
Identification of Polymorphisms of the CSN2 Gene Encoding β-Casein in Greek Local Breeds of Cattle

 , , and
, , and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Experimental Animals and Farms
2.1.1. Institutional Animal Care and Use Committee (IACUC)
2.1.2. Selection and Preparation of Control Samples
2.1.3. Selection and Preparation of Blood Samples
2.1.4. Genomic DNA Extraction
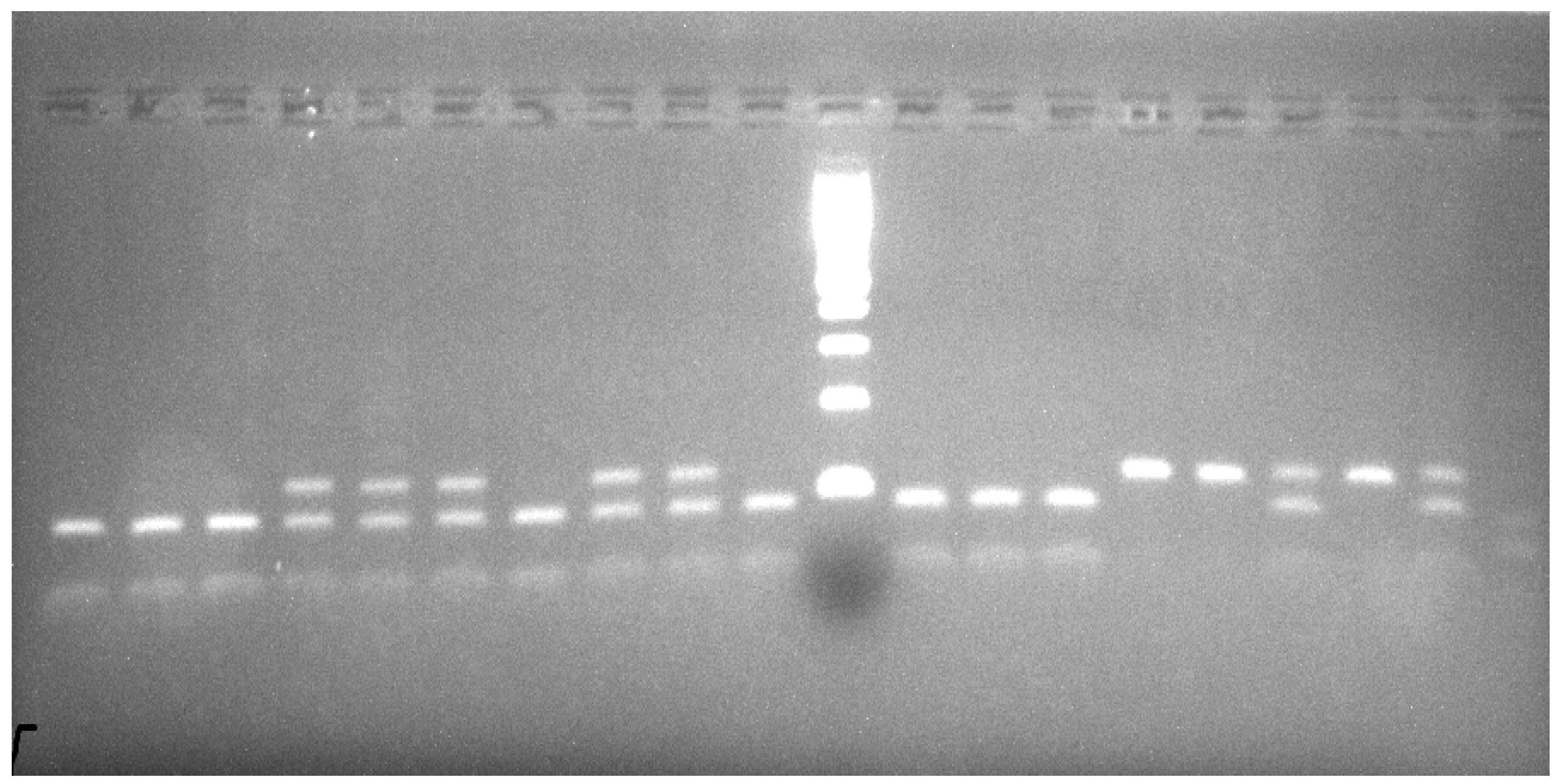
2.1.5. PCR Amplification and Restriction Fragment Length Polymorphism Detection I
2.1.6. PCR Amplification and Restriction Fragment Length Polymorphism Detection II
2.2. Statistical Analysis
3. Results
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Monaci, L.; Anklam, A.; Tregoat, V. Milk allergens, their characteristics and their detection in food: A review. Aust. J. Dairy Technol. 2007, 62, 62–71. [Google Scholar] [CrossRef]
- Miluchová, M.; Gábor, M.; Trakovická, A. Analysis of Beta-Casein Gene (CSN2) Polymorphism in Different Breeds of Cattle. Sci. Pap. Anim. Sci. Biotechnol. 2014, 47, 56–59. [Google Scholar]
- Massella, E.; Piva, S.; Giacometti, F.; Liuzzo, G.; Zambrini, A.V.; Serraino, A. Evaluation of bovine beta casein polymorphism in two dairy farms located in northern Italy. Ital. J. Food Saf. 2017, 6, 131–133. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kanwar, J.R.; Kanwar, R.K.; Sun, X.; Punj, V.; Matta, H.; Morley, S.M.; Parrat, A.; Puri, M.; Sehgal, R. Molecular and Biotechnological Advances in Milk Proteins in Relation to Human Health. Curr. Protein Pept. Sci. 2009, 10, 308–338. [Google Scholar] [CrossRef] [PubMed]
- Sodhi, M.; Kataria, R.S.; Niranjan, S.K.; Verma, P.; Swami, S.K.; Mukesh, M. Sequence 38 Characterisation and Genotyping of Allelic Variants of Beta Casein Gene Establishes Native Cattle of Ladakh to be a Natural Resource for A2 Milk. Def. Lif. Sci. J. 2018, 3, 177. [Google Scholar] [CrossRef]
- Miluchová, M.; Trakovická, A.; Gábor, M. Analysis Polymorphism of Beta Casein of Slovak Pinzgau Cattle by Pcr-Rflp for Allels A1 and A2. Sci. Pap. Anim. Sci. Biotechnol. 2009, 42, 288–292. [Google Scholar]
- Kumar, A.; Singh, R.V.; Chauhan, A.; Ilayakumar, K.; Kumar, S.; Kumar, A.; Sonwane, A.; Kumar, S.; Panigrahi, M.; Bhushan, B. Genetic association analysis reveals significant effect of β-casein A1/A2 loci on production & reproduction traits in Frieswal crossbred cows. Biol. Rhythm Res. 2019, 51, 1259–1272. [Google Scholar] [CrossRef]
- Woodford, K.B. A2 milk, farmer decisions, and risk management. In Proceedings of the 16th International Farm Management Congress: Peer Reviewed Papers; International Farm Management Association: Bristol, UK, 2007; pp. 641–648. Available online: http://researcharchive.lincoln.ac.nz/dspace/handle/10182/417 (accessed on 25 June 2021).
- Georgoudis, A.; Ligda, C.; Karkavelia, E.; Kotsaftiki, A.; Mizeli, C. Autochthonous Farm Animal Breeds of Greece. Greek Focal Point Manag. Anim. Genet. Resour. 2011, 49, 107. [Google Scholar]
- Bizelis, I. Current Situation of Autochthonous Breeds in Greece. 2015. Available online: https://www.save-foundation.net/images/konferenzen/2015/Iosif_Bizelis_Amaltheia.pdf (accessed on 2 July 2021).
- Saran, M.; Gurao, A.; Joshi, R.K.; Kashyap, S.K. Prevalence of the BETA case in variants among THARPARKAR, Rathi, sahiwal, Kankrej and cross breed and its influence under selective pressure. Int. J. Curr. Microbiol. Appl. Sci. 2019, 8, 1842–1848. [Google Scholar] [CrossRef]
- Rubino, G.; Pieragostini, E.; Caroli, A.; Petazzi, F. Quadro ematologico e proteico quali indici di diversità tra i bovini allevati in puglia: La razza podolica a confronto con le razze cosmopolite da latte. In Taurus Speciale; Anno XIX—Numero 6—Novembre/Dicembre 2007 ANABIC; Associazione Nazionale Allevatori Bovini Italiani da Carne: Perugia, Italy, 2007. [Google Scholar]
- Karatosidi, D.; Marsico, G.; Ligda, C.; Tarricone, S. Assessment of the meat quality of Italian Podolian and Greek Katerini cattle. Anim. Genet. Resour. 2012, 53, 141–146. [Google Scholar] [CrossRef]
- Georgoudis, A.G.; Ligd, C.; Boyazoglu, J. Population characteristics of water buffaloes in Greek wetlands. Anim. Genet. Resour. 1994, 14, 83–95. [Google Scholar] [CrossRef]
- Georgoudis, A.G.; Papanastasis, V.P.; Boyazoglu, J.G. Use of water buffalo for the environmental conservation of waterland-Review. Asian-Australas. J. Anim. Sci. 1999, 12, 1324–1331. [Google Scholar] [CrossRef]
- Groeneveld, L.F.; Lenstra, J.A.; Eding, H.; Toro, M.A.; Scherf, B.; Pilling, D.; Negrini, R.; Finlay, E.K.; Jianlin, H.; Groeneveld, E.; et al. Genetic diversity in farm animals—A review. Anim. Genet. 2010, 41, 6–31. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Flori, L.; Moazami-Goudarzi, K.; Alary, V.; Araba, A.; Boujenane, I.; Boushaba, N.; Casabianca, F.; Casu, S.; Ciampolini, R.; Coeur D’Acier, A.; et al. A genomic map of climate adaptation in Mediterranean cattle breeds. Mol. Ecol. 2019, 28, 1009–1029. [Google Scholar] [CrossRef] [PubMed]
- Çardak, A.D. Effects of genetic variants in milk protein on yield and composition of milk from Holstein-Friesian and Simmentaler cows. S. Afr. J. Anim. Sci. 2005, 35, 41–47. [Google Scholar] [CrossRef] [Green Version]
- Lin, C.Y.; McAllister, A.J.; Ng-Kwai-Hanh, K.F.; Hayes, J.F. Effects of milk protein loci on first lactation production in dairy cattle. J. Dairy Sci. 1986, 69, 704–712. [Google Scholar] [CrossRef]
- Ivanković, A.; Pećina, M.; Ramlja, J.; PašićGenetic, V. Ppolymorphism and effect on milk production of CSN2 gene in conventional and local cattle breeds in Croatia. Mljekarstvo 2021, 71, 3–12. [Google Scholar] [CrossRef]


{kind=link}
| Breeds | Total No. of Samples | Genotype Frequency | Allele Frequency | |||
|---|---|---|---|---|---|---|
| A1A1 | A1A2 | A2A2 | A1 | A2 | ||
| Holstein–Friesian | 780 | (n = 26) 3.3% | (n = 347) 44.5% | (n = 407) 52.2% | 25.6% | 74.4% |
| Vrachykeratiki | 46 | (n = 0) 0% | (n = 28) 60.9% | (n = 18) 39.1% | 30.4% | 58.7% |
| Katerinis | 20 | (n = 0) 0% | (n = 0) 0% | (n = 20) 100% | 0% | 100% |
| Sykias | 20 | (n = 0) 0% | (n = 0) 0% | (n = 20) 100% | 0% | 100% |
| Greek buffalo | 14 | (n = 0) 0% | (n = 0) 0% | (n = 14) 100% | 0% | 100% |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Antonopoulos, D.; Vougiouklaki, D.; Laliotis, G.P.; Tsironi, T.; Valasi, I.; Chatzilazarou, A.; Halvatsiotis, P.; Houhoula, D. Identification of Polymorphisms of the CSN2 Gene Encoding β-Casein in Greek Local Breeds of Cattle. Vet. Sci. 2021, 8, 257. https://0-doi-org.brum.beds.ac.uk/10.3390/vetsci8110257
Antonopoulos D, Vougiouklaki D, Laliotis GP, Tsironi T, Valasi I, Chatzilazarou A, Halvatsiotis P, Houhoula D. Identification of Polymorphisms of the CSN2 Gene Encoding β-Casein in Greek Local Breeds of Cattle. Veterinary Sciences. 2021; 8(11):257. https://0-doi-org.brum.beds.ac.uk/10.3390/vetsci8110257
Chicago/Turabian StyleAntonopoulos, Dionysios, Despina Vougiouklaki, George P. Laliotis, Theofania Tsironi, Irene Valasi, Archodoula Chatzilazarou, Panagiotis Halvatsiotis, and Dimitra Houhoula. 2021. "Identification of Polymorphisms of the CSN2 Gene Encoding β-Casein in Greek Local Breeds of Cattle" Veterinary Sciences 8, no. 11: 257. https://0-doi-org.brum.beds.ac.uk/10.3390/vetsci8110257