
Effects of Digestion, Cell Culture Media, and Mucous on the Physical Properties, Cellular Effects, and Translocation of Polystyrene and Polymethacrylate Nanoparticles

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Nanoparticle Properties
2.2. In Vitro Digestion Procedure
2.3. Cell Culture
2.4. Nanoparticle Translocation Experiments
2.5. Statistics
3. Results
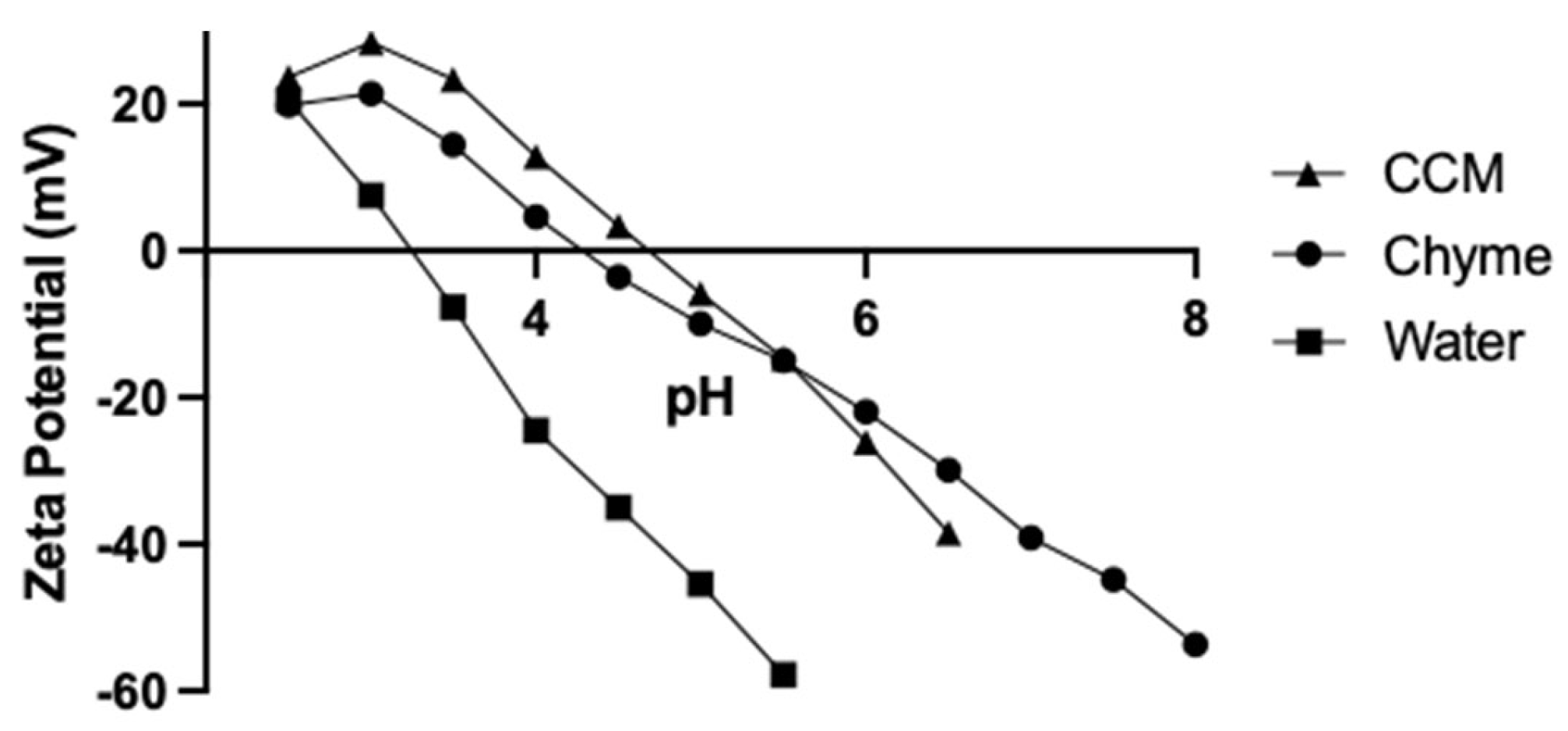
3.1. Effect of Digestion on Nanoparticle Hydrodynamic Diameter and Zeta Potential
3.2. The Effects of Nanoparticle Exposure on the Monolayer Barrier Properties and Translocation
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Sharma, S.; Parveen, R.; Chatterji, B.P. Toxicology of Nanoparticles in Drug Delivery. Curr. Pathobiol. Rep. 2021, 9, 133–144. [Google Scholar] [CrossRef] [PubMed]
- Suriya Prabha, A.; Dorothy, R.; Jancirani, S.; Rajendran, S.; Singh, G.; Senthil Kumaran, S. Chapter 7—Recent advances in the study of toxicity of polymer-based nanomaterials. In Nanotoxicity; Rajendran, S., Mukherjee, A., Nguyen, T.A., Godugu, C., Shukla, R.K., Eds.; Elsevier: Chennai, India, 2020; pp. 143–165. [Google Scholar]
- Sorasan, C.; Edo, C.; González-Pleiter, M.; Fernández-Piñas, F.; Leganés, F.; Rodríguez, A.; Rosal, R. Ageing and fragmentation of marine microplastics. Sci. Total Environ. 2022, 827, 154438. [Google Scholar] [CrossRef]
- Horton, A.A. Plastic pollution: When do we know enough? J. Hazard. Mater. 2022, 422, 126885. [Google Scholar] [CrossRef] [PubMed]
- Voigt, N.; Henrich-Noack, P.; Kockentiedt, S.; Hintz, W.; Tomas, J.; Sabel, B.A. Toxicity of polymeric nanoparticles in vivo and in vitro. J. Nanoparticle Res. 2014, 16, 2379. [Google Scholar] [CrossRef] [PubMed]
- Weber, A.; Schwiebs, A.; Solhaug, H.; Stenvik, J.; Nilsen, A.M.; Wagner, M.; Relja, B.; Radeke, H.H. Nanoplastics affect the inflammatory cytokine release by primary human monocytes and dendritic cells. Environ. Int. 2022, 163, 107173. [Google Scholar] [CrossRef]
- Stock, V.; Böhmert, L.; Coban, G.; Tyra, G.; Vollbrecht, M.-L.; Voss, L.; Paul, M.B.; Braeuning, A.; Sieg, H. Microplastics and nanoplastics: Size, surface and dispersant–What causes the effect? Toxicol. In Vitro 2022, 80, 105314. [Google Scholar] [CrossRef]
- Shen, M.; Zhang, Y.; Zhu, Y.; Song, B.; Zeng, G.; Hu, D.; Wen, X.; Ren, X. Recent advances in toxicological research of nanoplastics in the environment: A review. Environ. Pollut. 2019, 252, 511–521. [Google Scholar] [CrossRef]
- Walczyk, D.; Bombelli, F.B.; Monopoli, M.P.; Lynch, I.; Dawson, K.A. What the cell “sees” in bionanoscience. J. Am. Chem. Soc. 2010, 132, 5761–5768. [Google Scholar] [CrossRef]
- Badawy, A.M.E.; Luxton, T.P.; Silva, R.G.; Scheckel, K.G.; Suidan, M.T.; Tolaymat, T.M. Impact of environmental conditions (pH, ionic strength, and electrolyte type) on the surface charge and aggregation of silver nanoparticles suspensions. Environ. Sci. Technol. 2010, 44, 1260–1266. [Google Scholar] [CrossRef]
- Zhang, T.; Zhu, G.; Lu, B.; Qian, Z.; Peng, Q. Protein corona formed in the gastrointestinal tract and its impacts on oral delivery of nanoparticles. Med. Res. Rev. 2021, 41, 1835–1850. [Google Scholar] [CrossRef]
- Ji, Y.; Wang, Y.; Shen, D.; Kang, Q.; Chen, L. Mucin corona delays intracellular trafficking and alleviates cytotoxicity of nanoplastic-benzopyrene combined contaminant. J. Hazard. Mater. 2021, 406, 124306. [Google Scholar] [CrossRef] [PubMed]
- Lundqvist, M. Nanoparticles: Tracking protein corona over time. Nat. Nanotechnol. 2013, 8, 701–702. [Google Scholar] [CrossRef] [PubMed]
- Inkielewicz-Stepniak, I.; Tajber, L.; Behan, G.; Zhang, H.; Radomski, M.W.; Medina, C.; Santos-Martinez, M.J. The role of mucin in the toxicological impact of polystyrene nanoparticles. Materials 2018, 11, 724. [Google Scholar] [CrossRef]
- Bhattacharjee, S.; Mahon, E.; Harrison, S.M.; McGetrick, J.; Muniyappa, M.; Carrington, S.D.; Brayden, D.J. Nanoparticle passage through porcine jejunal mucus: Microfluidics and rheology. Nanomed. Nanotechnol. Biol. Med. 2017, 13, 863–873. [Google Scholar] [CrossRef] [PubMed]
- Johansson, M.E.; Sjövall, H.; Hansson, G.C. The gastrointestinal mucus system in health and disease. Nat. Rev. Gastroenterol. Hepatol. 2013, 10, 352–361. [Google Scholar] [CrossRef]
- Guri, A.; Gülseren, I.; Corredig, M. Utilization of solid lipid nanoparticles for enhanced delivery of curcumin in cocultures of HT29-MTX and Caco-2 cells. Food Funct. 2013, 4, 1410–1419. [Google Scholar] [CrossRef]
- Lunov, O.; Syrovets, T.; Loos, C.; Nienhaus, G.U.; Mailänder, V.; Landfester, K.; Rouis, M.; Simmet, T. Amino-functionalized polystyrene nanoparticles activate the NLRP3 inflammasome in human macrophages. ACS Nano 2011, 5, 9648–9657. [Google Scholar] [CrossRef]
- Busch, M. Development and Application of Intestinal In Vitro Models to Investigate the Link between Plastic Particle Toxicity, Intestinal Inflammation, and the NLRP3 Inflammasome. Ph.D. Thesis, Heinrich-Heine-Universität Düsseldorf, Düsseldorf, Germany, 2022. [Google Scholar]
- Hu, Q.; Wang, H.; He, C.; Jin, Y.; Fu, Z. Polystyrene nanoparticles trigger the activation of p38 MAPK and apoptosis via inducing oxidative stress in zebrafish and macrophage cells. Environ. Pollut. 2021, 269, 116075. [Google Scholar] [CrossRef]
- Garcia-Rodriguez, A.; Vila, L.; Cortes, C.; Hernandez, A.; Marcos, R. Exploring the usefulness of the complex in vitro intestinal epithelial model Caco-2/HT29/Raji-B in nanotoxicology. Food Chem. Toxicol. 2018, 113, 162–170. [Google Scholar] [CrossRef]
- des Rieux, A.; Fievez, V.; Théate, I.; Mast, J.; Préat, V.; Schneider, Y.-J. An improved in vitro model of human intestinal follicle-associated epithelium to study nanoparticle transport by M cells. Eur. J. Pharm. Sci. 2007, 30, 380–391. [Google Scholar] [CrossRef]
- Busch, M.; Kämpfer, A.A.; Schins, R.P. An inverted in vitro triple culture model of the healthy and inflamed intestine: Adverse effects of polyethylene particles. Chemosphere 2021, 284, 131345. [Google Scholar] [CrossRef] [PubMed]
- Stojkovic, M.; Stojkovic, P.; Stankovic, K.M. Human pluripotent stem cells—Unique tools to decipher the effects of environmental and intracellular plastic pollution on human health. Environ. Pollut. 2021, 269, 116144. [Google Scholar] [CrossRef] [PubMed]
- Kämpfer, A.A.M.; Busch, M.; Büttner, V.; Bredeck, G.; Stahlmecke, B.; Hellack, B.; Masson, I.; Sofranko, A.; Albrecht, C.; Schins, R.P.F. Model Complexity as Determining Factor for In Vitro Nanosafety Studies: Effects of Silver and Titanium Dioxide Nanomaterials in Intestinal Models. Small 2021, 17, 2004223. [Google Scholar] [CrossRef] [PubMed]
- Rubio, L.; Marcos, R.; Hernández, A. Potential adverse health effects of ingested micro- and nanoplastics on humans. Lessons learned from in vivo and in vitro mammalian models. J. Toxicol. Environ. Health Part B 2020, 23, 51–68. [Google Scholar] [CrossRef] [PubMed]
- Liu, S.; Wu, X.; Gu, W.; Yu, J.; Wu, B. Influence of the digestive process on intestinal toxicity of polystyrene microplastics as determined by in vitro Caco-2 models. Chemosphere 2020, 256, 127204. [Google Scholar] [CrossRef] [PubMed]
- Předota, M.; Machesky, M.L.; Wesolowski, D.J. Molecular origins of the zeta potential. Langmuir 2016, 32, 10189–10198. [Google Scholar] [CrossRef]
- Lefebvre, D.E.; Venema, K.; Gombau, L.; Valerio, L.G.; Raju, J.; Bondy, G.S.; Bouwmeester, H.; Singh, R.P.; Clippinger, A.J.; Collnot, E.-M.; et al. Utility of models of the gastrointestinal tract for assessment of the digestion and absorption of engineered nanomaterials released from food matrices. Nanotoxicology 2015, 9, 523–542. [Google Scholar] [CrossRef]
- Hattori, T.; Arizono, N. Cell kinetics and secretion of mucus in the gastrointestinal mucosa, and their diurnal rhythm. J. Clin. Gastroenterol. 1988, 10, S1–S6. [Google Scholar] [CrossRef]
- DeLoid, G.; Cohen, J.M.; Darrah, T.; Derk, R.; Rojanasakul, L.; Pyrgiotakis, G.; Wohlleben, W.; Demokritou, P. Estimating the effective density of engineered nanomaterials for in vitro dosimetry. Nat. Commun. 2014, 5, 3514. [Google Scholar] [CrossRef]
- Khanbeigi, R.A.; Kumar, A.; Sadouki, F.; Lorenz, C.; Forbes, B.; Dailey, L.A.; Collins, H. The delivered dose: Applying particokinetics to in vitro investigations of nanoparticle internalization by macrophages. J. Control. Release 2012, 162, 259–266. [Google Scholar] [CrossRef]
- Paredes, A.; Acuña, S.M.; Gutiérrez, L.; Toledo, P.G. Zeta potential of pyrite particles in concentrated solutions of monovalent seawater electrolytes and amyl xanthate. Minerals 2019, 9, 584. [Google Scholar] [CrossRef]
- Versantvoort, C.H.M.; Oomen, A.G.; Van de Kamp, E.; Rompelberg, C.J.M.; Sips, A.J.A.M. Applicability of an in vitro digestion model in assessing the bioaccessibility of mycotoxins from food. Food Chem. Toxicol. 2005, 43, 31–40. [Google Scholar] [CrossRef]
- Walczak, A.P.; Kramer, E.; Hendriksen, P.J.; Helsdingen, R.; van der Zande, M.; Rietjens, I.M.; Bouwmeester, H. In vitro gastrointestinal digestion increases the translocation of polystyrene nanoparticles in an in vitro intestinal co-culture model. Nanotoxicology 2015, 9, 886–894. [Google Scholar] [CrossRef]
- Busch, M.; Bredeck, G.; Kämpfer, A.A.M.; Schins, R.P.F. Investigations of acute effects of polystyrene and polyvinyl chloride micro- and nanoplastics in an advanced in vitro triple culture model of the healthy and inflamed intestine. Environ. Res. 2021, 193, 110536. [Google Scholar] [CrossRef] [PubMed]
- Walczak, A.P.; Fokkink, R.; Peters, R.; Tromp, P.; Herrera Rivera, Z.E.; Rietjens, I.M.; Hendriksen, P.J.; Bouwmeester, H. Behaviour of silver nanoparticles and silver ions in an in vitro human gastrointestinal digestion model. Nanotoxicology 2013, 7, 1198–1210. [Google Scholar] [CrossRef]
- Srinivasan, B.; Kolli, A.R.; Esch, M.B.; Abaci, H.E.; Shuler, M.L.; Hickman, J.J. TEER measurement techniques for in vitro barrier model systems. SLAS Technol. 2015, 20, 107–126. [Google Scholar] [CrossRef]
- Farnham, R.; Stafford, S.H.; Sutton, S.C. Impact of Dose on Amount of Polystyrene Nanoparticles Translocated across Caco-2 Cell Monolayers. In Proceedings of the AAPS Northeastern Regional Diuscussion Group, Framingham, CT, USA; 2017. [Google Scholar]
- Pires, C.L.; Praça, C.; Martins, P.A.; Batista de Carvalho, A.L.; Ferreira, L.; Marques, M.P.M.; Moreno, M.J. Re-use of caco-2 monolayers in permeability assays—Validation regarding cell monolayer integrity. Pharmaceutics 2021, 13, 1563. [Google Scholar] [CrossRef] [PubMed]
- Schwarzfischer, M.; Rogler, G. The Intestinal Barrier—Shielding the Body from Nano-and Microparticles in Our Diet. Metabolites 2022, 12, 223. [Google Scholar] [CrossRef] [PubMed]
- Pedata, P.; Ricci, G.; Malorni, L.; Venezia, A.; Cammarota, M.; Volpe, M.G.; Iannaccone, N.; Guida, V.; Schiraldi, C.; Romano, M. In vitro intestinal epithelium responses to titanium dioxide nanoparticles. Food Res. Int. 2019, 119, 634–642. [Google Scholar] [CrossRef] [PubMed]
- Vita, A.A.; Royse, E.A.; Pullen, N.A. Nanoparticles and danger signals: Oral delivery vehicles as potential disruptors of intestinal barrier homeostasis. J. Leukoc. Biol. 2019, 106, 95–103. [Google Scholar] [CrossRef]
- Brown, T.D.; Whitehead, K.A.; Mitragotri, S. Materials for oral delivery of proteins and peptides. Nat. Rev. Mater. 2020, 5, 127–148. [Google Scholar]
- Sutton, S.C.; Forbes, A.E.; Cargill, R.; Hochman, J.H.; LeCluyse, E.L. Simultaneous in Vitro Measurement of Intestinal Tissue Permeability and Transepithelial Electrical Resistance (TEER) Using Sweetana–Grass Diffusion Cells. Pharm. Res. 1992, 9, 316–319. [Google Scholar] [CrossRef]
- Boraschi, D.; Canesi, L.; Drobne, D.; Kemmerling, B.; Pinsino, A.; Prochazkova, P. Interaction between nanomaterials and the innate immune system across evolution. Biol. Rev. 2023, 98, 747–774. [Google Scholar] [CrossRef] [PubMed]
- Amaral-Zettler, L.A.; Zettler, E.R.; Slikas, B.; Boyd, G.D.; Melvin, D.W.; Morrall, C.E.; Proskurowski, G.; Mincer, T.J. The biogeography of the Plastisphere: Implications for policy. Front. Ecol. Environ. 2015, 13, 541–546. [Google Scholar] [CrossRef] [PubMed]
- Hinderliter, P.M.; Minard, K.R.; Orr, G.; Chrisler, W.B.; Thrall, B.D.; Pounds, J.G.; Teeguarden, J.G. ISDD: A computational model of particle sedimentation, diffusion and target cell dosimetry for in vitro toxicity studies. Part. Fibre Toxicol. 2010, 7, 36. [Google Scholar] [CrossRef]






Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Jabor, Z.; Sutton, S.C. Effects of Digestion, Cell Culture Media, and Mucous on the Physical Properties, Cellular Effects, and Translocation of Polystyrene and Polymethacrylate Nanoparticles. Toxics 2023, 11, 708. https://0-doi-org.brum.beds.ac.uk/10.3390/toxics11080708
Jabor Z, Sutton SC. Effects of Digestion, Cell Culture Media, and Mucous on the Physical Properties, Cellular Effects, and Translocation of Polystyrene and Polymethacrylate Nanoparticles. Toxics. 2023; 11(8):708. https://0-doi-org.brum.beds.ac.uk/10.3390/toxics11080708
Chicago/Turabian StyleJabor, Zainab, and Steven C. Sutton. 2023. "Effects of Digestion, Cell Culture Media, and Mucous on the Physical Properties, Cellular Effects, and Translocation of Polystyrene and Polymethacrylate Nanoparticles" Toxics 11, no. 8: 708. https://0-doi-org.brum.beds.ac.uk/10.3390/toxics11080708