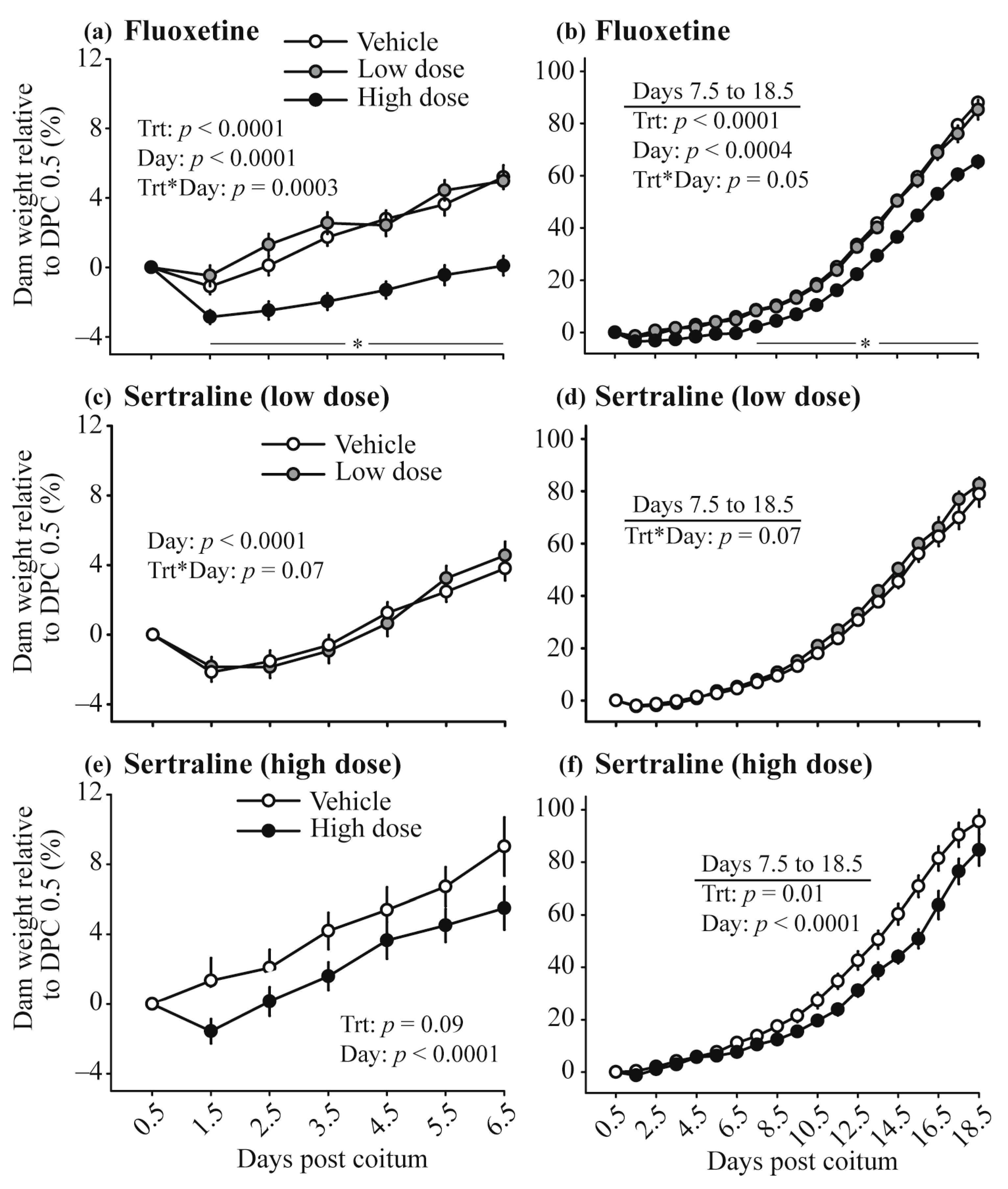
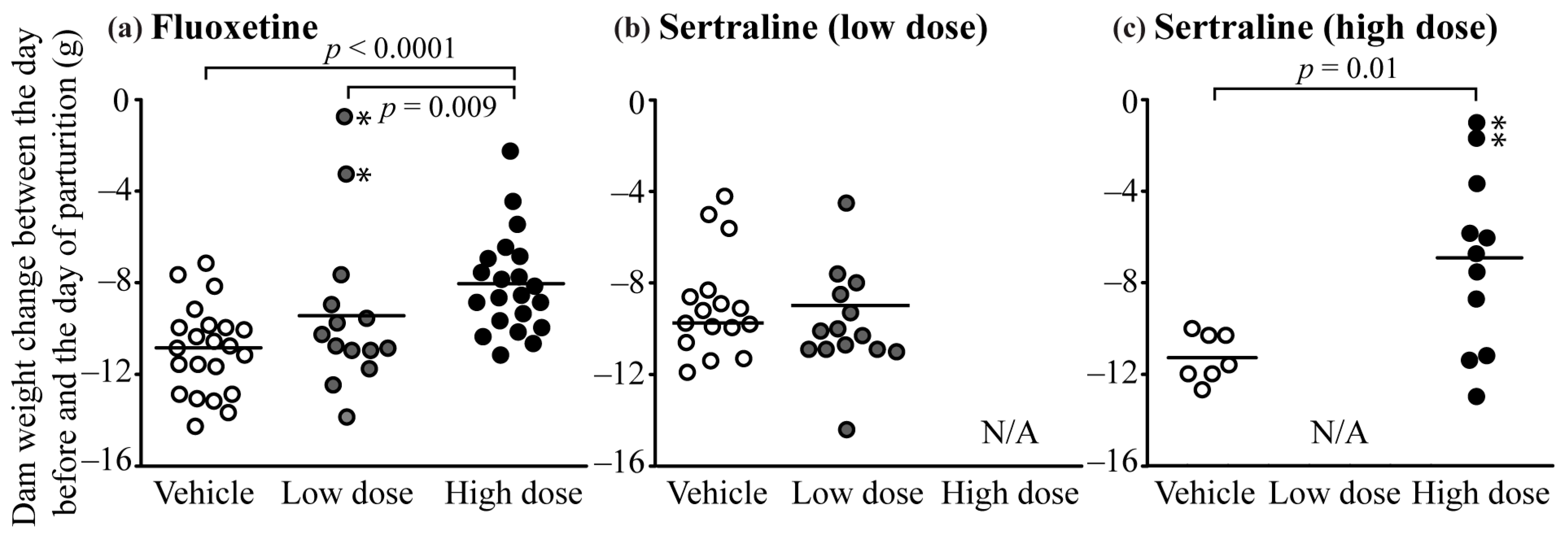
3.2. Maternal Weight
3.3. Pregnancy Establishment and Maintenance
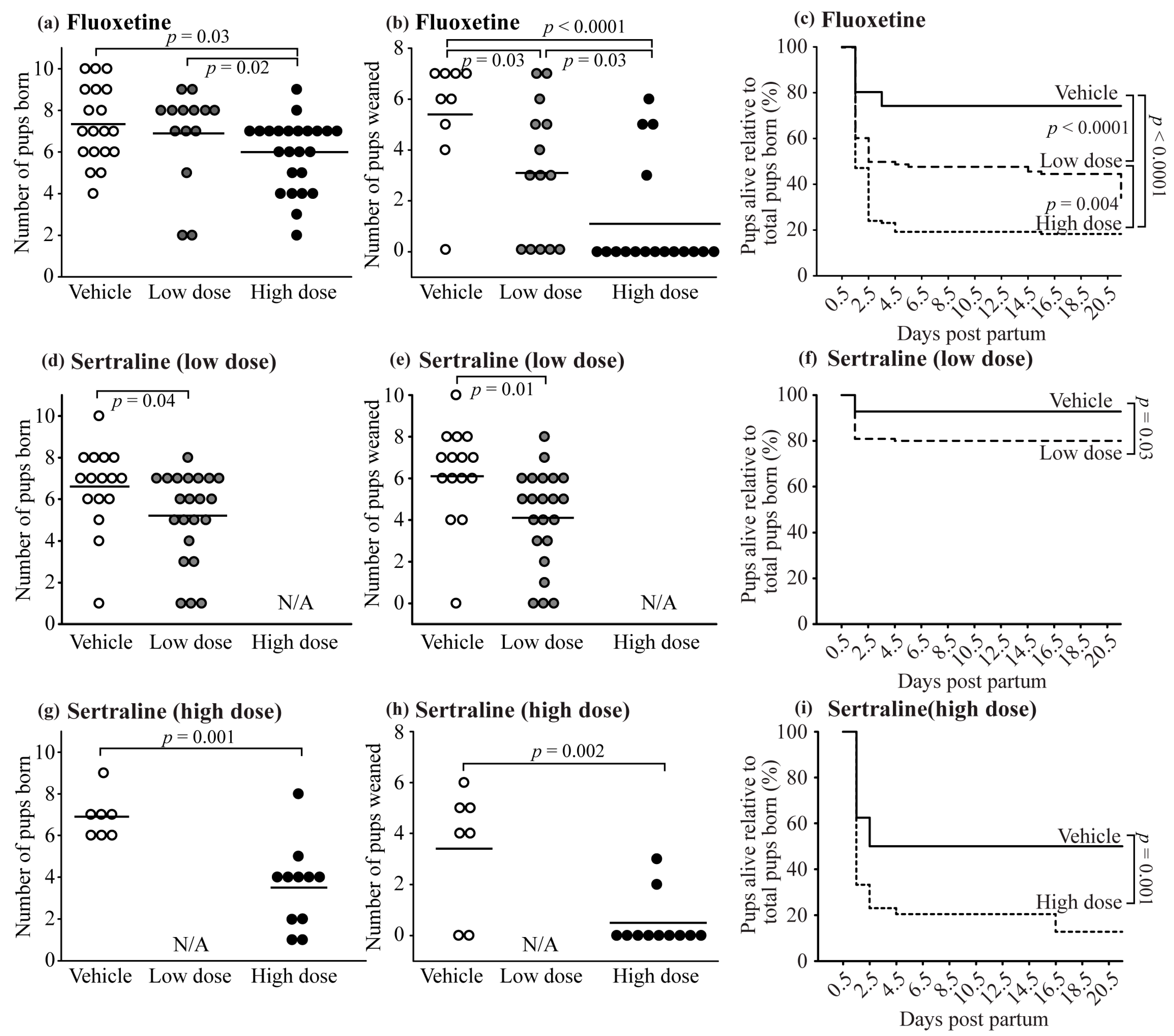
3.4. Neonatal Outcomes and Pup Survival
3.5. Pregnancy Complications
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Brajcich, M.R.; Palau, M.A.; Messer, R.D.; Murphy, M.E.; Marks, J. Why the Maternal Medication List Matters: Neonatal Toxicity From Combined Serotonergic Exposures. Pediatrics 2021, 147, e2250. [Google Scholar] [CrossRef]
- Belik, J. Fetal and Neonatal Effects of Maternal Drug Treatment for Depression. Semin. Perinatol. 2008, 32, 350–354. [Google Scholar] [CrossRef] [PubMed]
- Velasquez, J.C.; Goeden, N.; Bonnin, A. Placental serotonin: Implications for the developmental effects of SSRIs and maternal depression. Front. Cell. Neurosci. 2013, 7, 1–7. [Google Scholar] [CrossRef] [Green Version]
- Oberlander, T.F.; Warburton, W.; Misri, S.; Aghajanian, J.; Hertzman, C. Neonatal outcomes after prenatal exposure to selective serotonin reuptake inhibitor antidepressants and maternal depression using population-based linked health data. Arch. Gen. Psychiatry 2006, 63, 898–906. [Google Scholar] [CrossRef] [Green Version]
- Tran, H.; Robb, A.S. SSRI use during pregnancy. Semin. Perinatol. 2015, 39, 545–547. [Google Scholar] [CrossRef]
- Bandoli, G.; Chambers, C.D.; Wells, A.; Palmsten, K. Prenatal Antidepressant Use and Risk of Adverse Neonatal Outcomes. Pediatrics 2020, 146, e2493. [Google Scholar] [CrossRef]
- Berard, A.; Iessa, N.; Chaabane, S.; Muanda, F.T.; Boukhris, T.; Zhao, J.P. The risk of major cardiac malformations associated with paroxetine use during the first trimester of pregnancy: A systematic review and meta-analysis. Br. J. Clin. Pharmacol. 2016, 81, 589–604. [Google Scholar] [CrossRef]
- Alwan, S.; Friedman, J.M.; Chambers, C. Safety of Selective Serotonin Reuptake Inhibitors in Pregnancy: A Review of Current Evidence. CNS Drugs 2016, 30, 499–515. [Google Scholar] [CrossRef]
- Zhao, X.F.; Liu, Q.; Cao, S.X.; Pang, J.Y.; Zhang, H.J.; Feng, T.T.; Deng, Y.J.; Yao, J.; Li, H.F. A meta-analysis of selective serotonin reuptake inhibitors (SSRIs) use during prenatal depression and risk of low birth weight and small for gestational age. J. Affect. Disord. 2018, 241, 563–570. [Google Scholar] [CrossRef] [PubMed]
- Rosenfeld, C.S. Placental serotonin signaling, pregnancy outcomes, and regulation of fetal brain development. Biol. Reprod. 2020, 102, 532–538. [Google Scholar] [CrossRef] [PubMed]
- Lang, U.; Prada, J.; Clark, K.E. Systemic and uterine vascular response to serotonin in third trimester pregnant ewes. Eur. J. Obstet. Gynecol. Reprod. Biol. 1993, 51, 131–138. [Google Scholar] [CrossRef]
- Blardi, P.; De Lalla, A.; Leo, A.; Auteri, A.; Iapichino, S.; Di Muro, A.; Dell’Erba, A.; Castrogiovanni, P. Serotonin and fluoxetine levels in plasma and platelets after fluoxetine treatment in depressive patients. J. Clin. Psychopharmacol. 2002, 22, 131–136. [Google Scholar] [CrossRef]
- Morrison, J.L.; Chien, C.; Riggs, K.W.; Gruber, N.; Rurak, D. Effect of maternal fluoxetine administration on uterine blood flow, fetal blood gas status, and growth. Pediatr. Res. 2002, 51, 433–442. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Malhotra, A.; Allison, B.J.; Castillo-Melendez, M.; Jenkin, G.; Polglase, G.R.; Miller, S.L. Neonatal Morbidities of Fetal Growth Restriction: Pathophysiology and Impact. Front. Endocrinol. 2019, 10, 55. [Google Scholar] [CrossRef] [Green Version]
- Ranzil, S.; Walker, D.W.; Borg, A.J.; Wallace, E.M.; Ebeling, P.R.; Murthi, P. The relationship between the placental serotonin pathway and fetal growth restriction. Biochimie 2019, 161, 80–87. [Google Scholar] [CrossRef]
- Ranzil, S.; Ellery, S.; Walker, D.W.; Vaillancourt, C.; Alfaidy, N.; Bonnin, A.; Borg, A.; Wallace, E.M.; Ebeling, P.R.; Erwich, J.J.; et al. Disrupted placental serotonin synthetic pathway and increased placental serotonin: Potential implications in the pathogenesis of human fetal growth restriction. Placenta 2019, 84, 74–83. [Google Scholar] [CrossRef]
- Zullino, S.; Simoncini, T. Impact of selective serotonin reuptake inhibitors (SSRIs) during pregnancy and lactation: A focus on short and long-term vascular effects. Vasc. Pharmacol. 2018, 108, 74–76. [Google Scholar] [CrossRef]
- Levy, M.; Kovo, M.; Miremberg, H.; Anchel, N.; Herman, H.G.; Bar, J.; Schreiber, L.; Weiner, E. Maternal use of selective serotonin reuptake inhibitors (SSRI) during pregnancy-neonatal outcomes in correlation with placental histopathology. J. Perinatol. 2020, 40, 1017–1024. [Google Scholar] [CrossRef] [PubMed]
- Gagnon, R. Placental insufficiency and its consequences. Eur. J. Obstet. Gynecol. Reprod. Biol. 2003, 110, S99–S107. [Google Scholar] [CrossRef]
- Nardozza, L.M.M.; Caetano, A.C.R.; Zamarian, A.C.P.; Mazzola, J.B.; Silva, C.P.; Marcal, V.M.G.; Lobo, T.F.; Peixoto, A.B.; Araujo, E. Fetal growth restriction: Current knowledge. Arch. Gynecol. Obstet. 2017, 295, 1061–1077. [Google Scholar] [CrossRef] [PubMed]
- DeVane, C.L. Metabolism and pharmacokinetics of selective serotonin reuptake inhibitors. Cell. Mol. Neurobiol. 1999, 19, 443–466. [Google Scholar] [CrossRef]
- DeVane, C.L.; Liston, H.L.; Markowitz, J.S. Clinical pharmacokinetics of sertraline. Clin. Pharmacokinet. 2002, 41, 1247–1266. [Google Scholar] [CrossRef] [PubMed]
- Noorlander, C.W.; Ververs, F.F.T.; Nikkels, P.G.J.; van Echteld, C.J.A.; Visser, G.H.A.; Smidt, M.P. Modulation of Serotonin Transporter Function during Fetal Development Causes Dilated Heart Cardiomyopathy and Lifelong Behavioral Abnormalities. PLoS ONE 2008, 3, e2782. [Google Scholar] [CrossRef] [PubMed]
- Heinonen, E.; Blennow, M.; Blomdahl-Wetterholm, M.; Hovstadius, M.; Nasiell, J.; Pohanka, A.; Gustafsson, L.L.; Wide, K. Sertraline concentrations in pregnant women are steady and the drug transfer to their infants is low. Eur. J. Clin. Pharmacol. 2021, 77, 1323–1331. [Google Scholar] [CrossRef]
- Heyne, G.W.; Plisch, E.H.; Melberg, C.G.; Sandgren, E.P.; Peter, J.A.; Lipinski, R.J. A Simple and Reliable Method for Early Pregnancy Detection in Inbred Mice. J. Am. Assoc. Lab. Anim. Sci. 2015, 54, 368–371. [Google Scholar] [PubMed]
- Walia, V.; Gilhotra, N. GABAergic influence in the antidepressant effect of fluoxetine in unstressed and stressed mice. J. Appl. Pharm. Sci. 2017, 7, 141–147. [Google Scholar]
- Ma, L.; Tang, J.Y.; Zhou, J.Y.; Zhu, C.; Zhang, X.; Zhou, P.; Yu, Q.; Wang, Y.; Gu, X.J. Fluoxetine, a selective serotonin reuptake inhibitor used clinically, improves bladder function in a mouse model of moderate spinal cord injury. Neural Regen. Res. 2021, 16, 2093–2098. [Google Scholar] [CrossRef] [PubMed]
- Weaver, S.R.; Fricke, H.P.; Xie, C.; Lipinski, R.J.; Vezina, C.M.; Charles, J.F.; Hernandez, L.L. Peripartum Fluoxetine Reduces Maternal Trabecular Bone After Weaning and Elevates Mammary Gland Serotonin and PTHrP. Endocrinology 2018, 159, 2850–2862. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, J.S.; DeVane, C.L.; Gibson, B.B.; Donovan, J.L.; Markowitz, J.L.; Zhu, H.J. Population pharmacokinetic analysis of drug-drug interactions among risperidone, bupropion, and sertraline in CF1 mice. Psychopharmacology 2006, 183, 490–499. [Google Scholar] [CrossRef] [PubMed]
- Kaihola, H.; Yaldir, F.G.; Hreinsson, J.; Hörnaeus, K.; Bergquist, J.; Olivier, J.D.; Åkerud, H.; Sundström-Poromaa, I. Effects of fluoxetine on human embryo development. Front. Cell. Neurosci. 2016, 10, 160. [Google Scholar] [CrossRef] [Green Version]
- Uphouse, L.; Hensler, J.G.; Sarkar, J.; Grossie, B. Fluoxetine disrupts food intake and estrous cyclicity in Fischer female rats. Brain Res. 2006, 1072, 79–90. [Google Scholar] [CrossRef]
- Hansen, C.H.; Larsen, L.W.; Sorensen, A.M.; Halling-Sorensen, B.; Styrishave, B. The six most widely used selective serotonin reuptake inhibitors decrease androgens and increase estrogens in the H295R cell line. Toxicol. In Vitro 2017, 41, 1–11. [Google Scholar] [CrossRef] [PubMed]
- Jacobsen, N.W.; Hansen, C.H.; Nellemann, C.; Styrishave, B.; Halling-Sorensen, B. Effects of selective serotonin reuptake inhibitors on three sex steroids in two versions of the aromatase enzyme inhibition assay and in the H295R cell assay. Toxicol. In Vitro 2015, 29, 1729–1735. [Google Scholar] [CrossRef]
- Muller, J.C.; Imazaki, P.H.; Boareto, A.C.; Lourenco, E.L.B.; Golin, M.; Vechi, M.F.; Lombardi, N.F.; Minatovicz, B.C.; Scippo, M.L.; Martino-Andrade, A.J.; et al. In vivo and in vitro estrogenic activity of the antidepressant fluoxetine. Reprod. Toxicol. 2012, 34, 80–85. [Google Scholar] [CrossRef]
- Cartwright, J.E.; Fraser, R.; Leslie, K.; Wallace, A.E.; James, J.L. Remodelling at the maternal-fetal interface: Relevance to human pregnancy disorders. Reproduction 2010, 140, 803–813. [Google Scholar] [CrossRef] [Green Version]
- Kjaersgaard, M.I.S.; Parner, E.T.; Vestergaard, M.; Sorensen, M.J.; Olsen, J.; Christensen, J.; Bech, B.H.; Pedersen, L.H. Prenatal Antidepressant Exposure and Risk of Spontaneous Abortion—A Population-Based Study. PLoS ONE 2013, 8, e72095. [Google Scholar] [CrossRef] [Green Version]
- Almeida, N.D.; Basso, O.; Abrahamowicz, M.; Gagnon, R.; Tamblyn, R. Risk of Miscarriage in Women Receiving Antidepressants in Early Pregnancy, Correcting for Induced Abortions. Epidemiology 2016, 27, 538–546. [Google Scholar] [CrossRef] [PubMed]
- Bauer, S.; Monk, C.; Ansorge, M.; Gyamfi, C.; Myers, M. Impact of antenatal selective serotonin reuptake inhibitor exposure on pregnancy outcomes in mice. Am. J. Obstet. Gynecol. 2010, 203, 375.e1–375.e4. [Google Scholar] [CrossRef] [PubMed]
- Cabrera, R.M.; Lin, Y.L.; Law, E.; Kim, J.; Wlodarczyk, B.J. The teratogenic effects of sertraline in mice. Birth Defects Res. 2020, 112, 1014–1024. [Google Scholar] [CrossRef]
- Muller, J.C.; Boareto, A.C.; Lourenco, E.L.B.; Zaia, R.M.; Kienast, M.F.; Spercoski, K.M.; Morais, R.N.; Martino-Andrade, A.J.; Dalsenter, P.R. In Utero and Lactational Exposure to Fluoxetine in Wistar Rats: Pregnancy Outcomes and Sexual Development. Basic Clin. Pharmacol. Toxicol. 2013, 113, 132–140. [Google Scholar] [CrossRef]
- Vedernikov, Y.; Bolanos, S.; Bytautiene, E.; Fulep, E.; Saade, G.R.; Garfield, R.E. Effect of fluoxetine on contractile activity of pregnant rat uterine rings. Am. J. Obstet. Gynecol. 2000, 182, 296–299. [Google Scholar] [CrossRef]
- Sparenborg, S. Mortality in neonatal rats is increased by moderate prenatal exposure to some monoamine reuptake inhibitors—A brief review. In Cocaine: Effects on the Developing Brain; Harvey, J.A., Kosofsky, B.E., Eds.; Academy of Sciences: New York, NY, USA, 1998; Volume 846, pp. 423–426. [Google Scholar]
- Haskell, S.E.; Hermann, G.M.; Reinking, B.E.; Volk, K.A.; Peotta, V.A.; Zhu, V.; Roghair, R.D. Sertraline exposure leads to small left heart syndrome in adult mice. Pediatr. Res. 2013, 73, 286–293. [Google Scholar] [CrossRef]
- Marchand, G.J.; Meassick, K.; Wolf, H.; Hopewell, S.K.; Sainz, K.; Anderson, S.M.; Ware, K.; Vallejo, J.; King, A.; Ruther, S.; et al. Respiratory depression in a neonate born to mother on maximum dose sertraline: A case report. J. Med. Case Rep. 2021, 15, 1–5. [Google Scholar] [CrossRef]
- Velasquez, J.C.; Bonnin, A. Placental Transport and Metabolism: Implications for the Developmental Effects of Selective Serotonin Reuptake Inhibitors (SSRI) Antidepressants. In Prenatal and Postnatal Determinants of Development; Walker, D.W., Ed.; Humana Press Inc.: Totowa, NJ, USA, 2016; Volume 109, pp. 245–262. [Google Scholar]
- Haskell, S.E.; Lo, C.; Kent, M.E.; Eggleston, T.M.; Volk, K.A.; Reinking, B.E.; Roghair, R.D. Cardiac Outcomes After Perinatal Sertraline Exposure in Mice. J. Cardiovasc. Pharmacol. 2017, 70, 119–127. [Google Scholar] [CrossRef] [PubMed]
- Horackova, H.; Karahoda, R.; Cerveny, L.; Vachalova, V.; Ebner, R.; Abad, C.; Staud, F. Effect of Selected Antidepressants on Placental Homeostasis of Serotonin: Maternal and Fetal Perspectives. Pharmaceutics 2021, 13, 1306. [Google Scholar] [CrossRef]
- Bonnin, A.; Goeden, N.; Chen, K.; Wilson, M.L.; King, J.; Shih, J.C.; Blakely, R.D.; Deneris, E.S.; Levitt, P. A transient placental source of serotonin for the fetal forebrain. Nature 2011, 472, 347–352. [Google Scholar] [CrossRef] [Green Version]
- Kliman, H.J.; Quaratella, S.B.; Setaro, A.C.; Siegman, E.C.; Subha, Z.T.; Tal, R.; Milano, K.M.; Steck, T.L. Pathway of Maternal Serotonin to the Human Embryo and Fetus. Endocrinology 2018, 159, 1609–1629. [Google Scholar] [CrossRef]
- Karahoda, R.; Horackova, H.; Kastner, P.; Matthios, A.; Cerveny, L.; Kucera, R.; Kacerovsky, M.; Tebbens, J.D.; Bonnin, A.; Abad, C.; et al. Serotonin homeostasis in the materno-foetal interface at term: Role of transporters (SERT/SLC6A4 and OCT3/SLC22A3) and monoamine oxidase A (MAO-A) in uptake and degradation of serotonin by human and rat term placenta. Acta Physiol. 2020, 229, e13478. [Google Scholar] [CrossRef]
- Gasser, P.J.; Lowry, C.A. Organic cation transporter 3: A cellular mechanism underlying rapid, non-genomic glucocorticoid regulation of monoaminergic neurotransmission, physiology, and behavior. Horm. Behav. 2018, 104, 173–182. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sabbioni, C.; Bugamelli, F.; Varani, G.; Mercolini, L.; Musenga, A.; Saracino, M.A.; Fanali, S.; Raggi, M.A. A rapid HPLC-DAD method for the analysis of fluoxetine and norfluoxetine in plasma from overdose patients. J. Pharm. Biomed. Anal. 2004, 36, 351–356. [Google Scholar] [CrossRef] [PubMed]
- Aggarwal, A.; Jethani, S.L.; Rohatgi, R.K.; Kalra, J. Selective Serotonin Re-uptake Inhibitors (SSRIs) Induced Weight Changes: A Dose and Duration Dependent Study on Albino Rats. J. Clin. Diagn. Res. 2016, 10, AF1–AF3. [Google Scholar] [CrossRef] [PubMed]
- Ferguson, J.M. SSRI antidepressant medications: Adverse effects and tolerability. Prim. Care Companion J. Clin. Psychiatry 2001, 3, 22. [Google Scholar] [CrossRef] [PubMed]




{kind=link}
{kind=link}
{kind=link}
| Vehicle | Low Dose | High Dose | p Value | |
|---|---|---|---|---|
| DPC 0.75, ng/mL (range) | 0 Undetectable | 195.4 ± 14.6 b (169.5−219.9) | 4724.7 ± 354.0 a (2756.3–5828.4) | <0.0001 |
| DPC 17.75, ng/mL (range) | 0 Undetectable | 466.8 ± 61.6 b (336.8–746.6) | 20,059.2 ± 2176.5 a (13,477.8–29,815.6) | <0.0001 |
| Vehicle | Low Dose | High Dose | p Value | |
|---|---|---|---|---|
| Fluoxetine | ||||
| Vaginal plug, n | 28 | 32 | 127 | - |
| Pregnant dams, n | 24 | 25 | 24 | - |
| Pregnancy per plug, % | 85.7 a | 78.1 a | 18.9 b | <0.0001 |
| Gestation length, day | 19.1 ± 0.1 | 19.1 ± 0.1 | 18.9 ± 0.1 | 0.17 |
| Sertraline low dose | ||||
| Vaginal plug, n | 32 | 32 | N/A | - |
| Pregnant dams, n | 22 | 23 | N/A | - |
| Pregnancy per plug, % | 68.8 | 71.9 | N/A | 0.99 |
| Gestation length, day | 18.8 ± 0.1 B | 19.0 ± 0.1 A | N/A | 0.096 |
| Sertraline high dose | ||||
| Vaginal plug, n | 11 | N/A | 16 | - |
| Pregnant dams, n | 7 | N/A | 11 | - |
| Pregnancy per plug, % | 63.6 | N/A | 68.7 | 0.9 |
| Gestation length, day | 18.9 ± 0.3 b | N/A | 19.6 ± 0.2 a | 0.01 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Domingues, R.R.; Fricke, H.P.; Sheftel, C.M.; Bell, A.M.; Sartori, L.C.; Manuel, R.S.J.; Krajco, C.J.; Wiltbank, M.C.; Hernandez, L.L. Effect of Low and High Doses of Two Selective Serotonin Reuptake Inhibitors on Pregnancy Outcomes and Neonatal Mortality. Toxics 2022, 10, 11. https://0-doi-org.brum.beds.ac.uk/10.3390/toxics10010011
Domingues RR, Fricke HP, Sheftel CM, Bell AM, Sartori LC, Manuel RSJ, Krajco CJ, Wiltbank MC, Hernandez LL. Effect of Low and High Doses of Two Selective Serotonin Reuptake Inhibitors on Pregnancy Outcomes and Neonatal Mortality. Toxics. 2022; 10(1):11. https://0-doi-org.brum.beds.ac.uk/10.3390/toxics10010011
Chicago/Turabian StyleDomingues, Rafael R., Hannah P. Fricke, Celeste M. Sheftel, Autumn M. Bell, Luma C. Sartori, Robbie S. J. Manuel, Chandler J. Krajco, Milo C. Wiltbank, and Laura L. Hernandez. 2022. "Effect of Low and High Doses of Two Selective Serotonin Reuptake Inhibitors on Pregnancy Outcomes and Neonatal Mortality" Toxics 10, no. 1: 11. https://0-doi-org.brum.beds.ac.uk/10.3390/toxics10010011