
Flow Cytometric Analysis of the Cytotoxic T-Cell Recall Response to Theileria parva in Cattle Following Vaccination by the Infection and Treatment Method

Abstract
:1. Introduction
2. Materials and Methods
2.1. Animals and Cell Lines
2.2. Generation of T. parva-Specific Effector T Cells
2.3. Flow Cytometric Cytotoxicity Assay
2.4. Data Analysis
3. Results
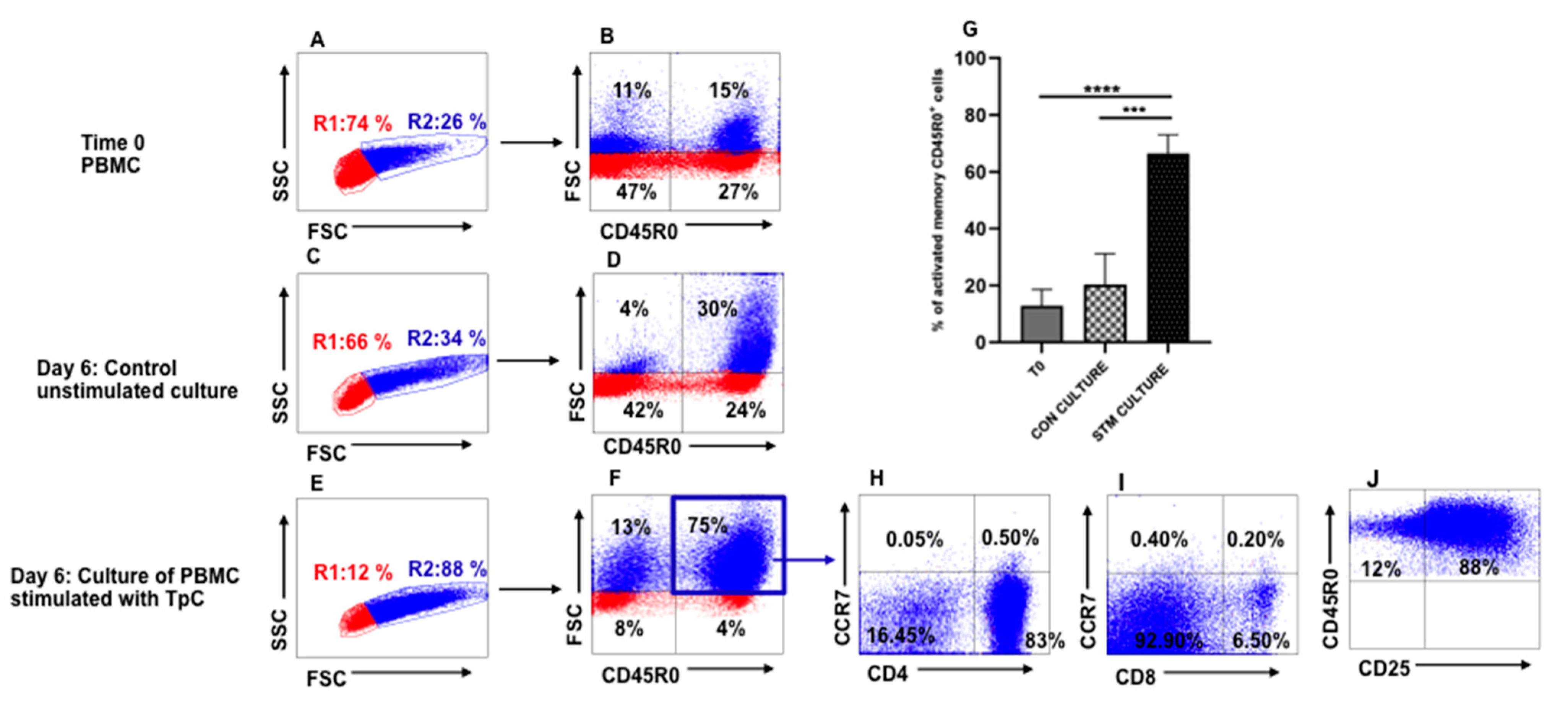
3.1. Recall Response to Stimulation of PBMC from T. parva-Immune Steers with Autologous T. parva-Infected Lymphocytes
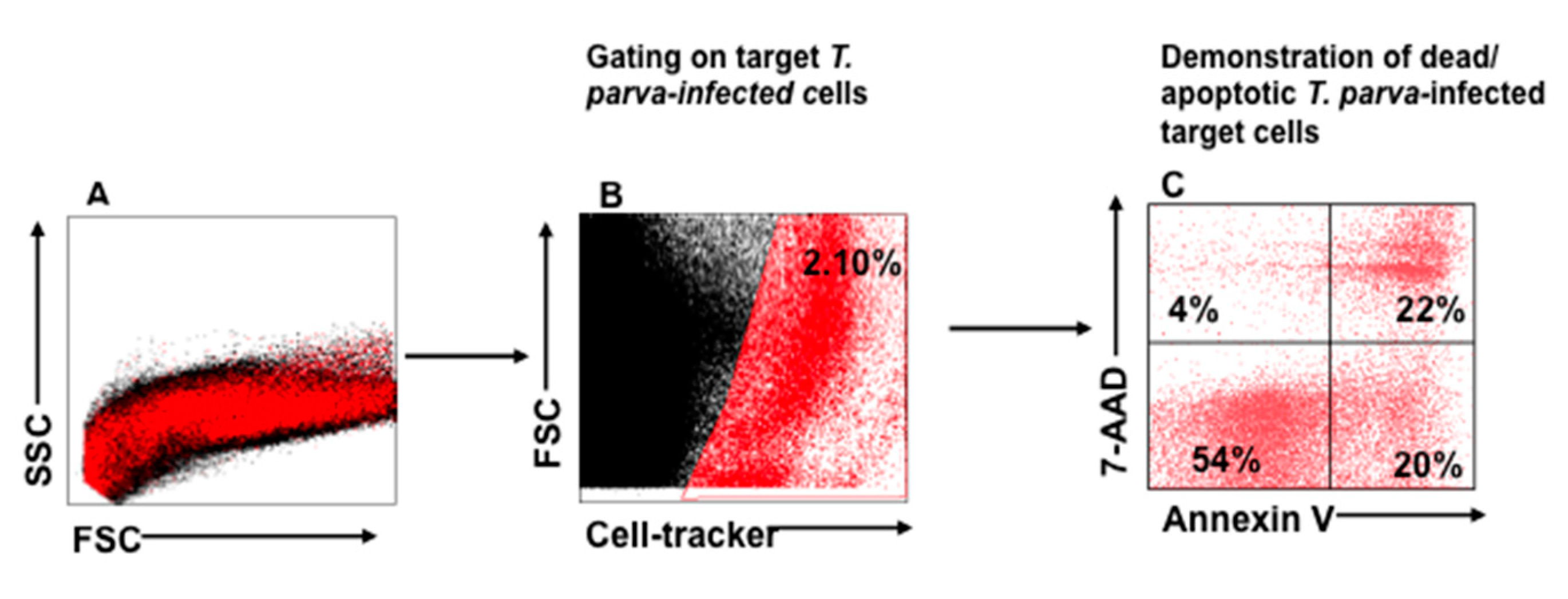
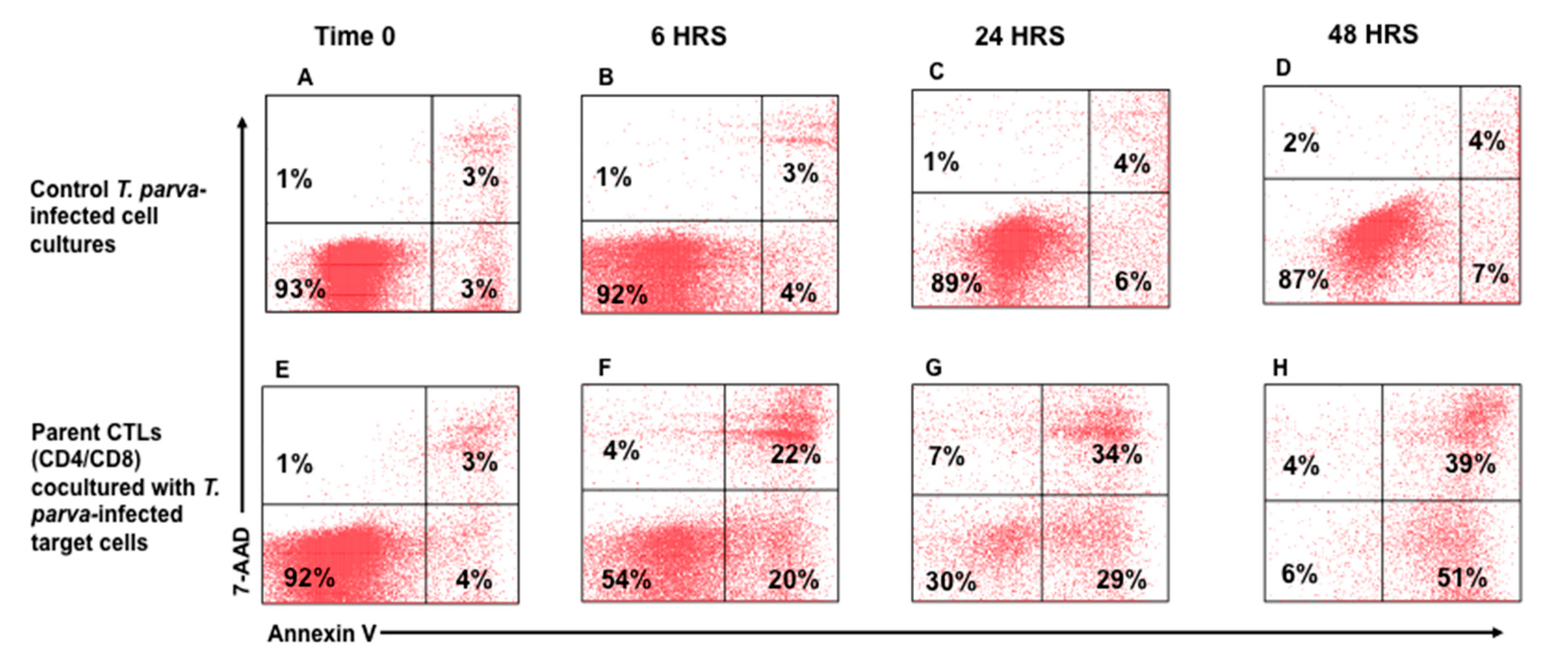
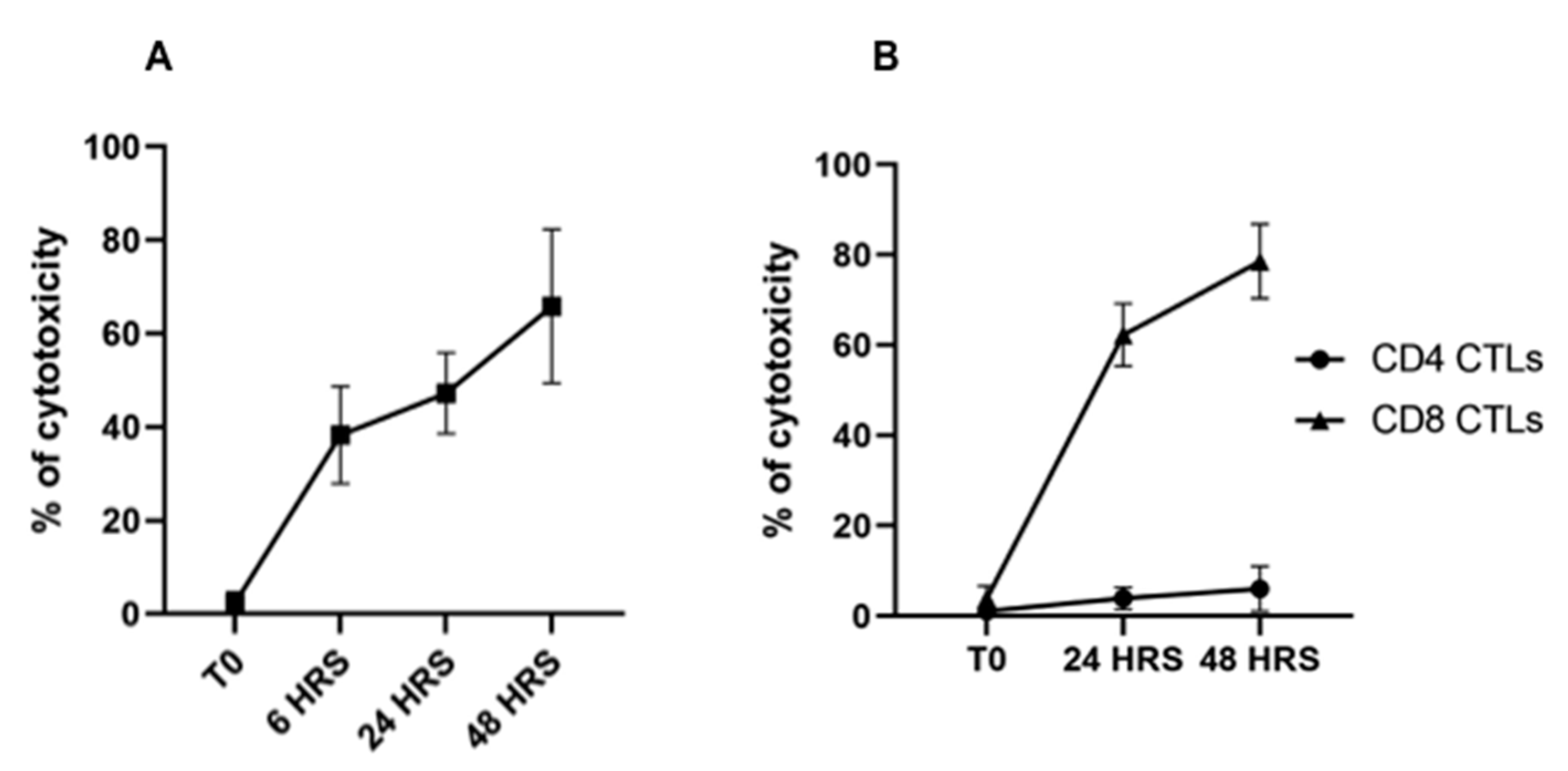
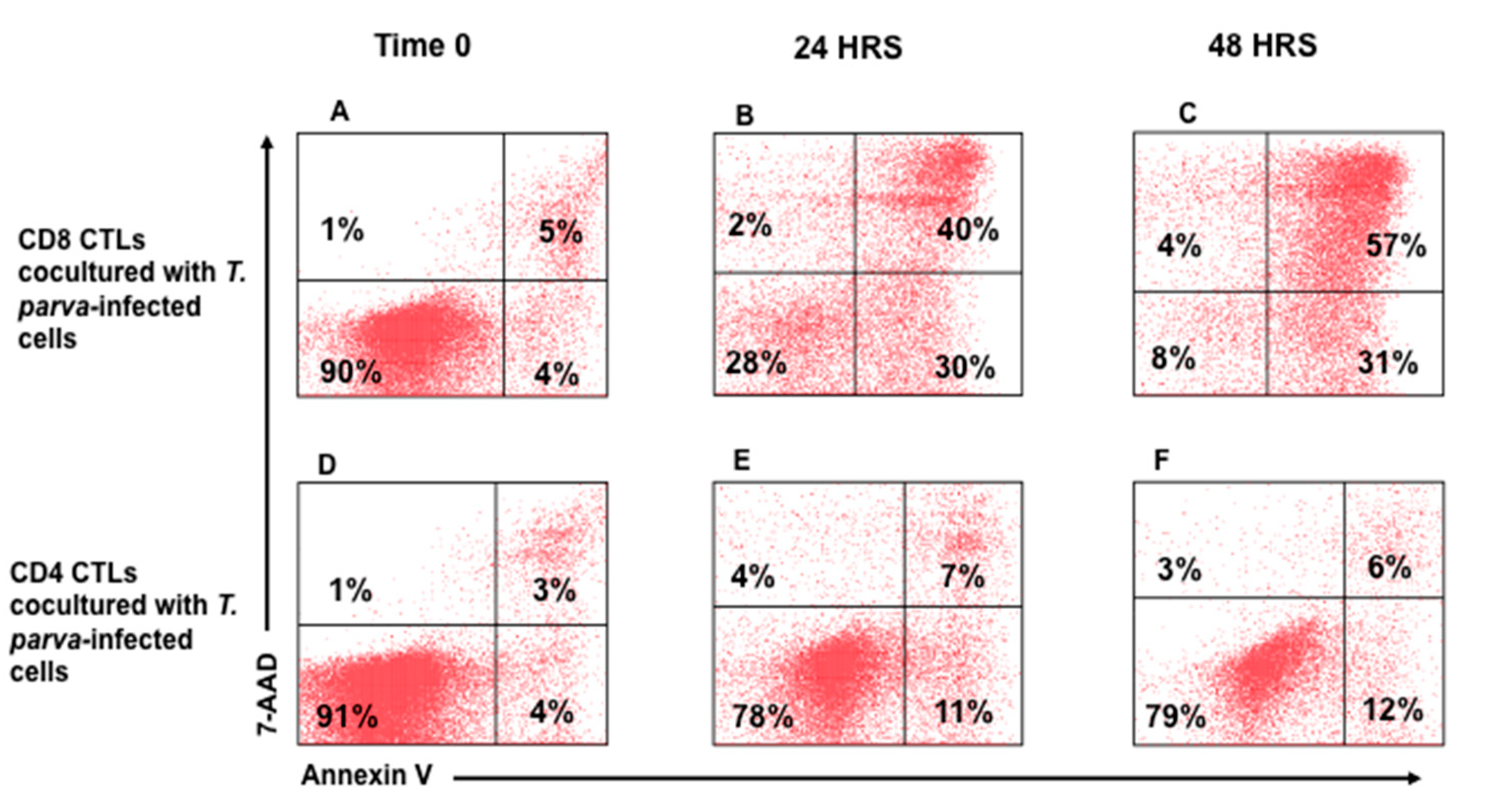
3.2. Flow Cytometric Detection of T-Cell Mediated Killing of Autologous, T. parva-Infected Cells
4. Discussion
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Nene, V.; Kiara, H.; Lacasta, A.; Pelle, R.; Svitek, N.; Steinaa, L. The biology of Theileria parva and control of East Coast fever-Current status and future trends. Ticks Tick Borne Dis. 2016, 7, 549–564. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shaw, M.K.; Tilney, L.G. How individual cells develop from a syncytium: Merogony in Theileria parva (Apicomplexa). J. Cell Sci. 1992, 101 Pt 1, 109–123. [Google Scholar] [CrossRef]
- Tretina, K.; Gotia, H.T.; Mann, D.J.; Silva, J.C. Theileria-transformed bovine leukocytes have cancer hallmarks. Trends Parasitol. 2015, 31, 306–314. [Google Scholar] [CrossRef]
- Houston, E.F.; Taracha, E.L.; Brackenbury, L.; MacHugh, N.D.; McKeever, D.J.; Charleston, B.; Morrison, W.I. Infection of cattle with Theileria parva induces an early CD8 T cell response lacking appropriate effector function. Int. J. Parasitol. 2008, 38, 1693–1704. [Google Scholar] [CrossRef] [PubMed]
- Bastos, R.G.; Sears, K.; Dinkel, K.D.; Knowles, D.P.; Fry, L.M. Changes in the Molecular and Functional Phenotype of Bovine Monocytes during Theileria parva Infection. Infect. Immun. 2019, 87. [Google Scholar] [CrossRef] [Green Version]
- Fry, L.M.; Schneider, D.A.; Frevert, C.W.; Nelson, D.D.; Morrison, W.I.; Knowles, D.P. East Coast Fever Caused by Theileria parva Is Characterized by Macrophage Activation Associated with Vasculitis and Respiratory Failure. PLoS ONE 2016, 11, e0156004. [Google Scholar] [CrossRef] [PubMed]
- Bastos, R.G.; Franceschi, V.; Tebaldi, G.; Connelley, T.; Morrison, W.I.; Knowles, D.P.; Donofrio, G.; Fry, L.M. Molecular and Antigenic Properties of Mammalian Cell-Expressed Theileria parva Antigen Tp9. Front. Immunol. 2019, 10, 897. [Google Scholar] [CrossRef] [Green Version]
- Bishop, R.P.; Odongo, D.; Ahmed, J.; Mwamuye, M.; Fry, L.M.; Knowles, D.P.; Nanteza, A.; Lubega, G.; Gwakisa, P.; Clausen, P.; et al. A review of recent research on Theileria parva: Implications for the infection and treatment vaccination method for control of East Coast fever. Transbound. Emerg. Dis. 2020, 67 (Suppl. 1), 56–67. [Google Scholar] [CrossRef]
- Patel, E.; Mwaura, S.; Kiara, H.; Morzaria, S.; Peters, A.; Toye, P. Production and dose determination of the Infection and Treatment Method (ITM) Muguga cocktail vaccine used to control East Coast fever in cattle. Ticks Tick Borne Dis. 2016, 7, 306–314. [Google Scholar] [CrossRef]
- Di Giulio, G.; Lynen, G.; Morzaria, S.; Oura, C.; Bishop, R. Live immunization against East Coast fever—Current status. Trends Parasitol. 2009, 25, 85–92. [Google Scholar] [CrossRef]
- McKeever, D.J.; Taracha, E.L.; Innes, E.L.; MacHugh, N.D.; Awino, E.; Goddeeris, B.M.; Morrison, W.I. Adoptive transfer of immunity to Theileria parva in the CD8+ fraction of responding efferent lymph. Proc. Natl. Acad. Sci. USA 1994, 91, 1959–1963. [Google Scholar] [CrossRef] [Green Version]
- Mwangi, D.M.; Honda, Y.; Graham, S.P.; Pelle, R.; Taracha, E.L.; Gachanja, J.; Nyanjui, J.K.; Bray, J.; Palmer, G.H.; Brown, W.C.; et al. Treatment of cattle with DNA-encoded Flt3L and GM-CSF prior to immunization with Theileria parva candidate vaccine antigens induces CD4 and CD8 T cell IFN-gamma responses but not CTL responses. Vet. Immunol. Immunopathol. 2011, 140, 244–251. [Google Scholar] [CrossRef]
- Graham, S.P.; Pelle, R.; Honda, Y.; Mwangi, D.M.; Tonukari, N.J.; Yamage, M.; Glew, E.J.; de Villiers, E.P.; Shah, T.; Bishop, R.; et al. Theileria parva candidate vaccine antigens recognized by immune bovine cytotoxic T lymphocytes. Proc. Natl. Acad. Sci. USA 2006, 103, 3286–3291. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Svitek, N.; Saya, R.; Awino, E.; Munyao, S.; Muriuki, R.; Njoroge, T.; Pelle, R.; Ndiwa, N.; Poole, J.; Gilbert, S.; et al. An Ad/MVA vectored Theileria parva antigen induces schizont-specific CD8(+) central memory T cells and confers partial protection against a lethal challenge. NPJ Vaccines 2018, 3, 35. [Google Scholar] [CrossRef] [Green Version]
- Fry, L.M.; Bastos, R.G.; Stone, B.C.; Williams, L.B.; Knowles, D.P.; Murphy, S.C. Gene gun DNA immunization of cattle induces humoral and CD4 T-cell-mediated immune responses against the Theileria parva polymorphic immunodominant molecule. Vaccine 2019, 37, 1546–1553. [Google Scholar] [CrossRef] [PubMed]
- Chung, C.J.; Cha, S.H.; Grimm, A.L.; Ajithdoss, D.; Rzepka, J.; Chung, G.; Yu, J.; Davis, W.C.; Ho, C.S. Pigs that recover from porcine reproduction and respiratory syndrome virus infection develop cytotoxic CD4+CD8+ and CD4+CD8- T-cells that kill virus infected cells. PLoS ONE 2018, 13, e0203482. [Google Scholar] [CrossRef] [Green Version]
- Abdellrazeq, G.S.; Elnaggar, M.M.; Bannantine, J.P.; Park, K.T.; Souza, C.D.; Backer, B.; Hulubei, V.; Fry, L.M.; Khaliel, S.A.; Torky, H.A.; et al. A Mycobacterium avium subsp. paratuberculosis relA deletion mutant and a 35 kDa major membrane protein elicit development of cytotoxic T lymphocytes with ability to kill intracellular bacteria. Vet. Res. 2018, 49, 53. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Radley, D.E.; Brown, C.G.; Cunnigham, M.P.; Kimber, C.D.; Musisi, F.L.; Purnell, R.E.; Stagg, S.M. East Coast fever: Challenge if immunised cattle by prolonged exposure to infected ticks. Vet. Rec. 1975, 96, 525–527. [Google Scholar] [CrossRef] [PubMed]
- Brown, C.G.; Radley, D.E.; Cunningham, M.P.; Kirimi, I.M.; Morzaria, S.P.; Musoke, A.J. Immunization against East Coast fever (Theileria parva infection of cattle) by infection and treatment: Chemoprophylaxis with N-pyrrolidinomethyl tetracycline. Tropenmed. Parasitol. 1977, 28, 342–348. [Google Scholar]
- Young, A.S.; Leitch, B.L.; Dolan, T.T.; Mbogo, S.K.; Ndungu, S.G.; Grootenhuis, J.G.; De Castro, J.J. Evaluation of infection and treatment methods in immunization of improved cattle against theileriosis in an endemic area of Kenya. Vet Parasit. 1990, 35, 239–257. [Google Scholar] [CrossRef]
- Baldwin, C.L.; Black, S.J.; Brown, W.C.; Conrad, P.A.; Goddeeris, B.M.; Kinuthia, S.W.; Lalor, P.A.; MacHugh, N.D.; Morrison, W.I.; Morzaria, S.P. Bovine T cells, B cells, and null cells are transformed by the protozoan parasite Theileria parva. Infect. Immun. 1988, 56, 462–467. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Goddeeris, B.M.; Morrison, W.I. Techniques for the generation, cloning, and characterization of bovine cytotoxic T cells specific for the protozoan Theileria parva. J. Tissue Cult. Methods 1988, 11, 101–110. [Google Scholar] [CrossRef]
- Storset, A.K.; Kulberg, S.; Berg, I.; Boysen, P.; Hope, J.C.; Dissen, E. NKp46 defines a subset of bovine leukocytes with natural killer cell characteristics. Eur. J. Immunol. 2004, 34, 669–676. [Google Scholar] [CrossRef]
- Allen, A.J.; Park, K.T.; Barrington, G.M.; Hamilton, M.J.; Davis, W.C. Development of a bovine ileal cannulation model to study the immune response and mechanisms of pathogenesis paratuberculosis. Clin. Vaccine Immunol. 2009, 16, 453–463. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Packard, B.Z.; Telford, W.G.; Komoriya, A.; Henkart, P.A. Granzyme B activity in target cells detects attack by cytotoxic lymphocytes. J. Immunol. 2007, 179, 3812–3820. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kim, G.G.; Donnenberg, V.S.; Donnenberg, A.D.; Gooding, W.; Whiteside, T.L. A novel multiparametric flow cytometry-based cytotoxicity assay simultaneously immunophenotypes effector cells: Comparisons to a 4 h 51Cr-release assay. J. Immunol. Methods 2007, 325, 51–66. [Google Scholar] [CrossRef] [Green Version]
- Morrison, W.I.; Goddeeris, B.M.; Brown, W.C.; Baldwin, C.L.; Teale, A.J. Theileria parva in cattle: Characterization of infected lymphocytes and the immune responses they provoke. Vet. Immunol. Immunopathol. 1989, 20, 213–237. [Google Scholar] [CrossRef]
- Baldwin, C.; Goddeeris, B.M.; Morrison, W.I. Bovine helper T-cell clones specific for lymphocytes infected with Theileria parva (Muguga). Parasite Immunol. 1987, 9, 499–513. [Google Scholar] [CrossRef]
- Baldwin, C.; Iams, K.P.; Brown, W.C.; Grab, D.J. Theileria parva: CD4+ helper and cytotoxic T-cell clones react with a schizont-derived antigen associated with the surface of Theileria parva-infected lymphocytes. Exp. Parasitol. 1992, 75, 19–30. [Google Scholar] [CrossRef]
- Brown, W.C.; Lonsdale-Eccles, J.D.; Demartini, J.C.; Grab, D.J. Recognition of soluble Theileria parva antigen by bovine helper T cell clones: Characterization and partial purification of the antigen. J Immunol. 1990, 144, 271–277. [Google Scholar]
- Daubenberger, C.A.; Taracha, E.L.; Gaidulis, L.; Davis, W.C.; McKeever, D.J. Bovine gammadelta T-cell responses to the intracellular protozoan parasite Theileria parva. Infect Immun. 1999, 67, 2241–2249. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Morrison, W.I.; Goddeeris, B.M.; Teale, A.J.; Groocock, C.M.; Kemp, S.J.; Stagg, D.A. Cytotoxic T cells elicited in cattle challenged with Theileria parva (Muguga): Evidence for restriction by class I MHC determinants and parasite strain specificity. Parasite Immunol. 1987, 9, 563–578. [Google Scholar] [CrossRef] [PubMed]
- Zaritskaya, L.; Shurin, M.R.; Sayers, T.J.; Malyguine, A.M. New flow cytometric assays for monitoring cell-mediated cytotoxicity. Expert Rev. Vaccines 2010, 9, 601–616. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Staska, L.M.; Davies, C.J.; Brown, W.C.; McGuire, T.C.; Suarez, C.E.; Park, J.Y.; Mathison, B.A.; Abbott, J.R.; Baszler, T.V. Identification of vaccine candidate peptides in the NcSRS2 surface protein of Neospora caninum by using CD4+ cytotoxic T lymphocytes and gamma interferon-secreting T lymphocytes of infected holstein cattle. Infect. Immun. 2005, 73, 1321–1329. [Google Scholar] [CrossRef] [Green Version]
- Staska, L.M.; McGuire, T.C.; Davies, C.J.; Lewin, H.A.; Baszler, T.V. Neospora caninum-infected cattle develop parasite-specific CD4+ cytotoxic T lymphocytes. Infect. Immun. 2003, 71, 3272–3279. [Google Scholar] [CrossRef] [Green Version]
- Curiel, T.J.; Krug, E.C.; Purner, M.B.; Poignard, P.; Berens, R.L. Cloned human CD4+ cytotoxic T lymphocytes specific for Toxoplasma gondii lyse tachyzoite-infected target cells. J. Immunol. 1993, 151, 2024–2031. [Google Scholar]
- Purner, M.B.; Berens, R.L.; Nash, P.B.; van Linden, A.; Ross, E.; Kruse, C.; Krug, E.C.; Curiel, T.J. CD4-mediated and CD8-mediated cytotoxic and proliferative immune responses to Toxoplasma gondii in seropositive humans. Infect. Immun. 1996, 64, 4330–4338. [Google Scholar] [CrossRef] [Green Version]
- Overgaard, N.H.; Jung, J.; Steptoe, R.J.; Wells, J.W. CD4+/CD8+ double-positive T cells: More than just a developmental stage? J. Leukoc. Biol. 2015, 97, 31–48. [Google Scholar] [CrossRef]
- Taracha, E.L.N.; Awino, E.; McKeever, D.J. Distinct CD4+ T cell helper requirements in Theileria parva-immune and -naive bovine CTL precursors. J. Immunol. 1997, 159, 4539–4545. [Google Scholar]
- Franceschi, V.; Mahmoud, A.H.; Abdellrazeq, G.S.; Tebaldi, G.; Macchi, F.; Russo, L.; Fry, L.M.; Elnaggar, M.M.; Bannantine, J.P.; Park, K.T.; et al. Capacity to Elicit Cytotoxic CD8 T Cell Activity Against Mycobacterium avium subsp. paratuberculosis Is Retained in a Vaccine Candidate 35 kDa Peptide Modified for Expression in Mammalian Cells. Front. Immunol. 2019, 10, 2859. [Google Scholar] [CrossRef]
- Abdellrazeq, G.S.; Fry, L.M.; Elnaggar, M.M.; Bannantine, J.P.; Schneider, D.A.; Chamberlin, W.M.; Mahmoud, A.H.A.; Park, K.T.; Hulubei, V.; Davis, W.C. Simultaneous cognate epitope recognition by bovine CD4 and CD8 T cells is essential for primary expansion of antigen-specific cytotoxic T-cells following ex vivo stimulation with a candidate Mycobacterium avium subsp. paratuberculosis peptide vaccine. Vaccine 2020. [Google Scholar] [CrossRef] [PubMed]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Monoclonal or Secondary Antibody | Antibody Isotype | Antibody Specificity | Antibody Supplier |
|---|---|---|---|
| ILA11A | IgG2a | Bovine CD4 | WSUMAC, USA |
| 7C2B | IgG2a | Bovine CD8 | WSUMAC, USA |
| CACT80C | IgG1 | Bovine CD8 | WSUMAC, USA |
| ILA116A | IgG1 | Bovine CD45R0 | WSUMAC, USA |
| CACT116A | IgG1 | Bovine CD25 | WSUMAC, USA |
| GB21A | IgG2b | Bovine γδ TCR | WSUMAC, USA |
| 3D12 | IgG2a | Human CCR7 | BD Pharmingen, USA |
| AKS8 | IgG2a | Bovine CD335 | Norwegian University of Life Sciences [23] |
| Goat anti-mouse | IgG2a PE.CY5.5 | ThermoFisher, USA | |
| Goat anti-mouse | IgG1 Alexa Fluor 647 | ThermoFisher, USA | |
| Goat anti-mouse | IgG1 FITC | ThermoFisher, USA | |
| Goat anti-mouse | IgG3 FITC | Southern Biotech, USA | |
| Goat anti-rat | IgG FITC | Southern Biotech, USA | |
| Goat anti-mouse | IgG2b PE | Southern Biotech, USA | |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Elnaggar, M.M.; Knowles, D.P.; Davis, W.C.; Fry, L.M. Flow Cytometric Analysis of the Cytotoxic T-Cell Recall Response to Theileria parva in Cattle Following Vaccination by the Infection and Treatment Method. Vet. Sci. 2021, 8, 114. https://0-doi-org.brum.beds.ac.uk/10.3390/vetsci8060114
Elnaggar MM, Knowles DP, Davis WC, Fry LM. Flow Cytometric Analysis of the Cytotoxic T-Cell Recall Response to Theileria parva in Cattle Following Vaccination by the Infection and Treatment Method. Veterinary Sciences. 2021; 8(6):114. https://0-doi-org.brum.beds.ac.uk/10.3390/vetsci8060114
Chicago/Turabian StyleElnaggar, Mahmoud M., Donald P. Knowles, William C. Davis, and Lindsay M. Fry. 2021. "Flow Cytometric Analysis of the Cytotoxic T-Cell Recall Response to Theileria parva in Cattle Following Vaccination by the Infection and Treatment Method" Veterinary Sciences 8, no. 6: 114. https://0-doi-org.brum.beds.ac.uk/10.3390/vetsci8060114