
adipoSIGHT in Therapeutic Response: Consequences in Osteosarcoma Treatment

 , ,
, ,  , ,
, ,
Abstract
:1. Introduction
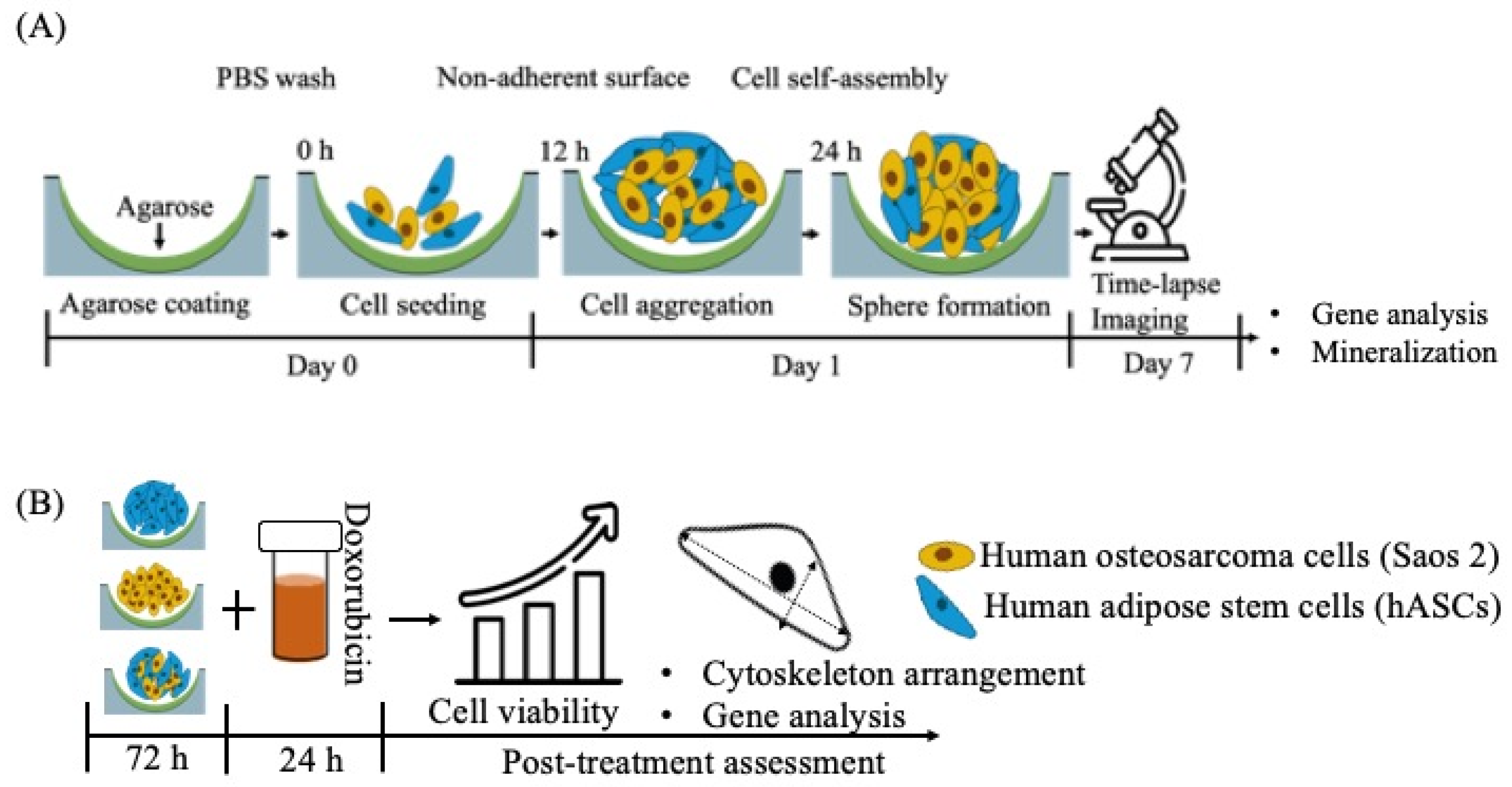
2. Materials and Methods
2.1. Preparation of Agarose-Coated Wells
2.2. Cell Isolation and Culture
2.3. 3D Time-Lapse Confocal Microscopy
2.4. Calcium Depletion Test
2.5. Mineralization Assay
2.6. Cytotoxicity Assay
2.7. Actin Staining
2.8. Gene Expression Analysis
2.9. Statistical Analysis
3. Results
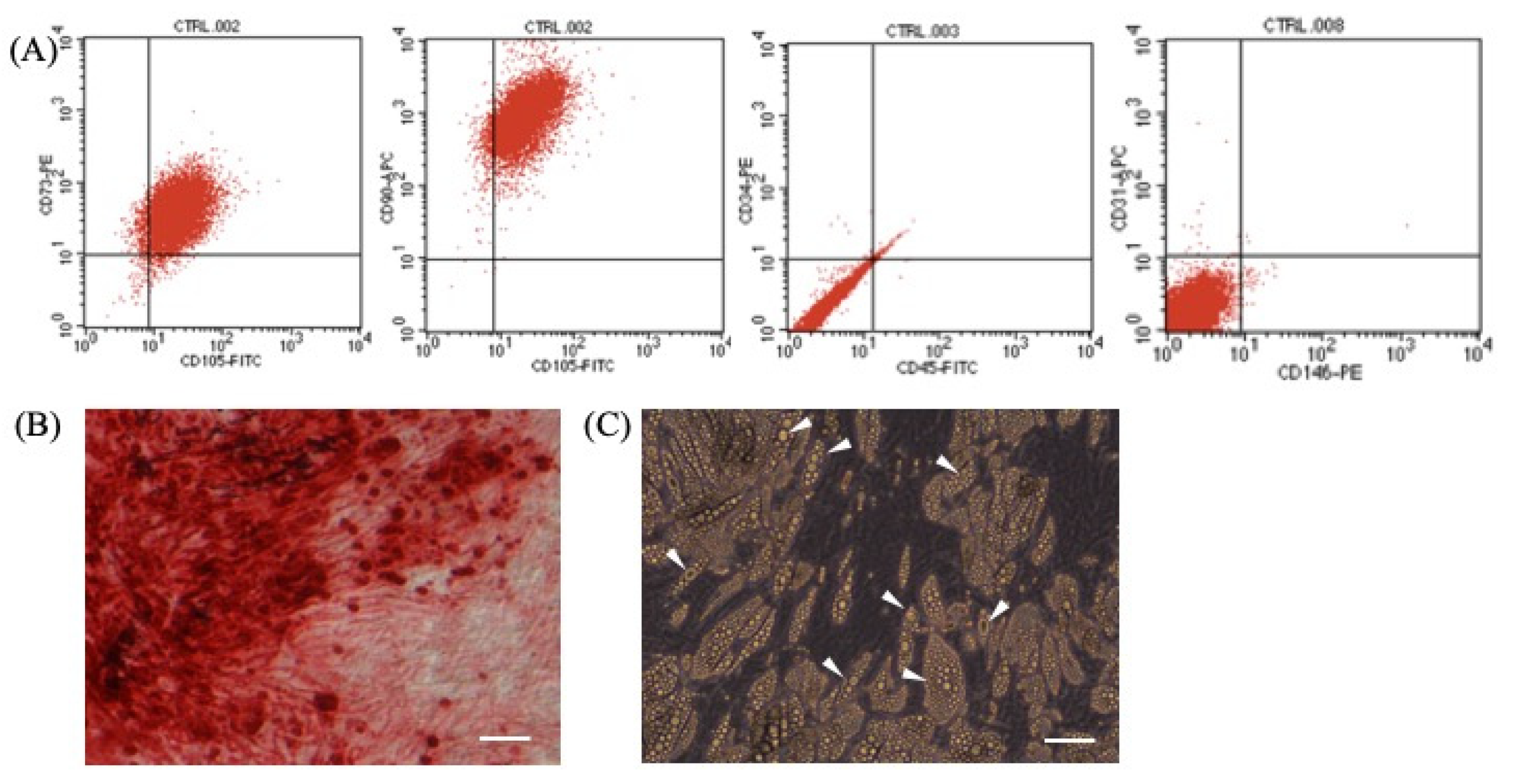
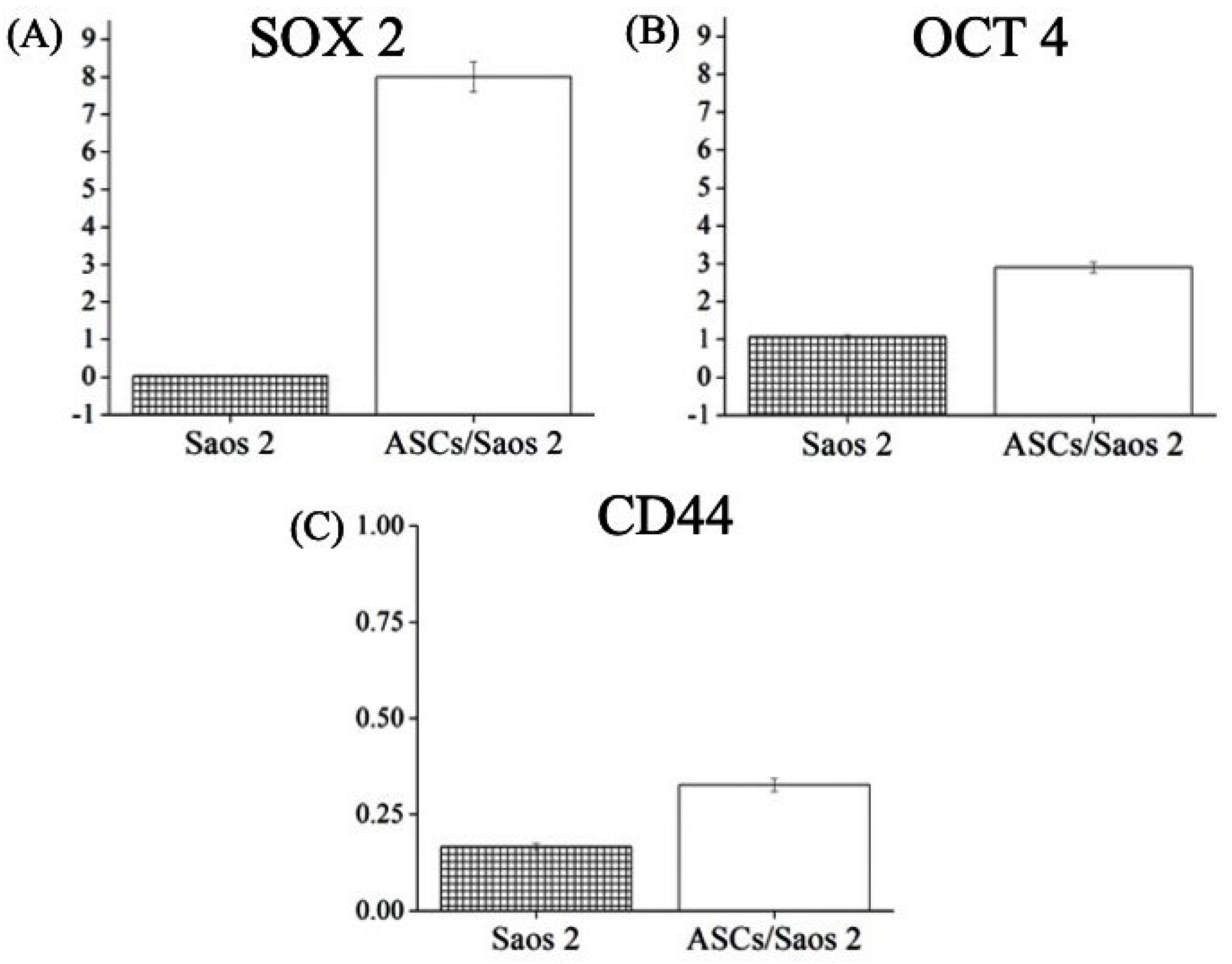
3.1. Determination of Stemness of ASCs
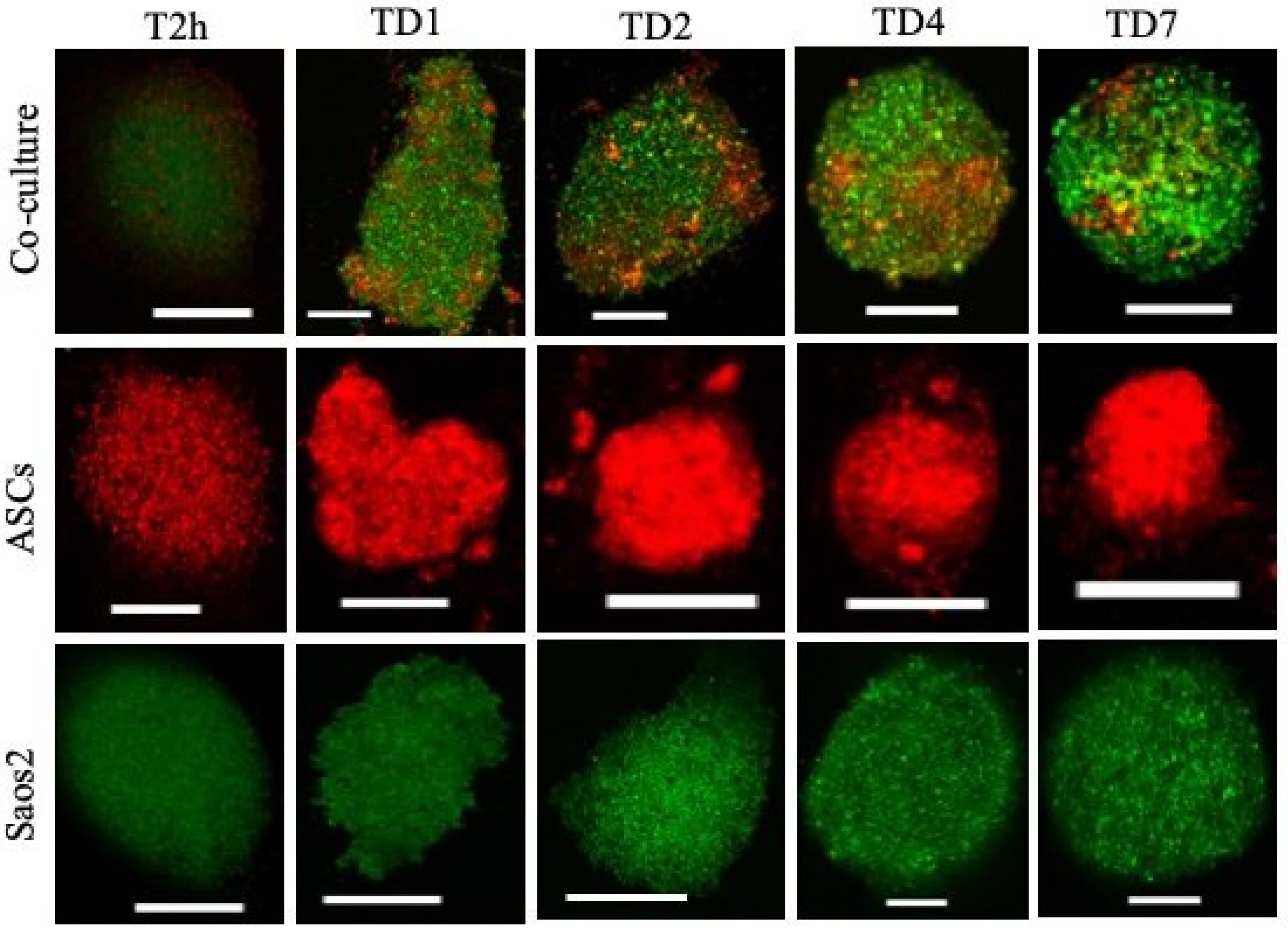
3.2. Spheroid Formation Dynamics
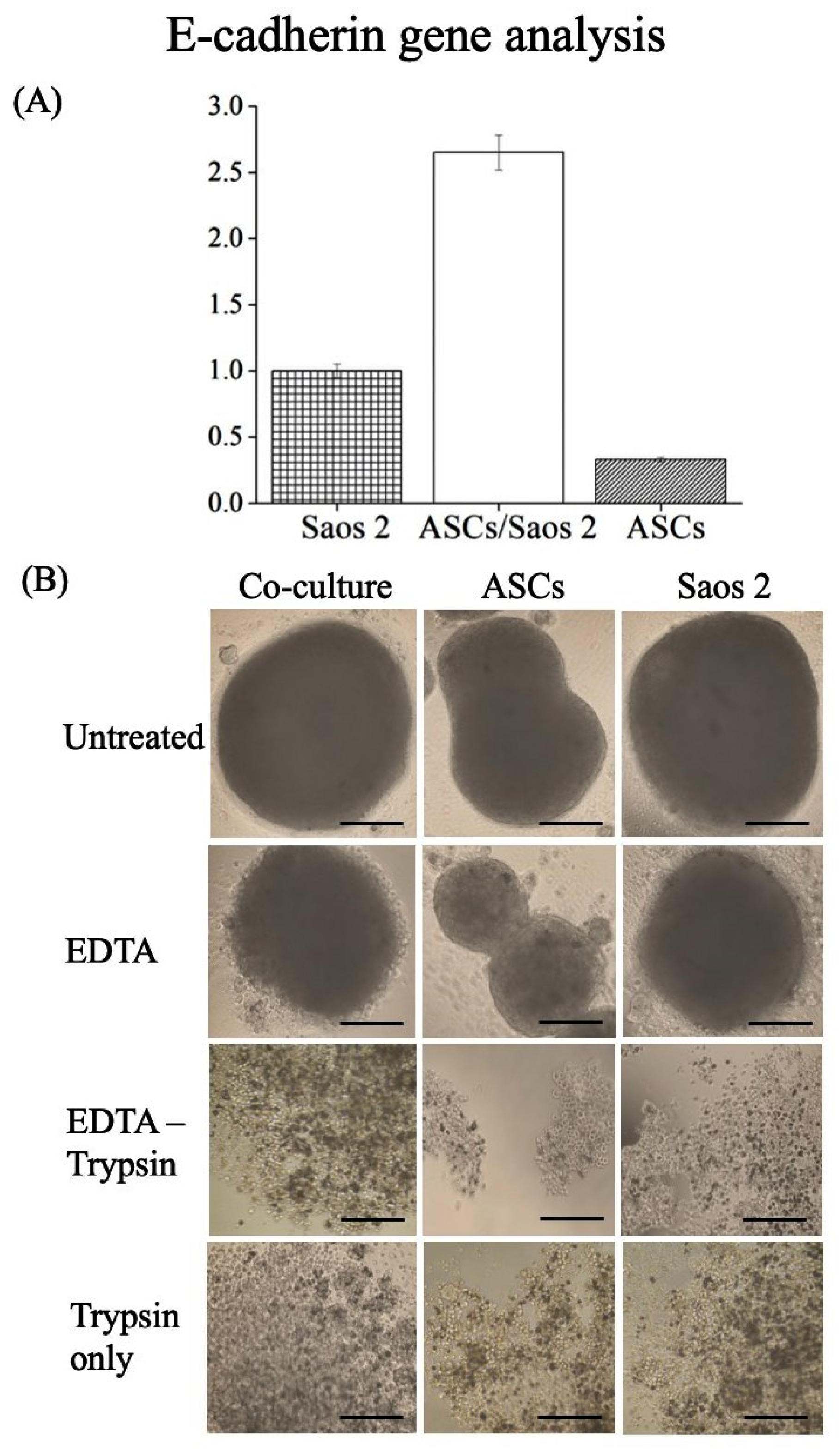
3.3. E-Cadherin Expression and Cell–Cell Interactions
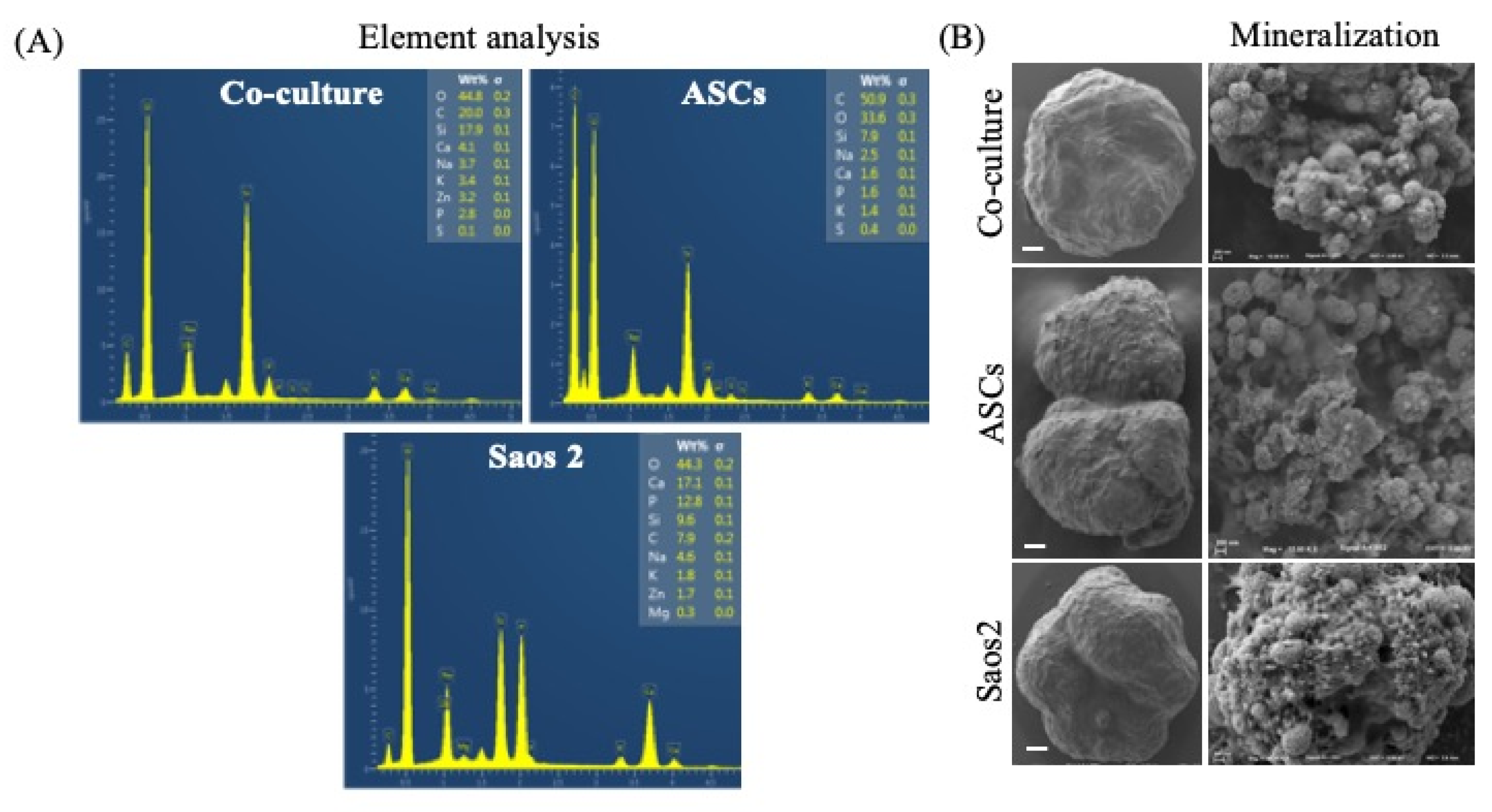
3.4. Mineralization
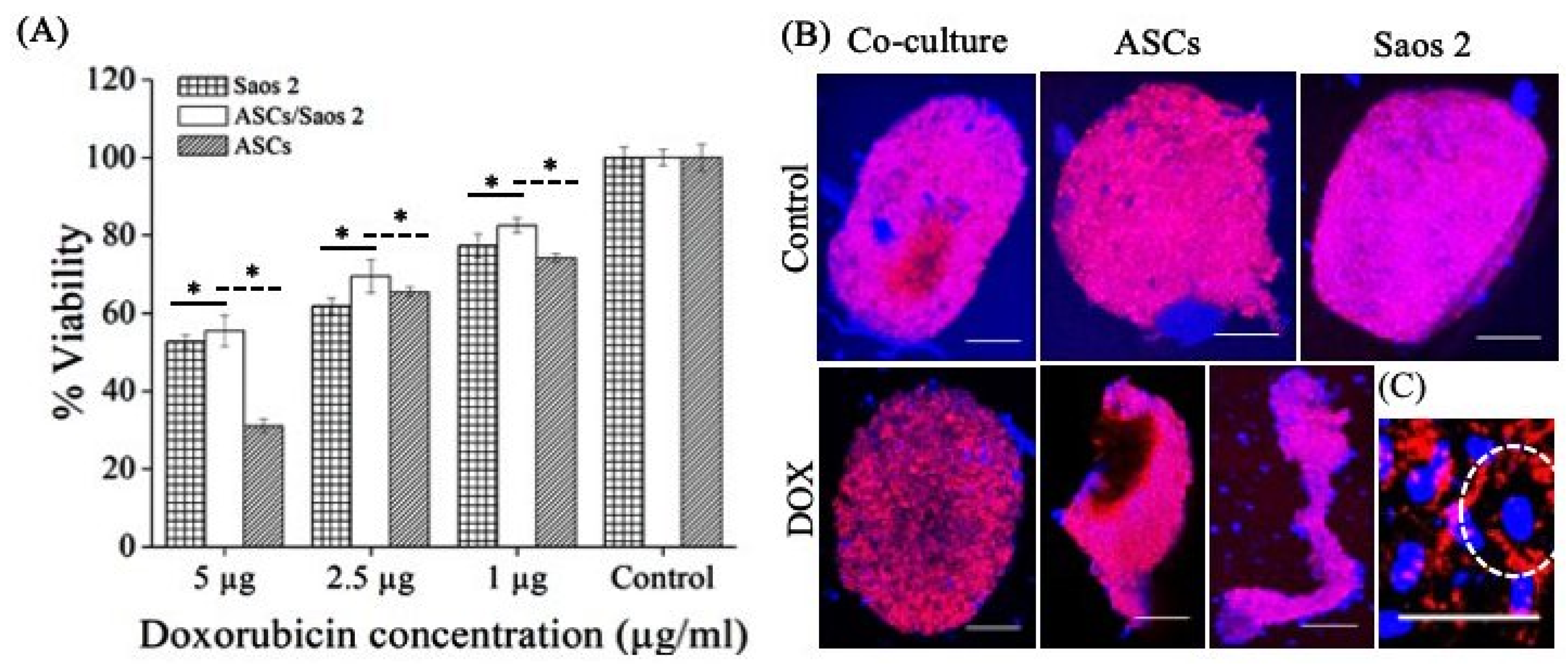
3.5. Drug Resistance and Cytoskeleton Arrangement
3.6. Differential Expression of Genes in Homo- and Heterotypic Tumorspheres
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Wang, Y.; Chu, Y.; Yue, B.; Ma, X.; Zhang, G.; Xiang, H.; Liu, Y.; Wang, T.; Wu, X.; Chen, B. Adipose-derived mesenchymal stem cells promote osteosarcoma proliferation and metastasis by activating the STAT3 pathway. Oncotarget 2017, 8, 23803–23816. [Google Scholar] [CrossRef] [PubMed]
- Anderson, M.E. Update on survival in osteosarcoma. Orthop. Clin. N. Am. 2016, 47, 283–292. [Google Scholar] [CrossRef]
- Huang, J.; Bi, W.; Han, G.; Jia, J.; Xu, M.; Wang, W. The multidisciplinary treatment of osteosarcoma of the proximal tibia: A retrospective study. BMC Musculoskelet Disord. 2018, 19, 315. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.; Chu, Y.; Li, K.; Zhang, G.; Guo, Z.; Wu, X.; Qiu, C.; Li, Y.; Wan, X.; Sui, J.; et al. Exosomes secreted by adipose-derived mesenchymal stem cells foster metastasis and osteosarcoma proliferation by increasing COLGALT2 expression. Front. Cell Dev. Biol. 2020, 8, 353. [Google Scholar] [CrossRef]
- Kundu, B.; Bastos, A.R.F.; Brancato, V.; Cerqueira, M.T.; Oliveira, J.M.; Correlo, V.M.; Reis, R.L.; Kundu, S.C. Mechanical property of hydrogels and the presence of adipose stem cells in tumor stroma affect spheroid formation in the 3D osteosarcoma model. ACS Appl. Mater. Interfaces 2019, 11, 14548–14559. [Google Scholar] [CrossRef]
- O’Halloran, N.; Khan, S.; Gilligan, K.; Dwyer, R.; Kerin, M.; Lowery, A. Oncological risk in autologous stem cell donation for novel tissue-engineering approaches to postmastectomy breast regeneration. Breast Cancer 2019, 13, 1. [Google Scholar] [CrossRef]
- Cantini, G.; Di Franco, A.; Mannelli, M.; Scimè, A.; Maggi, M.; Luconi, M. The role of metabolic changes in shaping the fate of cancer-associated adipose stem cells. Front. Cell Dev. Biol. 2020, 8, 332. [Google Scholar] [CrossRef] [PubMed]
- Langenbach, F.; Berr, K.; Naujoks, C.; Hassel, A.; Hentschel, M.; Depprich, R.; Kubler, N.R.; Meyer, U.; Wiesmann, H.-P.; Kögler, G.; et al. Generation and differentiation of microtissues from multipotent precursor cells for use in tissue engineering. Nat. Protoc. 2011, 6, 1726–1735. [Google Scholar] [CrossRef]
- Kundu, B.; Brancato, V.; Oliveira, J.M.; Correlo, V.M.; Reis, R.L.; Kundu, S.C. Silk fibroin promotes mineralization of gellan gum hydrogels. Int. J. Biol Macromol. 2019, 153, 1328. [Google Scholar] [CrossRef]
- Bucan, V.; Fliess, M.; Schnabel, R.; Peck, C.T.; Vaslaitis, D.; Fülbier, A.; Reimers, K.; Strauss, S.; Vogt, P.M.; Radtke, C. In vitro enhancement and functional characterization of neurite outgrowth by undifferentiated adipose-derived stem cells. Int. J. Mol. Med. 2019, 43, 593–602. [Google Scholar] [CrossRef] [Green Version]
- Xu, S.; Yang, Y.; Dong, L.; Qiu, W.; Yang, L.; Wang, X.; Liu, L. Construction and characteristics of an E-cadherin-related three-dimensional suspension growth model of ovarian cancer. Sci. Rep. 2014, 4, 5646. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mizerska-Kowalska, M.; Sławińska-Brych, A.; Kaławaj, K.; Żurek, A.; Pawińska, B.; Rzeski, W.; Zdzisińska, B. Betulin promotes differentiation of human osteoblasts in vitro and exerts an osteoinductive effect on the hFOB 1.19 cell line through activation of JNK, ERK1/2, and mTOR kinases. Molecules 2019, 24, 2637. [Google Scholar] [CrossRef] [Green Version]
- Pait, M.; Kundu, B.; Kundu, S.C.; Ray, D. Copper (II) complexes of biologically relevant piperazine based ligand: Synthesis, crystal structure, protein binding and evaluation of anti-cancerous therapeutic potential. Inorg. Chim. Acta 2014, 418, 30–41. [Google Scholar] [CrossRef]
- Livak, K.J.; Schmittgen, T.D. Analysis of relative gene expression data using real-time quantitative PCR and the 2−ΔΔCT method. Methods 2001, 25, 402–408. [Google Scholar] [CrossRef] [PubMed]
- Bonuccelli, G.; Avnet, S.; Grisendi, G.; Salerno, M.; Granchi, D.; Dominici, M.; Kusuzaki, K.; Baldini, N. Role of mesenchymal stem cells in osteosarcoma and metabolic reprogramming of tumor cells. Oncotarget 2014, 5, 7575–7588. [Google Scholar] [CrossRef]
- Mazini, L.; Ezzoubi, M.; Malka, G. Overview of current adipose-derived stem cell (ADSCs) processing involved in therapeutic advancements: Flow chart and regulation updates before and after COVID-19. Stem. Cell Res. Ther. 2021, 12, 1–17. [Google Scholar] [CrossRef]
- Smyrek, I.; Mathew, B.; Fischer, S.C.; Lissek, S.M.; Becker, S.; Stelzer, E.H.K. E-cadherin, actin, microtubules and FAK dominate different spheroid formation phases and important elements of tissue integrity. Biol. Open 2019, 8, bio037051. [Google Scholar] [CrossRef] [Green Version]
- West, G.W.; Weichselbaum, R.; Little, J.B. Limited penetration of methotrexate into human osteosarcoma spheroids as a proposed model for solid tumor resistance to adjuvant chemotherapy. Cancer Res. 1980, 40, 3665–3668. [Google Scholar] [PubMed]
- Kerbel, R.S.; Rak, J.; Kobayashi, H.; Man, M.S.; St Croix, B.; Graham, C.H. Multicellular resistance: A new paradigm to explain aspects of acquired drug resistance of solid tumors. Cold Spring Harb. Symp. Quant. Biol. 1994, 59, 661–672. [Google Scholar] [CrossRef] [PubMed]
- Maines-Bandiera, S.L.; Auersperg, N. Increased E-cadherin expression in ovarian surface epithelium: An early step in metaplasia and dysplasia. Int. J. Gynecol. Pathol. 1997, 16, 250–255. [Google Scholar] [CrossRef]
- Wei, L.; Surma, M.; Gough, G.; Shi, S.; Lambert-Cheatham, N.; Chang, J.; Shi, J. Dissecting the mechanisms of doxorubicin and oxidative stress-induced cytotoxicity: The involvement of actin cytoskeleton and ROCK1. PLoS ONE 2015, 10, e0131763. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ullah, M.; Akbar, A.; Ng, N.N.; Concepcion, W.; Thakor, A.S. Mesenchymal stem cells confer chemoresistance in breast cancer via a CD9 dependent mechanism. Oncotarget 2019, 10, 3435. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Basu-Roy, U.; Seo, E.; Ramanathapuram, L.; Rapp, T.B.; Perry, J.A.; Orkin, S.H.; Mansukhani, A.; Basilico, C. Sox2 maintains self-renewal of tumor-initiating cells in osteosarcomas. Oncogene 2012, 31, 2270–2282. [Google Scholar] [CrossRef] [Green Version]
- Decarvalho, A.C.; Nelson, K.; Lemke, N.; Lehman, N.L.; Arbab, A.S.; Kalkanis, S.; Mikkelsen, T. Gliosarcoma stem cells undergo glial and mesenchymal differentiation in vivo. Stem. Cells 2010, 28, 181–190. [Google Scholar] [CrossRef] [Green Version]
- He, J.; Shi, J.; Zhang, K.; Xue, J.; Li, J.; Yang, J.; Chen, J.; Wei, J.; Hong Ren, H.; Liu, X. Sox2 inhibits Wnt-β-catenin signaling and metastatic potency of cisplatin-resistant lung adenocarcinoma cells. Mol. Med. Rep. 2017, 15, 1693–1701. [Google Scholar] [CrossRef] [Green Version]
- Mayr, L.; Pirker, C.; Lötsch, D.; Schoonhoven, S.V.; Windhager, R.; Englinger, B.; Berger, W.; Kubista, B. CD44 drives aggressiveness and chemoresistance of a metastatic human osteosarcoma xenograft model. Oncotarget 2017, 8, 114095. [Google Scholar] [CrossRef] [PubMed]
- Steiner, E.; Holzmann, K.; Pirker, C.; Elbling, L.; Micksche, M.; Sutterluty, H.; Berger, W. The major vault protein is responsive to and interferes with interferon-gamma- mediated STAT1 signals. J. Cell Sci. 2006, 119, 459–469. [Google Scholar] [CrossRef] [Green Version]
- Karousou, E.; Misra, S.; Ghatak, S.; Dobra, K.; Gotte, M.; Vigetti, D.; Passi, A.; Karamanos, N.K.; Skandalis, S.S. Roles and targeting of the HAS/hyaluronan/ CD44 molecular system in cancer. Matrix Biol. 2017, 59, 3–22. [Google Scholar] [CrossRef]
- Rauwolf, M.; Pemmer, B.; Roschger, A.; Turyanskaya, A.; Smolek, S.; Maderitsch, A.; Hischenhuber, P.; Foelser, M.; Simon, R.; Lang, S.; et al. Increased zinc accumulation in mineralized osteosarcoma tissue measured by confocal synchrotron radiation micro X-ray fluorescence analysis. X-Ray Spectrom. 2017, 46, 56–62. [Google Scholar] [CrossRef]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Genes | Primer Sequences | Amplicon (bp) | Annealing T (°C) | ID |
|---|---|---|---|---|
| OCT4 | Fw 5′–TACTCCTCGGTCCCTTTCC–3′ | 167 | 60 | NM_002701.6 |
| Rw 5′–CAAAACCCTGGCACAAACT–3′ | ||||
| Sox-2 | Fw 5′–ACACCAATCCCATCCACACT–3′ | 224 | 60 | NM_003106.4 |
| Rw 5′–GCAAACTCCCTGCAAAGCTC–3′ | ||||
| CD44 | Fw 5′–CGCAGCCTGGGGACTCTG–3′, | 117 | 60 | NM_001202555.2 |
| Rev 5′–CGAGAGATGCTGTAGCGACCA–3′ | ||||
| E-Cadherin | Fw 5′–AAGAAGCTGGCTGACATGTAC–3′ | 105 | 58.8 | NM_001317185.2 |
| Rv 5′–ACCAGCAACGTGATTTCTGCAT–3′ | ||||
| GAPDH | Fw 5′–AGCCTCAAGATCATCAGCAA–3′ | 101 | - | NM_001289745.2 |
| Rv 5′–GTCATGAGTCCTTCCACGAT–3′ |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kundu, B.; Brancato, V.; Oliveira, J.; Correlo, V.M.; Reis, R.L.; Kundu, S.C. adipoSIGHT in Therapeutic Response: Consequences in Osteosarcoma Treatment. Bioengineering 2021, 8, 83. https://0-doi-org.brum.beds.ac.uk/10.3390/bioengineering8060083
Kundu B, Brancato V, Oliveira J, Correlo VM, Reis RL, Kundu SC. adipoSIGHT in Therapeutic Response: Consequences in Osteosarcoma Treatment. Bioengineering. 2021; 8(6):83. https://0-doi-org.brum.beds.ac.uk/10.3390/bioengineering8060083
Chicago/Turabian StyleKundu, Banani, Virginia Brancato, Joaquim Oliveira, Vitor M. Correlo, Rui L. Reis, and Subhas C. Kundu. 2021. "adipoSIGHT in Therapeutic Response: Consequences in Osteosarcoma Treatment" Bioengineering 8, no. 6: 83. https://0-doi-org.brum.beds.ac.uk/10.3390/bioengineering8060083