
Ivermectin (IVM) Possible Side Activities and Implications in Antimicrobial Resistance and Animal Welfare: The Authors’ Perspective

 , , , and
, , , and 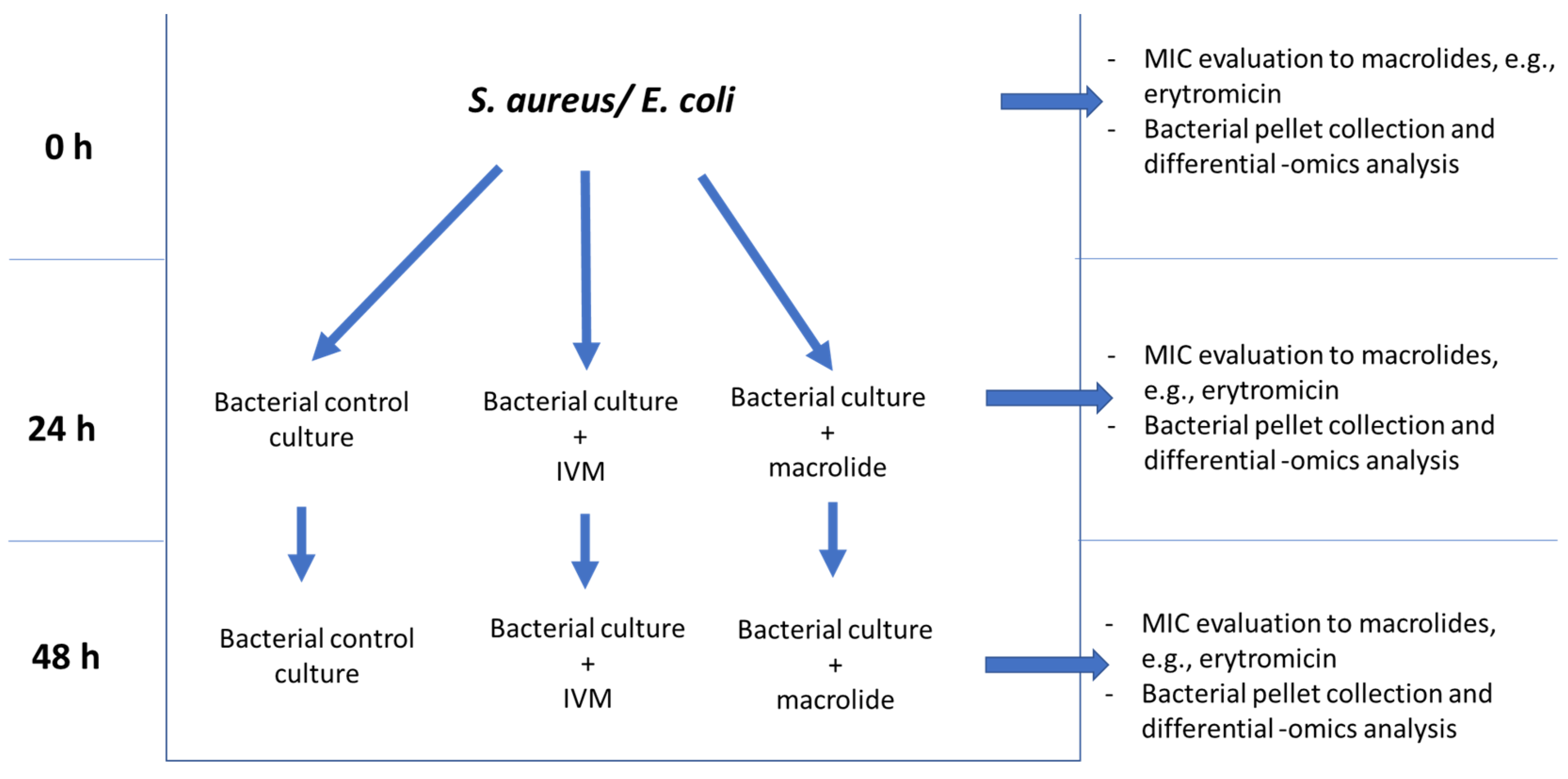{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Ivermectin Effect and Antimicrobial Resistance
3. Ivermectin’s Effect on Immune System
4. Discussion
4.1. Antimicrobial Resistance Experimental Design
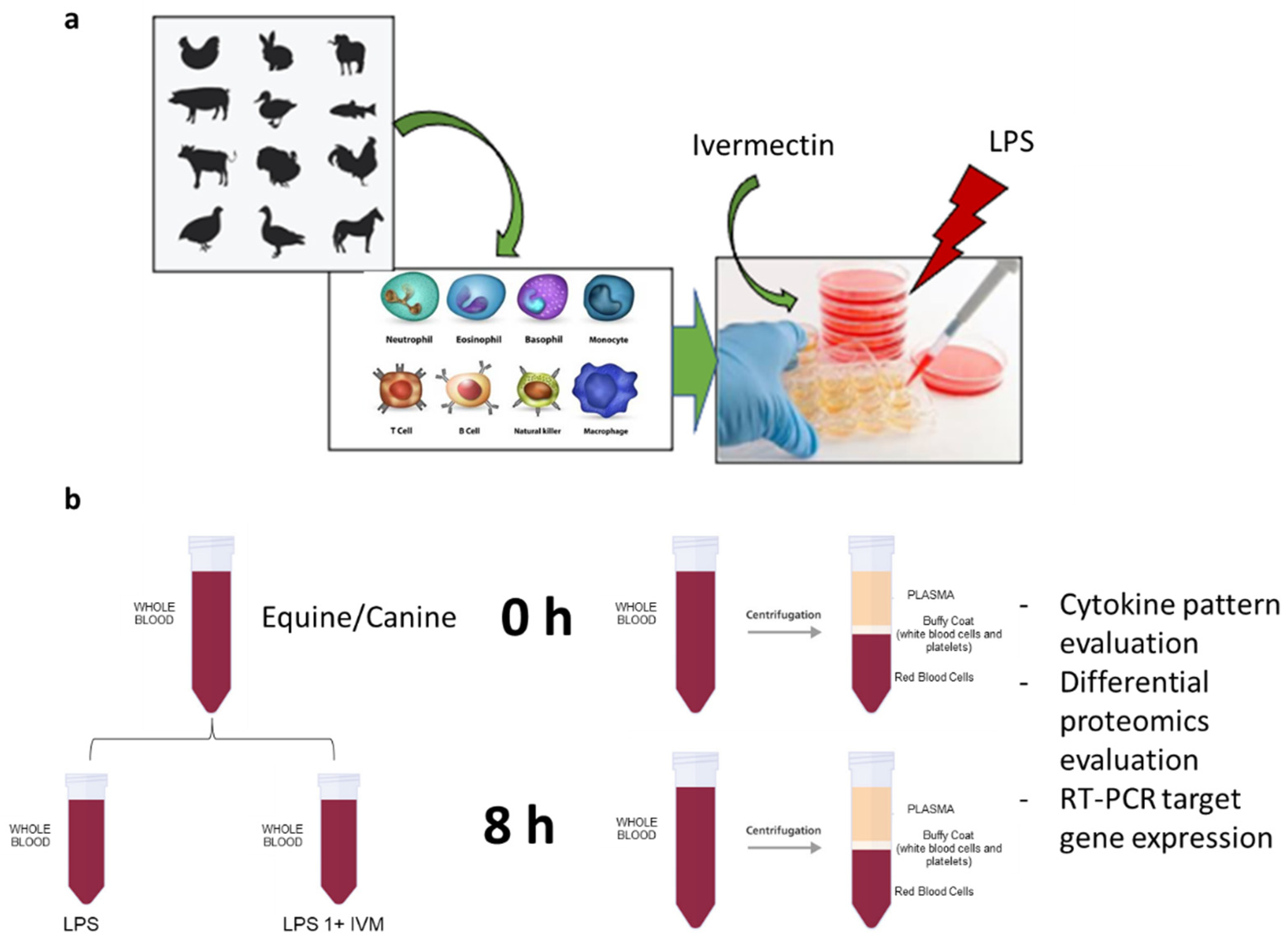
4.2. Evaluation of the Effect on Immunocompetent Cells
4.3. Green Veterinary Pharmacology
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Crump, A.; Omura, S. Ivermectin,‘wonder drug’from Japan: The human use perspective. Proc. Japan Acad. Ser. B 2011, 87, 13–28. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Crump, A. Ivermectin: Enigmatic multifaceted ‘wonder’drug continues to surprise and exceed expectations. J. Antibiot. 2017, 70, 495–505. [Google Scholar] [CrossRef] [Green Version]
- Guo, J.; Ma, R.; Su, B.; Li, Y.; Zhang, J.; Fang, J. Raising the avermectins production in Streptomyces avermitilis by utilizing nanosecond pulsed electric fields (nsPEFs). Sci. Rep. 2016, 6, 25949. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sutherland, I.H. Veterinary use of ivermectin. Acta Leiden. 1990, 59, 211–216. [Google Scholar] [PubMed]
- Lim, L.E.; Vilchèze, C.; Ng, C.; Jacobs, W.R.; Ramón-García, S.; Thompson, C.J. Anthelmintic avermectins kill Mycobacterium tuberculosis, including multidrug-resistant clinical strains. Antimicrob. Agents Chemother. 2013, 57, 1040–1046. [Google Scholar] [CrossRef] [Green Version]
- Pettengill, M.A.; Lam, V.W.; Ollawa, I.; Marques-da-Silva, C.; Ojcius, D.M. Ivermectin inhibits growth of Chlamydia trachomatis in epithelial cells. PLoS ONE 2012, 7, e48456. [Google Scholar] [CrossRef]
- Blakley, B.R.; Rousseaux, C.G. Effect of ivermectin on the immune response in mice. Am. J. Vet. Res. 1991, 52, 593–595. [Google Scholar] [PubMed]
- Omer, M.O.; Ashraf, M.; Javeed, A.; Maqbool, A. Immunostimulatory effect of ivermectin on macrophage engulfment and delayed type hypersensitivity in broilers. J. Anim. Plant Sci. 2012, 22, 250–255. [Google Scholar]
- Heidary, F.; Gharebaghi, R. Ivermectin: A systematic review from antiviral effects to COVID-19 complementary regimen. J. Antibiot. 2020, 73, 593–602. [Google Scholar] [CrossRef]
- Ianaro, A.; Ialenti, A.; Maffia, P.; Sautebin, L.; Rombolà, L.; Carnuccio, R.; Iuvone, T.; D’Acquisto, F.; Di Rosa, M. Anti-inflammatory activity of macrolide antibiotics. J. Pharmacol. Exp. Ther. 2000, 292, 156–163. [Google Scholar]
- Terao, H.; Asano, K.; Kanai, K.; Kyo, Y.; Watanabe, S.; Hisamitsu, T.; Suzaki, H. Suppressive activity of macrolide antibiotics on nitric oxide production by lipopolysaccharide stimulation in mice. Mediat. Inflamm. 2003, 12, 195–202. [Google Scholar] [CrossRef] [PubMed]
- Tkalčević, V.I.; Bošnjak, B.; Hrvačić, B.; Bosnar, M.; Marjanović, N.; Ferenčić, Ž.; Šitum, K.; Čulić, O.; Parnham, M.J.; Eraković, V. Anti-inflammatory activity of azithromycin attenuates the effects of lipopolysaccharide administration in mice. Eur. J. Pharmacol. 2006, 539, 131–138. [Google Scholar] [CrossRef]
- Viktorov, A.V.; Yurkiv, V.A. Effect of ivermectin on function of liver macrophages. Bull. Exp. Biol. Med. 2003, 136, 569–571. [Google Scholar] [CrossRef] [PubMed]
- Viktorov, A.V. Ivermectin inhibits activation of Kupffer cells induced by lipopolysaccharide toxin. Antibiot. I khimioterapiia Antibiot. Chemoterapy [sic] 2003, 48, 3–6. [Google Scholar]
- Zhang, X.; Song, Y.; Ci, X.; An, N.; Ju, Y.; Li, H.; Wang, X.; Han, C.; Cui, J.; Deng, X. Ivermectin inhibits LPS-induced production of inflammatory cytokines and improves LPS-induced survival in mice. Inflamm. Res. 2008, 57, 524–529. [Google Scholar] [CrossRef] [PubMed]
- Campbell, W.C. Ivermectin: An update. Parasitol. Today 1985, 1, 10–16. [Google Scholar] [CrossRef]
- Davies, H.G.; Green, R.H. Avermectins and milbemycins. Nat. Prod. Rep. 1986, 3, 87–121. [Google Scholar] [CrossRef]
- Blair, L.S.; Campbell, W.C. Efficacy of avermectin B1a against microfilariae of Dirofilaria immitis. Am. J. Vet. Res. 1979, 40, 1031–1032. [Google Scholar]
- Campbell, W.C.; Fisher, M.H.; Stapley, E.O.; Albers-Schonberg, G.; Jacob, T.A. Ivermectin: A potent new antiparasitic agent. Science 1983, 221, 823–828. [Google Scholar] [CrossRef]
- Piras, C.; Soggiu, A.; Bonizzi, L.; Nally, J.; Greco, V.; Urbani, A.; Martino, P.A.; Roncada, P. 2D DIGE comparative analysis of Escherichia coli strains with induced resistance to enrofloxacin. In Farm Animal Proteomics 2013; Wageningen Academic Publishers: Wageningen, The Netherlands, 2013; pp. 147–150. [Google Scholar] [CrossRef]
- Piras, C.; Soggiu, A.; Greco, V.; Martino, P.A.; Del Chierico, F.; Putignani, L.; Urbani, A.; Nally, J.E.; Bonizzi, L.; Roncada, P. Mechanisms of antibiotic resistance to enrofloxacin in uropathogenic Escherichia coli in dog. J. Proteom. 2015, 127, 365–376. [Google Scholar] [CrossRef]
- Eldholm, V.; Monteserin, J.; Rieux, A.; Lopez, B.; Sobkowiak, B.; Ritacco, V.; Balloux, F. Four decades of transmission of a multidrug-resistant Mycobacterium tuberculosis outbreak strain. Nat. Commun. 2015, 6, 7119. [Google Scholar] [CrossRef] [Green Version]
- Cohen, K.A.; Abeel, T.; McGuire, A.M.; Desjardins, C.A.; Munsamy, V.; Shea, T.P.; Walker, B.J.; Bantubani, N.; Almeida, D.V.; Alvarado, L. Evolution of extensively drug-resistant tuberculosis over four decades: Whole genome sequencing and dating analysis of Mycobacterium tuberculosis isolates from KwaZulu-Natal. PLoS Med. 2015, 12, e1001880. [Google Scholar] [CrossRef] [Green Version]
- Piras, C.; Greco, V.; Gugliandolo, E.; Soggiu, A.; Tilocca, B.; Bonizzi, L.; Zecconi, A.; Cramer, R.; Britti, D.; Urbani, A.; et al. Raw cow milk bacterial consortium as bioindicator of circulating anti-microbial resistance (Amr). Animals 2020, 10, 2378. [Google Scholar] [CrossRef]
- Rajter, J.C.; Sherman, M.S.; Fatteh, N.; Vogel, F.; Sacks, J.; Rajter, J.-J. Use of ivermectin is associated with lower mortality in hospitalized patients with COVID-19. Chest 2021, 159, 85–92. [Google Scholar] [CrossRef]
- Stankiewicz, M.; Cabaj, W.; Jonas, W.E.; Moore, L.G.; Millar, K.; Chie, W.N. Influence of ivermectin on cellular and humoral immune responses of lambs. Vet. Immunol. Immunopathol. 1995, 44, 347–358. [Google Scholar] [CrossRef]
- Sajid, M.S.; Iqbal, Z.; Muhammad, G.; Iqbal, M.U. Immunomodulatory effect of various anti-parasitics: A review. Parasitology 2006, 132, 301. [Google Scholar] [CrossRef] [PubMed]
- Gupta, D.; Sahoo, A.K.; Singh, A. Ivermectin: Potential candidate for the treatment of Covid 19. Brazilian J. Infect. Dis. 2020, 24, 369–371. [Google Scholar] [CrossRef] [PubMed]
- Berge, T.; Eriksson, A.; Brorson, I.S.; Høgestøl, E.A.; Berg-Hansen, P.; Døskeland, A.; Mjaavatten, O.; Bos, S.D.; Harbo, H.F.; Berven, F. Quantitative proteomic analyses of CD4+ and CD8+ T cells reveal differentially expressed proteins in multiple sclerosis patients and healthy controls. Clin. Proteom. 2019, 16, 19. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wiśniewski, J.R.; Zougman, A.; Nagaraj, N.; Mann, M. Universal sample preparation method for proteome analysis. Nat. Methods 2009, 6, 359–362. [Google Scholar] [CrossRef]
- Couch, C.E.; Arnold, H.K.; Crowhurst, R.S.; Jolles, A.E.; Sharpton, T.J.; Witczak, M.F.; Epps, C.W.; Beechler, B.R. Bighorn sheep gut microbiomes associate with genetic and spatial structure across a metapopulation. Sci. Rep. 2020, 10, 6582. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kolmogorov, M.; Rayko, M.; Yuan, J.; Polevikov, E.; Pevzner, P. metaFlye: Scalable long-read metagenome assembly using repeat graphs. Nat. Methods 2020, 17, 1103–1110. [Google Scholar] [CrossRef] [PubMed]
- Alcock, B.P.; Raphenya, A.R.; Lau, T.T.Y.; Tsang, K.K.; Bouchard, M.; Edalatmand, A.; Huynh, W.; Nguyen, A.-L.V.; Cheng, A.A.; Liu, S. CARD 2020: Antibiotic resistome surveillance with the comprehensive antibiotic resistance database. Nucleic Acids Res. 2020, 48, D517–D525. [Google Scholar] [CrossRef]
- Piras, C.; Soggiu, A.; Greco, V.; Nally, J.; Urbani, A.; Anna, P.; Drigo, M.; Roncada, P. Mechanisms of Enrofloxacin Antibiotic Resistance in Zoonotic E. coli; Elsevier: Amsterdan, The Netherlands, 2015. [Google Scholar]
- Castagna, F.; Britti, D.; Oliverio, M.; Bosco, A.; Bonacci, S.; Iriti, G.; Ragusa, M.; Musolino, V.; Rinaldi, L.; Palma, E. In Vitro Anthelminthic Efficacy of Aqueous Pomegranate (Punica granatum L.) Extracts against Gastrointestinal Nematodes of Sheep. Pathogens 2020, 9, 1063. [Google Scholar] [CrossRef] [PubMed]
- Castagna, F.; Piras, C.; Palma, E.; Musolino, V.; Lupia, C.; Bosco, A.; Rinaldi, L.; Cringoli, G.; Musella, V.; Britti, D. Green Veterinary Pharmacology Applied to Parasite Control: Evaluation of Punica granatum, Artemisia campestris, Salix caprea Aqueous Macerates against Gastrointestinal Nematodes of Sheep. Vet. Sci. 2021, 8, 237. [Google Scholar] [CrossRef] [PubMed]


Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Piras, C.; Gugliandolo, E.; Castagna, F.; Palma, E.; Britti, D. Ivermectin (IVM) Possible Side Activities and Implications in Antimicrobial Resistance and Animal Welfare: The Authors’ Perspective. Vet. Sci. 2022, 9, 24. https://0-doi-org.brum.beds.ac.uk/10.3390/vetsci9010024
Piras C, Gugliandolo E, Castagna F, Palma E, Britti D. Ivermectin (IVM) Possible Side Activities and Implications in Antimicrobial Resistance and Animal Welfare: The Authors’ Perspective. Veterinary Sciences. 2022; 9(1):24. https://0-doi-org.brum.beds.ac.uk/10.3390/vetsci9010024
Chicago/Turabian StylePiras, Cristian, Enrico Gugliandolo, Fabio Castagna, Ernesto Palma, and Domenico Britti. 2022. "Ivermectin (IVM) Possible Side Activities and Implications in Antimicrobial Resistance and Animal Welfare: The Authors’ Perspective" Veterinary Sciences 9, no. 1: 24. https://0-doi-org.brum.beds.ac.uk/10.3390/vetsci9010024