
Cumulative Protective Effect of Melatonin and Indole-3-Propionic Acid against KIO3—Induced Lipid Peroxidation in Porcine Thyroid


{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Chemicals
2.2. Animals
2.3. Assay of Lipid Peroxidation
2.4. Measurement of Lipid Peroxidation Products
2.5. Statistical Analyses
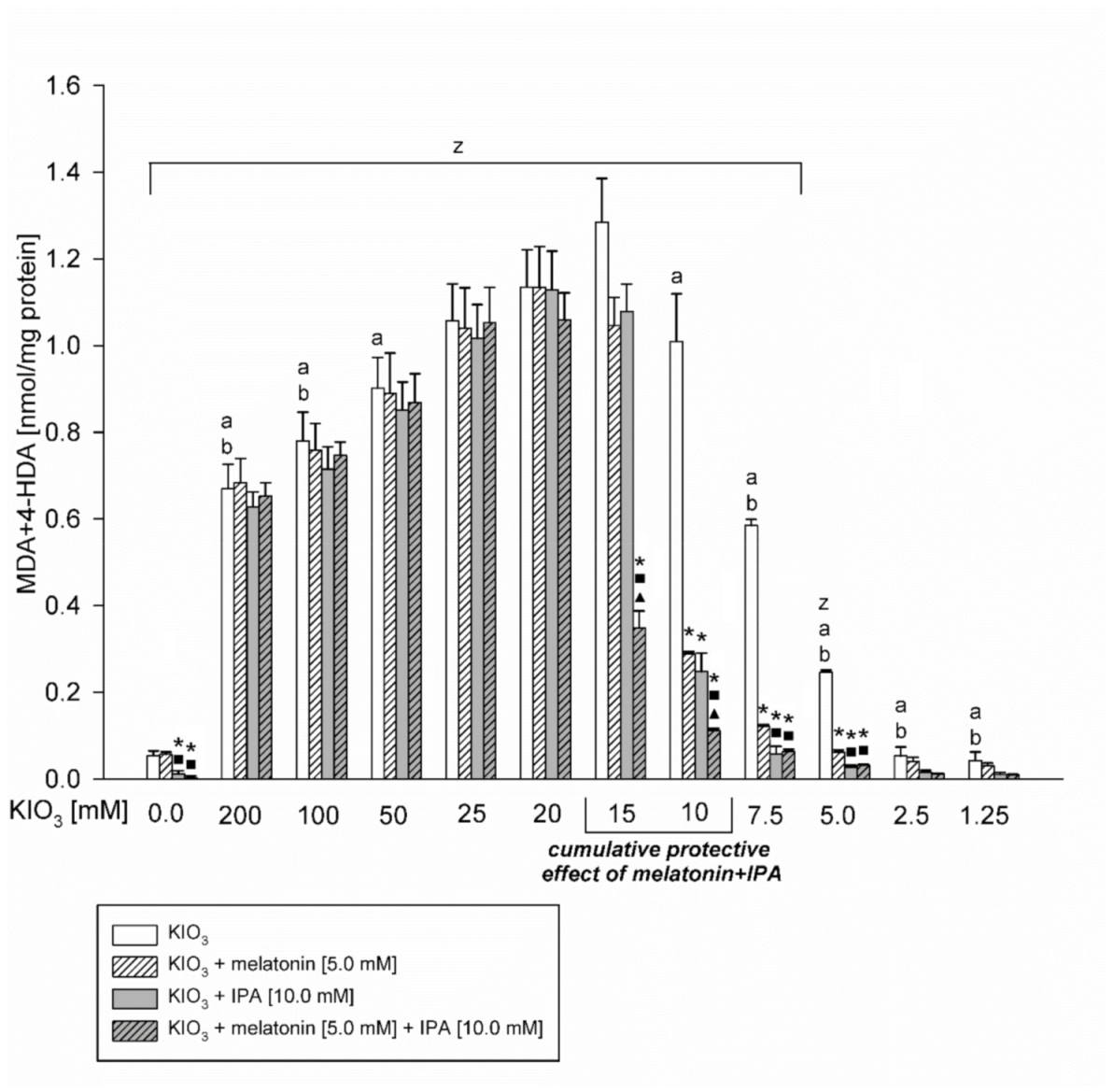
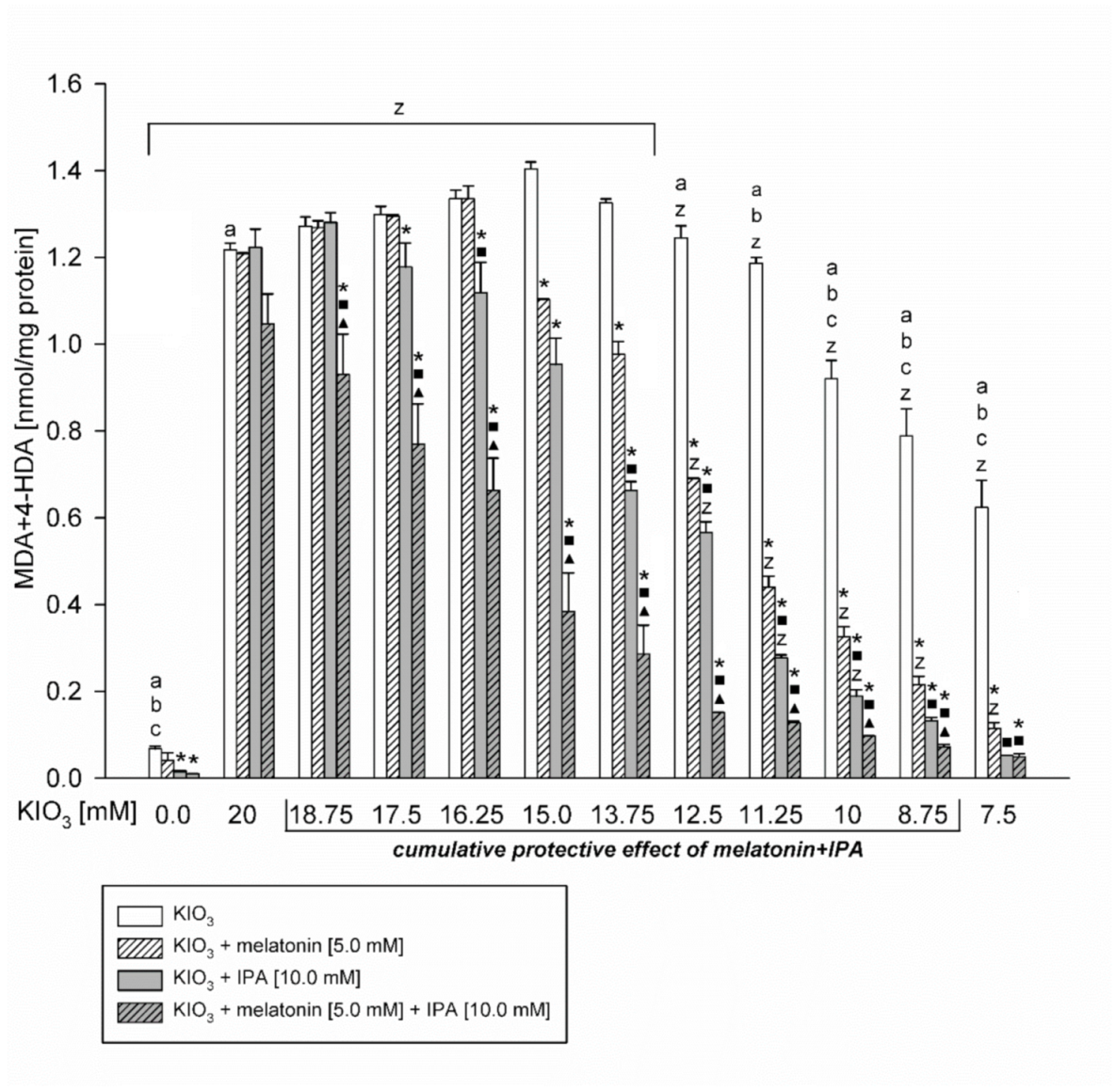
3. Results
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Hunyadi, A. The mechanism(s) of action of antioxidants: From scavenging reactive oxygen/nitrogen species to redox signaling and the generation of bioactive secondary metabolites. Med. Res. Rev. 2019, 39, 2505–2533. [Google Scholar] [CrossRef] [Green Version]
- Schieber, M.; Chandel, N.S. ROS function in redox signaling and oxidative stress. Curr. Biol. 2014, 24, R453–R462. [Google Scholar] [CrossRef] [Green Version]
- Gorrini, C.; Harris, I.S.; Mak, T.W. Modulation of oxidative stress as an anticancer strategy. Nat. Rev. Drug Discov. 2013, 12, 931–947. [Google Scholar] [CrossRef]
- Carvalho, D.P.; Dupuy, C. Thyroid hormone biosynthesis and release. Mol. Cell Endocrinol. 2017, 458, 6–15. [Google Scholar] [CrossRef]
- Karbownik-Lewinska, M.; Kokoszko-Bilska, A. Oxidative damage to macromolecules in the thyroid—Experimental evidence. Thyroid Res. 2012, 5, 25. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Alsen, M.; Sinclair, C.; Cooke, P.; Ziadkhanpour, K.; Genden, E.; van Gerwen, M. Endocrine Disrupting Chemicals and Thyroid Cancer: An Overview. Toxics 2021, 9, 14. [Google Scholar] [CrossRef]
- Zimmermann, M.B.; Galetti, V. Iodine intake as a risk factor for thyroid cancer: A comprehensive review of animal and human studies. Thyroid Res. 2015, 8, 8. [Google Scholar] [CrossRef] [Green Version]
- Szybinski, Z.; Delange, F.; Lewinski, A.; Podoba, J.; Rybakowa, M.; Wasik, R.; Szewczyk, L.; Huszno, B.; Gołkowski, F.; Przybylik-Mazurek, E.; et al. A programme of iodine supplementation using only iodised household salt is efficient—The case of Poland. Eur. J. Endocrinol. 2001, 144, 331–337. [Google Scholar] [CrossRef] [Green Version]
- Southern, A.P.; Jwayyed, S. Iodine Toxicity, StatPearls. Available online: https://www.statpearls.com/ArticleLibrary/viewarticle/40905 (accessed on 11 April 2021).
- Wu, T.; Liu, G.J.; Li, P.; Clar, C. Iodised salt for preventing iodine deficiency disorders. Cochrane Database Syst. Rev. 2002, CD003204. [Google Scholar] [CrossRef] [PubMed]
- Cao, X.; Ma, W.; Liu, L.; Xu, J.; Wang, H.; Li, X.; Wang, J.; Hang, J.; Wang, Z.; Gu, Y. Analysis of potassium iodate reduction in tissue homogenates using high performance liquid chromatography-inductively coupled plasma-mass spectrometry. J. Trace Elem. Med. Biol. 2015, 32, 1–6. [Google Scholar] [CrossRef]
- Milczarek, M.; Stepniak, J.; Lewinski, A.; Karbownik-Lewinska, M. Potassium iodide, but not potassium iodate, as a potential protective agent against oxidative damage to membrane lipids in porcine thyroid. Thyroid Res. 2013, 6, 10. [Google Scholar] [CrossRef] [Green Version]
- Karbownik-Lewinska, M.; Stepniak, J.; Milczarek, M.; Lewinski, A. Protective effect of KI in mtDNA in porcine thyroid: Comparison with KIO3 and nDNA. Eur. J. Nutr. 2015, 54, 319–323. [Google Scholar] [CrossRef] [Green Version]
- Iwan, P.; Stepniak, J.; Karbownik-Lewinska, M. Melatonin reduces high levels of lipid peroxidation induced by potassium iodate in porcine thyroid. Int. J. Vitam. Nutr. Res. 2019, 17, 1–7. [Google Scholar] [CrossRef]
- Iwan, P.; Karbownik-Lewinska, M. Indole-3-propionic acid reduces lipid peroxidation induced by potassium iodate in porcine thyroid. Interdiscip. Toxicol. 2020, 13, 101–105. [Google Scholar]
- Taurog, A.; Chaikoff, I.L.; Feller, D.D. The mechanism of iodine concentration by the thyroid gland: Its non-organic iodine-binding capacity in the normal and propylthiouracil-treated rat. J. Biol. Chem. 1947, 171, 189–201. [Google Scholar] [CrossRef]
- Taurog, A.; Tong, W.; Chaikoff, I.L. Non-thyroglobulin iodine of the thyroid gland II. Inorganic iodide. J. Biol. Chem. 1951, 191, 677–682. [Google Scholar] [CrossRef]
- Tiran, B.; Karpf, E.; Tiran, A.; Lax, S.; Langsteger, W.; Eber, O.; Lorenz, O. Iodine content of thyroid tissue in the Styrian population. Acta Med. Austriaca 1993, 20, 6–8. [Google Scholar] [PubMed]
- Trumbo, P.R. FDA regulations regarding iodine addition to foods and labeling of foods containing added iodine. Am. J. Clin. Nutr. 2016, 104, 864S–867S. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Reiter, R.J.; Mayo, J.C.; Tan, D.X.; Sainz, R.M.; Alatorre-Jimenez, M.; Qin, L. Melatonin as an antioxidant: Under promises but over delivers. J. Pineal Res. 2016, 61, 253–278. [Google Scholar] [CrossRef]
- Tan, D.X.; Manchester, L.C.; Esteban-Zubero, E.; Zhou, Z.; Reiter, R.J. Melatonin as a Potent and Inducible Endogenous Antioxidant: Synthesis and Metabolism. Molecules 2015, 16, 18886–18906. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tan, D.X.; Manchester, L.C.; Terron, M.P.; Flores, L.J.; Reiter, R.J. One molecule, many derivatives: A never-ending interaction of melatonin with reactive oxygen and nitrogen species? J. Pineal Res. 2007, 42, 28–42. [Google Scholar] [CrossRef]
- Galano, A.; Tan, D.X.; Reiter, R.J. On the free radical scavenging activities of melatonin’s metabolites, AFMK and AMK. J. Pineal Res. 2013, 54, 245–257. [Google Scholar] [CrossRef] [PubMed]
- Reiter, R.J.; Tan, D.X.; Galano, A. Melatonin: Exceeding expectations. Physiology 2014, 29, 325–333. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Karbownik, M.; Reiter, R.J. Melatonin protects against oxidative stress caused by delta-aminolevulinic acid: Implications for cancer reduction. Cancer Invest. 2002, 20, 276–286. [Google Scholar] [CrossRef]
- Karbownik, M.; Lewinski, A.; Reiter, R.J. Anticarcinogenic actions of melatonin which involve antioxidative processes: Comparison with other antioxidants. Int. J. Biochem. Cell Biol. 2001, 33, 735–753. [Google Scholar] [CrossRef]
- Bonmati-Carrion, M.A.; Arguelles-Prieto, R.; Martinez-Madrid, M.J.; Reiter, R.J.; Hardeland, R.; Rol, M.A.; Madrid, J.A. Protecting the melatonin rhythm through circadian healthy light exposure. Int. J. Mol. Sci. 2014, 15, 23448–23500. [Google Scholar] [CrossRef]
- Tordjman, S.; Chokron, S.; Delorme, R.; Charrier, A.; Bellissant, E.; Jaafari, N.; Fougerou, C. Melatonin: Pharmacology, Functions and Therapeutic Benefits. Curr. Neuropharmacol. 2017, 15, 434–443. [Google Scholar] [CrossRef]
- Pierce, M.; Linnebur, S.A.; Pearson, S.M.; Fixen, D.R. Optimal Melatonin Dose in Older Adults: A Clinical Review of the Literature. Sr. Care Pharm. 2019, 34, 419–431. [Google Scholar] [CrossRef] [PubMed]
- Madsen, B.K.; Zetner, D.; Møller, A.M.; Rosenberg, J. Melatonin for preoperative and postoperative anxiety in adults. Cochrane Database Syst. Rev. 2020, 8, 12. [Google Scholar] [CrossRef]
- Andersen, L.P.H.; Gögenur, I.; Rosenberg, J.; Reiter, R.J. The Safety of Melatonin in Humans. Clin. Drug Investig. 2015, 36, 169–175. [Google Scholar] [CrossRef]
- Poeggeler, B.; Pappolla, M.A.; Hardeland, R.; Rassoulpour, A.; Hodgkins, P.S.; Guidetti, P.; Schwarcz, R. Indole-3-propionate: A potent hydroxyl radical scavenger in rat brain. Brain Res. 1999, 815, 382–388. [Google Scholar] [CrossRef]
- Karbownik, M.; Stasiak, M.; Zasada, K.; Zygmunt, A.; Lewinski, A. Comparison of potential protective effects of melatonin, indole-3-propionic acid, and propylthiouracil against lipid peroxidation caused by potassium bromate in the thyroid gland. J. Cell Biochem. 2005, 95, 131–138. [Google Scholar] [CrossRef] [PubMed]
- Karbownik, M.; Stasiak, M.; Zygmunt, A.; Zasada, K.; Lewinski, A. Protective effects of melatonin and indole-3-propionic acid against lipid peroxidation, caused by potassium bromate in the rat kidney. Cell Biochem. Funct. 2006, 24, 483–489. [Google Scholar] [CrossRef] [PubMed]
- Karbownik, M.; Reiter, R.J.; Garcia, J.J.; Cabrera, J.; Burkhardt, S.; Osuna, C.; Lewinski, A. Indole-3-propionic acid, a melatonin-related molecule, protects hepatic microsomal membranes from iron-induced oxidative damage: Relevance to cancer reduction. J. Cell Biochem. 2001, 81, 507–513. [Google Scholar] [CrossRef]
- Karbownik, M.; Garcia, J.J.; Lewinski, A.; Reiter, R.J. Carcinogen-induced, free radical-mediated reduction in microsomal membrane fluidity: Reversal by indole-3-propionic acid. J. Bioenerg. Biomembr. 2001, 33, 73–78. [Google Scholar] [CrossRef]
- Bendheim, P.E.; Poeggeler, B.; Neria, E.; Ziv, V.; Pappolla, M.A.; Chain, D.G. Development of indole-3-propionic acid (OXIGON) for Alzheimer’s disease. J. Mol. Neurosci. 2002, 19, 213–217. [Google Scholar] [CrossRef] [PubMed]
- Karbownik, M.; Lewinski, A. Melatonin reduces Fenton reaction-induced lipid peroxidation in porcine thyroid tissue. J. Cell Biochem. 2003, 90, 806–811. [Google Scholar] [CrossRef] [PubMed]
- Karbownik, M.; Lewinski, A. The role of oxidative stress in physiological and pathological processes in the thyroid gland; possible involvement in pineal-thyroid interactions. Neuro. Endocrinol. Lett. 2003, 24, 293–303. [Google Scholar]
- Lewinski, A.; Karbownik, M. REVIEW. Melatonin and the thyroid gland. Neuro. Endocrinol. Lett. 2002, 23 (Suppl. 1), 73–78.–78. [Google Scholar]
- Bradford, M.M. A rapid and sensitive method for the quantitation of microgram quantities of protein utilizing the principle of protein-dye binding. Anal. Biochem. 1976, 7, 248–254. [Google Scholar] [CrossRef]
- FDA. Available online: https://www.accessdata.fda.gov/scripts/cdrh/cfdocs/cfcfr/CFRSearch.cfm?fr=184.1635 (accessed on 25 September 2020).
- Bürgi, H.; Schaffner, T.H.; Seiler, J.P. The toxicology of iodate: A review of the literature. Thyroid 2001, 11, 449–456. [Google Scholar] [CrossRef] [Green Version]
- IARC. Available online: https://monographs.iarc.fr/wp-content/uploads/2018/06/mono73-22.pdf (accessed on 25 September 2020).
- Toloza, F.J.K.; Motahari, H.; Maraka, S. Consequences of Severe Iodine Deficiency in Pregnancy: Evidence in Humans. Front. Endocrinol. 2020, 11, 409. [Google Scholar] [CrossRef]
- Zimmermann, M.B.; Jooste, P.L.; Pandav, C.S. Iodine-deficiency disorders. Lancet 2008, 372, 1251–1262. [Google Scholar] [CrossRef]
- Karbownik, M.; Reiter, R.J.; Garcia, J.J.; Tan, D.X. Melatonin reduces phenylhydrazine-induced oxidative damage to cellular membranes: Evidence for the involvement of iron. Int. J. Biochem. Cell Biol. 2000, 32, 1045–1054. [Google Scholar] [CrossRef]
- Karbownik, M.; Reiter, R.J.; Garcia, J.J.; Tan, D.X.; Qi, W.; Manchester, L.C. Melatonin reduces rat hepatic macromolecular damage due to oxidative stress caused by delta-aminolevulinic acid. Biochim. Biophys. Acta 2000, 18, 140–146. [Google Scholar] [CrossRef]
- Chyan, Y.J.; Poeggeler, B.; Omar, R.A.; Chain, D.G.; Frangione, B.; Ghiso, J.; Pappolla, M.A. Potent neuroprotective properties against the Alzheimer β-Amyloid by an endogenous melatonin-related indole structure, indole-3-propionic acid. J. Biol. Chem. 1999, 274, 21937–21942. [Google Scholar] [CrossRef] [Green Version]
- Candeias, L.P.; Folkes, L.K.; Porssa, M.; Parrick, J.; Wardman, P. Enhancement of Lipid Peroxidation by Indole-3-Acetic Acid and Derivatives: Substituent Effects. Free Radic Res. 1995, 23, 403–418. [Google Scholar] [CrossRef]
- Zetner, D.; Andersen, L.P.K.; Alder, R.; Jessen, M.L.; Tolstrup, A.; Rosenberg, J. Pharmacokinetics and Safety of Intravenous, Intravesical, Rectal, Transdermal, and Vaginal Melatonin in Healthy Female Volunteers: A Cross-Over Study. Pharmacology 2021, 106, 169–176. [Google Scholar] [CrossRef]
- Andersen, L.P.H.; Werner, M.U.; Rosenkilde, M.M.; Harpsøe, N.G.; Fuglsang, H.; Rosenberg, J.; Gögenur, I. Pharmacokinetics of oral and intravenous melatonin in healthy volunteers. BMC Pharmacol. Toxicol. 2016, 19, 8. [Google Scholar] [CrossRef] [Green Version]
- Baltatu, O.C.; Senar, S.; Campos, L.A.; Cipolla-Neto, J. Cardioprotective Melatonin: Translating from Proof-of-Concept Studies to Therapeutic Use. Int. J. Mol. Sci. 2019, 20, 4342. [Google Scholar] [CrossRef] [Green Version]
- Lin, L.; Huang, Q.X.; Yang, S.S.; Chu, J.; Wang, J.Z.; Tian, Q. Melatonin in Alzheimer’s disease. Int. J. Mol. Sci. 2013, 14, 14575–14593. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Romero, A.; Ramos, E.; López-Muñoz, F.; Gil-Martín, E.; Escames, G.; Reiter, R.J. Coronavirus Disease 2019 (COVID-19) and Its Neuroinvasive Capacity: Is It Time for Melatonin? Cell Mol. Neurobiol. 2020, 9, 1–12. [Google Scholar] [CrossRef]
- Acuña-Castroviejo, D.; Escames, G.; Figueira, J.C.; de la Oliva, P.; Borobia, A.M.; Acuña-Fernández, C. Clinical trial to test the efficacy of melatonin in COVID-19. J. Pineal Res. 2020, 69, e12683. [Google Scholar] [CrossRef]
- Reiter, R.J.; Abreu-Gonzalez, P.; Marik, P.E.; Dominguez-Rodriguez, A. Therapeutic Algorithm for Use of Melatonin in Patients With COVID-19. Front. Med. 2020, 7, 226. [Google Scholar] [CrossRef]
- Zhang, R.; Wang, X.; Ni, L.; Di, X.; Ma, B.; Niu, S.; Liu, C.; Reiter, R.J. COVID-19: Melatonin as a potential adjuvant treatment. Life Sci. 2020, 250, 117583. [Google Scholar] [CrossRef] [PubMed]
- Li, X.; Cao, X.; Li, J.; Xu, J.; Ma, W.; Wang, H.; Wang, J.; Zhang, Y. Effects of high potassium iodate intake on iodine metabolism and antioxidant capacity in rats. J. Trace Elem. Med. Biol. 2020, 126575. [Google Scholar] [CrossRef] [PubMed]


Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Iwan, P.; Stepniak, J.; Karbownik-Lewinska, M. Cumulative Protective Effect of Melatonin and Indole-3-Propionic Acid against KIO3—Induced Lipid Peroxidation in Porcine Thyroid. Toxics 2021, 9, 89. https://0-doi-org.brum.beds.ac.uk/10.3390/toxics9050089
Iwan P, Stepniak J, Karbownik-Lewinska M. Cumulative Protective Effect of Melatonin and Indole-3-Propionic Acid against KIO3—Induced Lipid Peroxidation in Porcine Thyroid. Toxics. 2021; 9(5):89. https://0-doi-org.brum.beds.ac.uk/10.3390/toxics9050089
Chicago/Turabian StyleIwan, Paulina, Jan Stepniak, and Malgorzata Karbownik-Lewinska. 2021. "Cumulative Protective Effect of Melatonin and Indole-3-Propionic Acid against KIO3—Induced Lipid Peroxidation in Porcine Thyroid" Toxics 9, no. 5: 89. https://0-doi-org.brum.beds.ac.uk/10.3390/toxics9050089