
Effect and Mechanism of siRNAs Targeting IL-1β/TNF-α Combined with BMSCs Transplantation in Ameliorating Rheumatoid Arthritis in Rats

Abstract
:Simple Summary
Abstract
1. Background
2. Materials and Methods
2.1. Animals
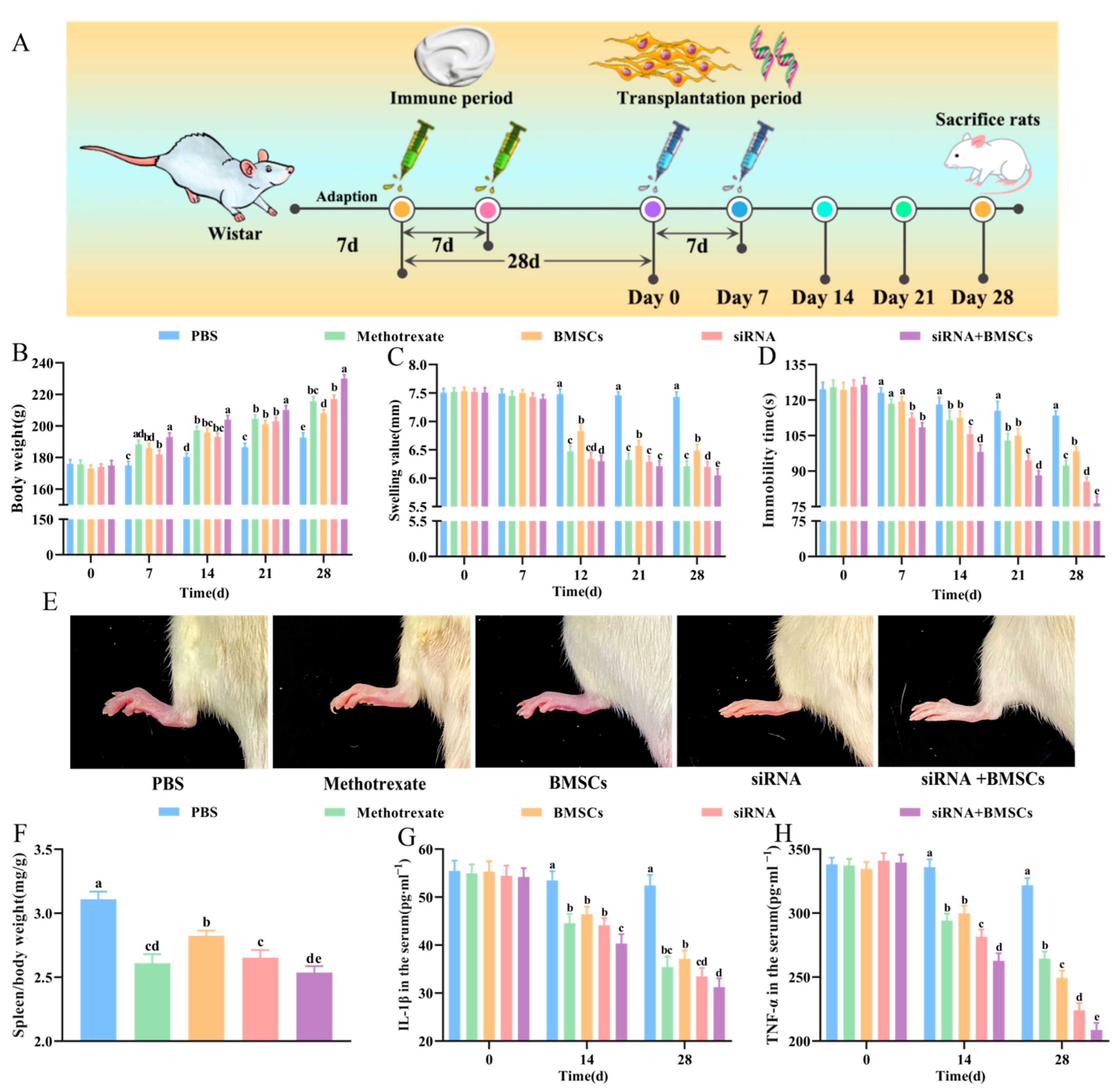
2.2. Experimental Design
2.3. Design and Synthesize IL-1β/TNF-α siRNAs
2.4. Cell Culture and IL-1β/TNF-α siRNAs Transfection
2.5. In Vivo Tracing of BMSCs
2.6. Body Weight, Toe Swelling, Forced Immobility Time and Spleen Index Measurement
2.7. Elisa
2.8. Immunohistochemistry Analysis
2.9. Haematoxylin & Eosin and Safranin O-Fast Green Staining
2.10. RNA Isolation and RT-qPCR
2.11. Statistics Analysis
3. Results
3.1. IL-1β/TNF-α siRNAs and BMSCs on LPS-Induced RAW264.7 Inflammation
3.2. Body Weight Gain, Toe Swelling, Forced Swimming Immobility Time and Spleen Index
3.3. Serum IL-1β and TNF-α Concentrations
3.4. Histopathological Analysis
3.5. Immunohistochemical Analysis
3.6. Effects of siRNA, BMSCs and siRNA + BMSCs on mRNA Expression of Inflammatory Factors in Spleen Tissue
4. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
Abbreviations
References
- Scott, D.L.; Wolfe, F.; Huizinga, T.W. Rheumatoid arthritis. Lancet 2010, 376, 1094–1108. [Google Scholar] [CrossRef]
- Napetschnig, J.; Wu, H. Molecular basis of NF-κB signaling. Annu. Rev. Biophys. 2013, 42, 443–468. [Google Scholar] [CrossRef]
- Baldwin, A. The NF-kappa B and I kappa B proteins: New discoveries and insights. Annu. Rev. Immunol. 1996, 14, 649–683. [Google Scholar] [CrossRef] [PubMed]
- Miyazawa, K.; Mori, A.; Yamamoto, K.; Okudaira, H. Constitutive transcription of the human interleukin-6 gene by rheumatoid synoviocytes: Spontaneous activation of NF-kappaB and CBF1. Am. J. Pathol. 1998, 152, 793–803. [Google Scholar] [PubMed]
- Schottelius, A.; Mayo, M.; Sartor, R.; Baldwin, A. Interleukin-10 signaling blocks inhibitor of kappaB kinase activity and nuclear factor kappaB DNA binding. J. Biol. Chem. 1999, 274, 31868–31874. [Google Scholar] [CrossRef]
- Jimi, E.; Fei, H.; Nakatomi, C. NF-κB Signaling Regulates Physiological and Pathological Chondrogenesis. Int. J. Mol. Sci. 2019, 20, 6275. [Google Scholar] [CrossRef] [PubMed]
- Quinn, M.A.; Conaghan, P.G.; O’Connor, P.J.; Karim, Z.; Greenstein, A.; Brown, A.; Brown, C.; Fraser, A.; Jarret, S.; Emery, P. Very early treatment with infliximab in addition to methotrexate in early, poor-prognosis rheumatoid arthritis reduces magnetic resonance imaging evidence of synovitis and damage, with sustained benefit after infliximab withdrawal: Results from a twelve-month randomized, double-blind, placebo-controlled trial. Arthritis Rheum. 2005, 52, 27–35. [Google Scholar] [PubMed]
- Li, J.Y.; Ren, K.K.; Zhang, W.J.; Xiao, L.; Wu, H.Y.; Liu, Q.Y.; Ding, T.; Zhang, X.C.; Nie, W.J.; Ke, Y.; et al. Human amniotic mesenchymal stem cells and their paracrine factors promote wound healing by inhibiting heat stress-induced skin cell apoptosis and enhancing their proliferation through activating PI3K/AKT signaling pathway. Stem Cell Res. Ther. 2019, 10, 247. [Google Scholar] [CrossRef] [PubMed]
- Xu, J.; Wang, B.; Sun, Y.; Wu, T.; Liu, Y.; Zhang, J.; Lee, W.Y.; Pan, X.; Chai, Y.; Li, G. Human fetal mesenchymal stem cell secretome enhances bone consolidation in distraction osteogenesis. Stem Cell Res. Ther. 2016, 7, 134. [Google Scholar] [CrossRef] [PubMed]
- Di Nicola, M.; Carlo-Stella, C.; Magni, M.; Milanesi, M.; Longoni, P.D.; Matteucci, P.; Grisanti, S.; Gianni, A.M. Human bone marrow stromal cells suppress T-lymphocyte proliferation induced by cellular or nonspecific mitogenic stimuli. Blood 2002, 99, 3838–3843. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Z.; Huang, S.; Wu, S.; Qi, J.; Li, W.; Liu, S.; Cong, Y.; Chen, H.; Lu, L.; Shi, S.; et al. Clearance of apoptotic cells by mesenchymal stem cells contributes to immunosuppression via PGE2. EBioMedicine 2019, 45, 341–350. [Google Scholar] [CrossRef]
- Pan, S.; Dong, X.; Wang, Y.; Zhou, T.; Liu, Y.; Zhou, A.; Xing, H. Transplantation of IL-1β siRNA-modified bone marrow mesenchymal stem cells ameliorates type II collagen-induced rheumatoid arthritis in rats. Exp. Ther. Med. 2022, 23, 139. [Google Scholar] [CrossRef]
- Esser, R.E.; Hildebrand, A.R.; Angelo, R.A.; Watts, L.M.; Murphey, M.D.; Baugh, L.E. Measurement of radiographic changes in adjuvant-induced arthritis in rats by quantitative image analysis. Arthritis Rheum. 1995, 38, 129–138. [Google Scholar] [CrossRef] [PubMed]
- Liu, C.; He, L.; Wang, J.; Wang, Q.; Sun, C.; Li, Y.; Jia, K.; Wang, J.; Xu, T.; Ming, R.; et al. Anti-angiogenic effect of Shikonin in rheumatoid arthritis by downregulating PI3K/AKT and MAPKs signaling pathways. J. Ethnopharmacol. 2020, 260, 113039. [Google Scholar] [CrossRef]
- Liu, C.; Kong, X.; Li, X.; Guo, W.; Zhang, C.; Sun, Y.; Su, X.; Liu, X.; Lu, A.; Lin, N. Wen Luo Yin inhibits angiogenesis in collagen-induced arthritis rat model and in vitro. J. Ethnopharmacol. 2013, 149, 478–489. [Google Scholar] [CrossRef] [PubMed]
- Atabaki, M.; Hashemi, M.; Daneshvar, H.; Alijani, E. Association between interleukin-1 receptor associated kinase 1 rs3027898 A/C gene polymorphism and rheumatoid arthritis. Biomed. Rep. 2017, 6, 335–338. [Google Scholar] [CrossRef] [PubMed]
- van der Helm-van Mil, A.H.; Huizinga, T.W. Advances in the genetics of rheumatoid arthritis point to subclassification into distinct disease subsets. Arthritis Res. Ther. 2008, 10, 205. [Google Scholar] [CrossRef] [PubMed]
- Anderson, K.L.; O’Neill, D.G.; Brodbelt, D.C.; Church, D.B.; Meeson, R.L.; Sargan, D.; Summers, J.F.; Zulch, H.; Collins, L.M. Prevalence, duration and risk factors for appendicular osteoarthritis in a UK dog population under primary veterinary care. Sci. Rep. 2018, 8, 5641. [Google Scholar] [CrossRef] [PubMed]
- McIlwraith, C.W.; Frisbie, D.D.; Kawcak, C.E. The horse as a model of naturally occurring osteoarthritis. Bone Jt. Res. 2012, 1, 297–309. [Google Scholar] [CrossRef]
- Amissah-Arthur, M.B.; Baah, W. Methotrexate-Induced Pancytopenia and Mucositis Caused by Medication Error. Ghana Med. J. 2020, 54, 68–71. [Google Scholar] [CrossRef] [PubMed]
- Fragoulis, G.E.; Conway, R.; Nikiphorou, E. Methotrexate and interstitial lung disease: Controversies and questions. A narrative review of the literature. Rheumatology 2019, 58, 1900–1906. [Google Scholar] [CrossRef] [PubMed]
- Rodenburg, R.J.; Wijnands, M.J.; Rensma, P.L. Serious complications during treatment with methotrexate: Also in chronic low-dosage use. Ned. Tijdschr. Voor Geneeskd. 2017, 161, D575. [Google Scholar]
- Juliano, R.L.; Ming, X. Recent Developments in Oligonucleotide Based Therapeutics. Preface. Adv. Drug Deliv. Rev. 2015, 87, 1–2. [Google Scholar] [CrossRef] [PubMed]
- Chi, X.; Gatti, P.; Papoian, T. Safety of antisense oligonucleotide and siRNA-based therapeutics. Drug Discov. Today 2017, 22, 823–833. [Google Scholar] [CrossRef] [PubMed]
- Chiriboga, C.A. Nusinersen for the treatment of spinal muscular atrophy. Expert Rev. Neurother. 2017, 17, 955–962. [Google Scholar] [CrossRef]
- Arend, W.P.; Dayer, J.M. Inhibition of the production and effects of interleukin-1 and tumor necrosis factor alpha in rheumatoid arthritis. Arthritis Rheum. 1995, 38, 151–160. [Google Scholar] [CrossRef]
- Arend, W.P.; Dayer, J.M. Cytokines and cytokine inhibitors or antagonists in rheumatoid arthritis. Arthritis Rheum. 1990, 33, 305–315. [Google Scholar] [CrossRef]
- McInnes, I.B.; Schett, G. Cytokines in the pathogenesis of rheumatoid arthritis. Nat. Rev. Immunol. 2007, 7, 429–442. [Google Scholar] [CrossRef]
- Brennan, F.M.; Chantry, D.; Jackson, A.; Maini, R.; Feldmann, M. Inhibitory effect of TNF alpha antibodies on synovial cell interleukin-1 production in rheumatoid arthritis. Lancet 1989, 2, 244–247. [Google Scholar] [CrossRef]
- Keffer, J.; Probert, L.; Cazlaris, H.; Georgopoulos, S.; Kaslaris, E.; Kioussis, D.; Kollias, G. Transgenic mice expressing human tumour necrosis factor: A predictive genetic model of arthritis. EMBO J. 1991, 10, 4025–4031. [Google Scholar] [CrossRef]
- Coughlan, A.R.; Robertson, D.H.; Bennett, D.; May, C.; Beynon, R.J.; Carter, S.D. Matrix metalloproteinases 2 and 9 in canine rheumatoid arthritis. Vet. Rec. 1998, 143, 219–223. [Google Scholar] [CrossRef] [PubMed]
- Reid, A.; Brady, A.; Blake, C.; Mongey, A.B.; Veale, D.J.; FitzGerald, O.; Cusack, T. Randomised controlled trial examining the effect of exercise in people with rheumatoid arthritis taking anti-TNFα therapy medication. BMC Musculoskelet. Disord. 2011, 12, 11. [Google Scholar] [CrossRef] [PubMed]
- Ross, T.N.; Kisiday, J.D.; Hess, T.; McIlwraith, C.W. Evaluation of the inflammatory response in experimentally induced synovitis in the horse: A comparison of recombinant equine interleukin 1 beta and lipopolysaccharide. Osteoarthr. Cartil. 2012, 20, 1583–1590. [Google Scholar] [CrossRef] [PubMed]
- Bazzani, C.; Filippini, M.; Caporali, R.; Bobbio-Pallavicini, F.; Favalli, E.G.; Marchesoni, A.; Atzeni, F.; Sarzi-Puttini, P.; Gorla, R. Anti-TNFalpha therapy in a cohort of rheumatoid arthritis patients: Clinical outcomes. Autoimmun. Rev. 2009, 8, 260–265. [Google Scholar] [CrossRef] [PubMed]
- Tan, W.; Gu, Z.; Leng, J.; Zou, X.; Chen, H.; Min, F.; Zhou, W.; Zhang, L.; Li, G. Let-7f-5p ameliorates inflammation by targeting NLRP3 in bone marrow-derived mesenchymal stem cells in patients with systemic lupus erythematosus. Biomed. Pharmacother. 2019, 118, 109313. [Google Scholar] [CrossRef]
- Yu, X.; Quan, J.; Long, W.; Chen, H.; Wang, R.; Guo, J.; Lin, X.; Mai, S. LL-37 inhibits LPS-induced inflammation and stimulates the osteogenic differentiation of BMSCs via P2X7 receptor and MAPK signaling pathway. Exp. Cell Res. 2018, 372, 178–187. [Google Scholar] [CrossRef]
- Watanabe, N.; Takai, S.; Morita, N.; Kawata, M.; Hirasawa, Y. A method of tracking donor cells after simulated autologous transplantation: A study using synovial cells of transgenic rats. Cell Tissue Res. 1999, 298, 519–525. [Google Scholar] [CrossRef]
- Ponte, A.L.; Marais, E.; Gallay, N.; Langonné, A.; Delorme, B.; Hérault, O.; Charbord, P.; Domenech, J. The in vitro migration capacity of human bone marrow mesenchymal stem cells: Comparison of chemokine and growth factor chemotactic activities. Stem Cells 2007, 25, 1737–1745. [Google Scholar] [CrossRef]
- Jiang, B.; Yan, L.; Wang, X.; Li, E.; Murphy, K.; Vaccaro, K.; Li, Y.; Xu, R.H. Concise Review: Mesenchymal Stem Cells Derived from Human Pluripotent Cells, an Unlimited and Quality-Controllable Source for Therapeutic Applications. Stem Cells 2019, 37, 572–581. [Google Scholar] [CrossRef]
- Ude, C.C.; Sulaiman, S.B.; Min-Hwei, N.; Hui-Cheng, C.; Ahmad, J.; Yahaya, N.M.; Saim, A.B.; Idrus, R.B. Cartilage regeneration by chondrogenic induced adult stem cells in osteoarthritic sheep model. PLoS ONE 2014, 9, e98770. [Google Scholar] [CrossRef]
- Tseng, W.J.; Huang, S.W.; Fang, C.H.; Hsu, L.T.; Chen, C.Y.; Shen, H.H.; Chang, J.Z.; Sun, J.S.; Lin, F.H. Treatment of osteoarthritis with collagen-based scaffold: A porcine animal model with xenograft mesenchymal stem cells. Histol. Histopathol. 2018, 33, 1271–1286. [Google Scholar] [PubMed]
- Lee, K.; Hui, J.; Song, I.; Ardany, L.; Lee, E. Injectable mesenchymal stem cell therapy for large cartilage defect—A porcine model. Stem Cells 2007, 25, 2964–2971. [Google Scholar] [CrossRef] [PubMed]
- Ghoryani, M.; Shariati-Sarabi, Z.; Tavakkol-Afshari, J.; Ghasemi, A.; Poursamimi, J.; Mohammadi, M. Amelioration of clinical symptoms of patients with refractory rheumatoid arthritis following treatment with autologous bone marrow-derived mesenchymal stem cells: A successful clinical trial in Iran. Biomed. Pharmacother. 2019, 109, 1834–1840. [Google Scholar] [CrossRef] [PubMed]
- Brennan, F.M.; Hayes, A.L.; Ciesielski, C.J.; Green, P.; Foxwell, B.M.; Feldmann, M. Evidence that rheumatoid arthritis synovial T cells are similar to cytokine-activated T cells: Involvement of phosphatidylinositol 3-kinase and nuclear factor kappaB pathways in tumor necrosis factor alpha production in rheumatoid arthritis. Arthritis Rheum. 2002, 46, 31–41. [Google Scholar] [CrossRef]
- Dayer, J.M.; Burger, D. Cell-cell interactions and tissue damage in rheumatoid arthritis. Autoimmun. Rev. 2004, 3 (Suppl. S1), S14–S16. [Google Scholar]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Target Genes | Sequences (5′-3′) |
|---|---|
| IL-1β siRNA | S: GGAAGGCAGUGUCACUCAUTT As: AUGAGUGACACUGCCUUCCTT |
| TNF-α siRNA | S: GGAUCUCAAAGACAACCAATT As: UUGGUUGUCUUUGAGAUCCTT |
| Name. | Gene Reference Number | Sequence (5′-3′) |
|---|---|---|
| β-actin | NM_031144.2 | F: CCTCTGAACCCTAAGGCCAA R: GTCTCCGGAGTCCATCACAA |
| IL-1β | NM_031512.2 | F: GGGATGATGACGACCTGCTA R: TGTCGTTGCTTGTCTCTCCT |
| TNF-α | X66539.1 | F: GGTCCCAACAAGGAGGAGAA R: CTCCTCTGCTTGGTGGTTTG |
| NF-κB | AF079314.2 | F: CGATCTGTTTCCCCTCATCTTTCC R: TGCGTCTTAGTGGTATCTGTGCTTCTC |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Pan, S.; Wang, L.; Wu, B.; Xing, H. Effect and Mechanism of siRNAs Targeting IL-1β/TNF-α Combined with BMSCs Transplantation in Ameliorating Rheumatoid Arthritis in Rats. Vet. Sci. 2022, 9, 531. https://0-doi-org.brum.beds.ac.uk/10.3390/vetsci9100531
Pan S, Wang L, Wu B, Xing H. Effect and Mechanism of siRNAs Targeting IL-1β/TNF-α Combined with BMSCs Transplantation in Ameliorating Rheumatoid Arthritis in Rats. Veterinary Sciences. 2022; 9(10):531. https://0-doi-org.brum.beds.ac.uk/10.3390/vetsci9100531
Chicago/Turabian StylePan, Shifeng, Lijun Wang, Bingxing Wu, and Hua Xing. 2022. "Effect and Mechanism of siRNAs Targeting IL-1β/TNF-α Combined with BMSCs Transplantation in Ameliorating Rheumatoid Arthritis in Rats" Veterinary Sciences 9, no. 10: 531. https://0-doi-org.brum.beds.ac.uk/10.3390/vetsci9100531