Morphological Structure of the Aortic Wall in Deep Diving Cetacean Species: Evidence for Diving Adaptation

 , , , , and
, , , , and
Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Animals
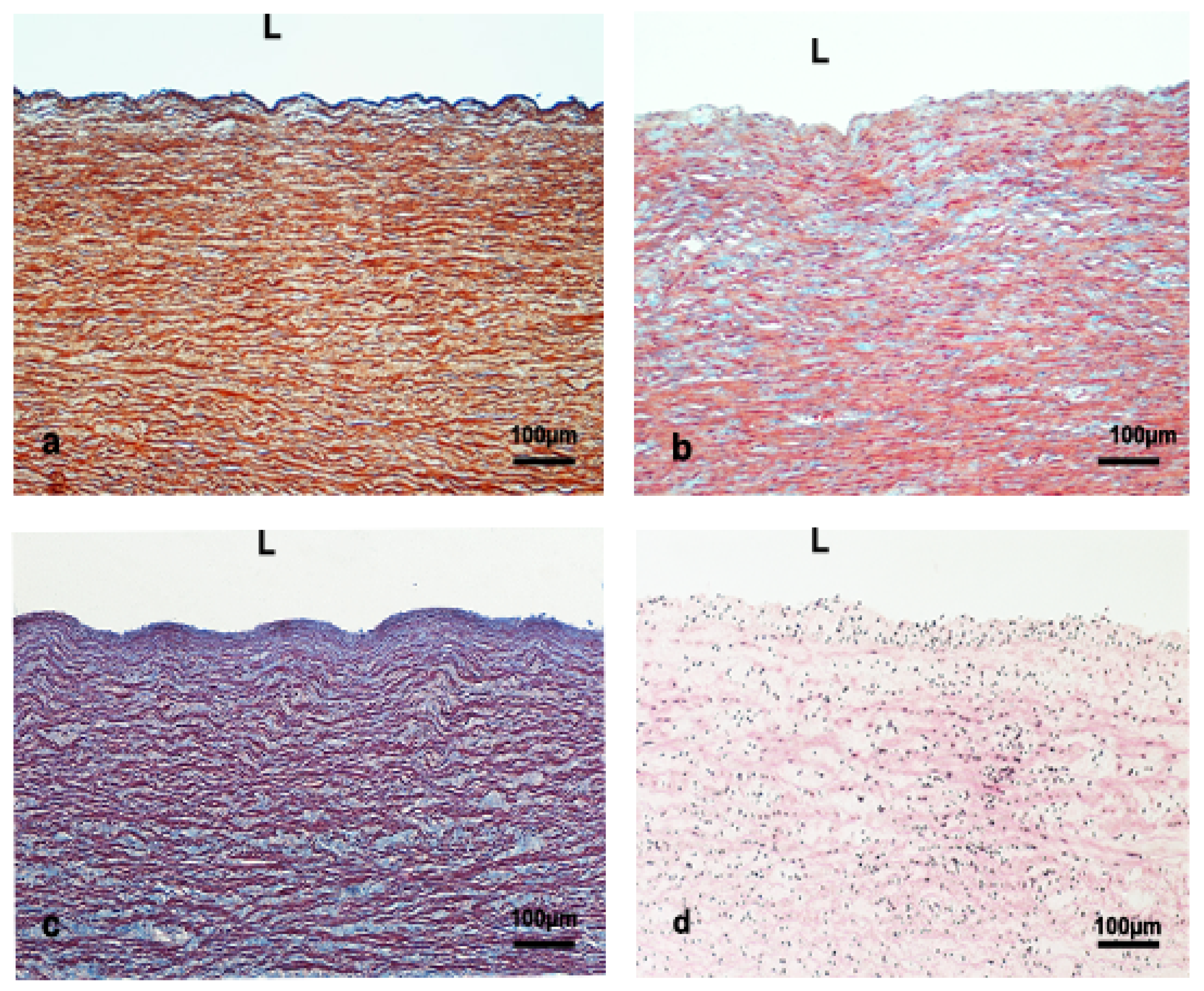
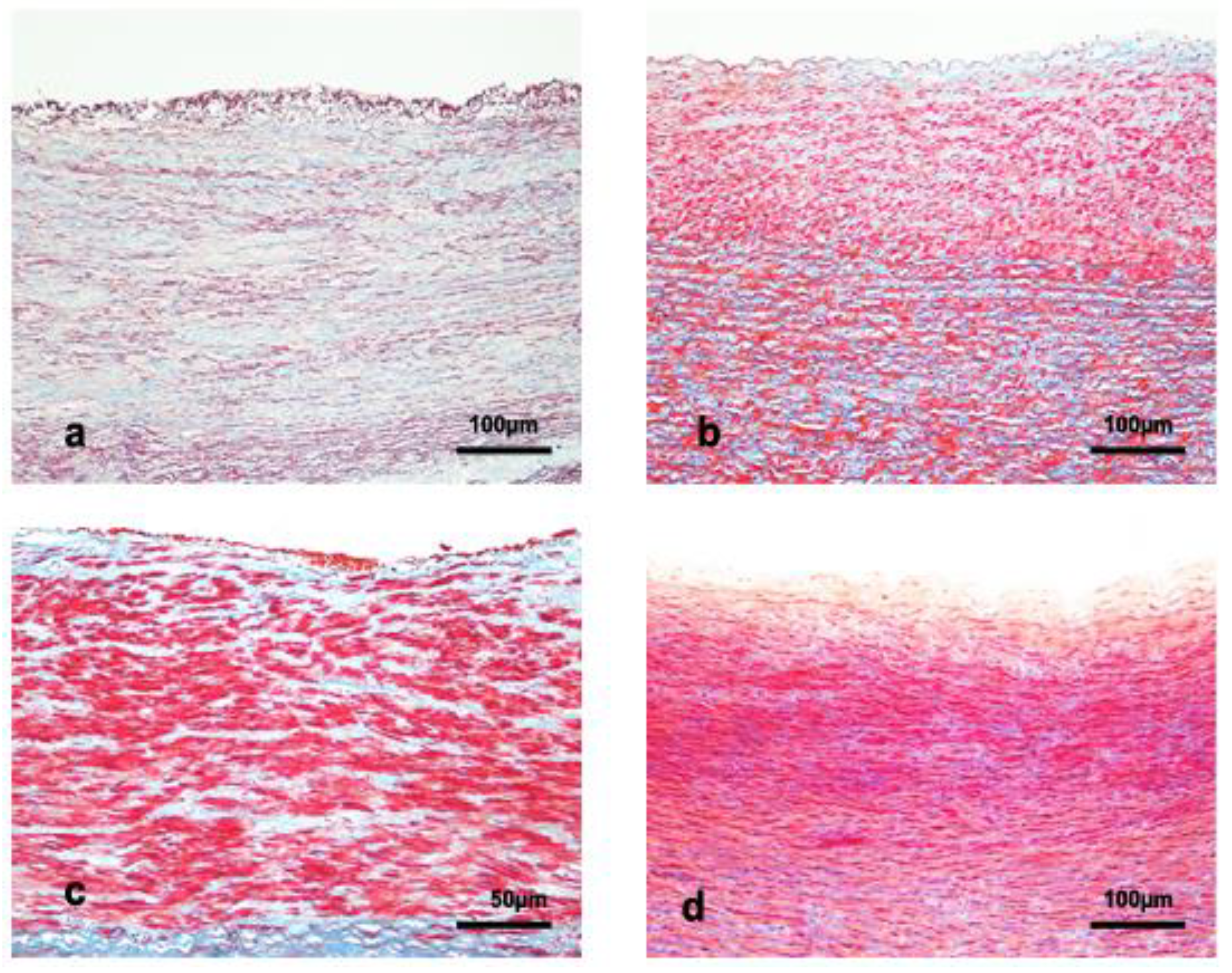
2.2. Tissue Preparation, Histology, and Histochemistry
2.3. Immunohistochemical Technique
2.4. Measurements of the Aortic Wall
2.5. Statistical Analysis
3. Results
3.1. Animals
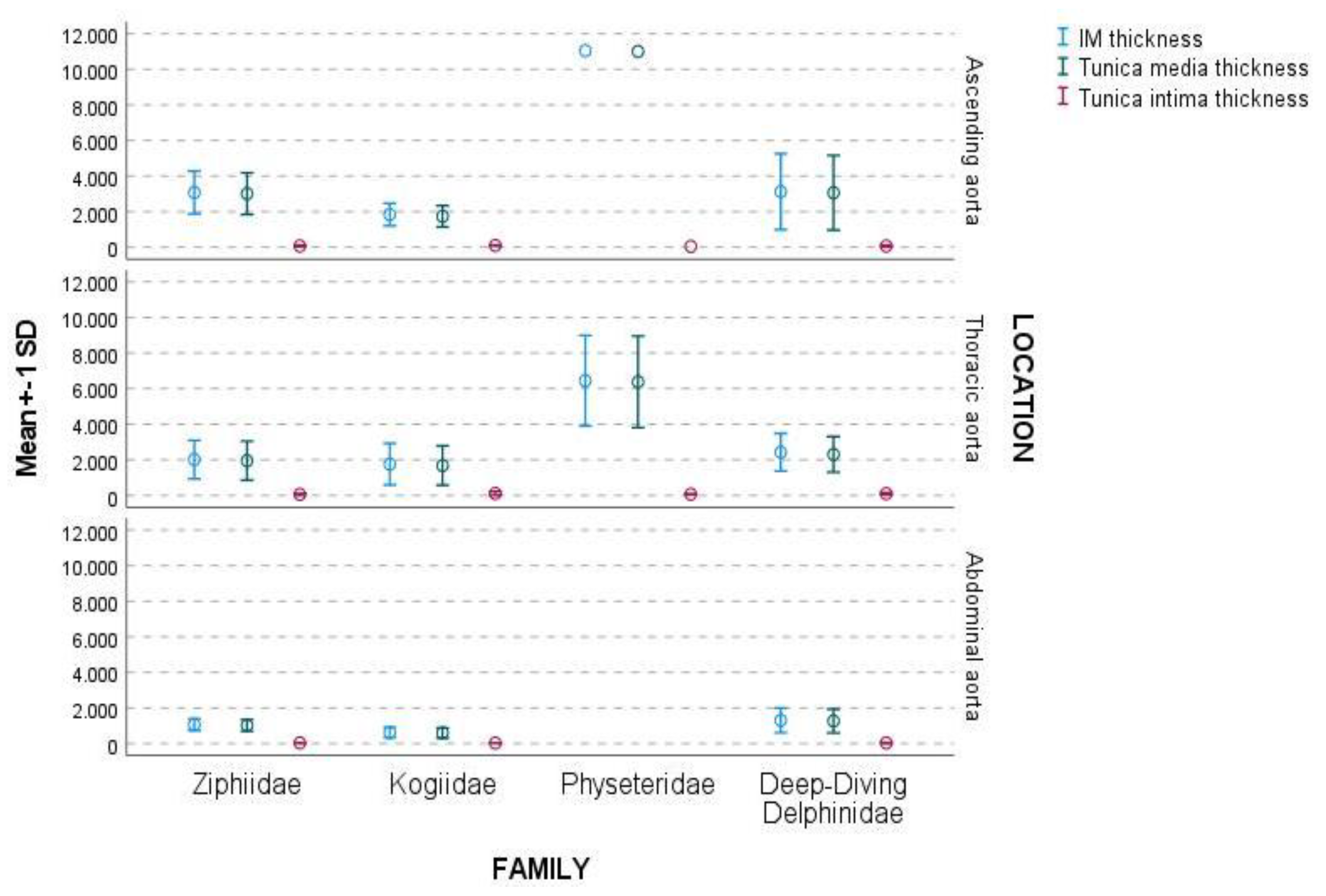
3.2. Ascending Aorta
3.3. Thoracic Aorta
3.4. Abdominal Aorta
3.5. Complete Aortic Wall
4. Discussion
4.1. Structure of the Aortic Wall
4.2. IM and Tunica Media Thickness
4.3. Lamellar Unit
4.4. Vasa Vasorum
4.5. Aortic Wall and Diving
4.6. Limitations
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Drabek, C.M.; Burns, J.M. Heart and aorta morphology of the deep-diving hooded seal (Cystophora cristata). Can. J. Zool. 2002, 80, 2030–2036. [Google Scholar] [CrossRef]
- Drabek, C.M. Some anatomical aspects of the cardiovascularv system of antarctic seals and their possible functional significance in diving. J. Morphol. 1975, 5, 85–106. [Google Scholar] [CrossRef] [PubMed]
- Shadwick, R.E.; Gosline, J.M. Arterial mechanics in the fin whale suggests unique hemodynamic design. Am. J. Physiol. 1994, 267, R805–R818. [Google Scholar] [CrossRef] [PubMed]
- Shadwick, R.E.; Gosline, J.M. Arterial windkessels in marine mammals. Symp. Soc. Exp. Biol. 1995, 49, 243–252. [Google Scholar]
- Melnikov, V.V. The arterial system of the sperm whale (Physeter macrocephalus). J. Morphol. 1997, 234, 37–50. [Google Scholar] [CrossRef]
- Lille, M.A.; Piscitelli, M.A.; Volg, A.W.; Gosline, J.M.; Shadwick, R.E. Cardiovascular design in fin whales: High-stiffness arteries protect against adverse pressure gradients at depth. J. Exp. Biol. 2013, 216, 2548–2563. [Google Scholar] [CrossRef]
- Campbell, K.B.; Rhode, E.A.; Cox, R.H.; Hunter, W.C.; Noordergraaf, A. Functional consequences of expanded aortic bulb: A model study. Am. J. Physiol. 1981, 240, R200–R210. [Google Scholar] [CrossRef] [PubMed]
- Rhode, E.A.; Elsner, R.; Peterson, T.M.; Campbell, K.B.; Spangler, W. Pressure-volume characteristics of aortas of harbor and Weddell seals. Am. J. Physiol. 1986, 251, R174–R180. [Google Scholar] [CrossRef]
- Shadwick, R.E. Mechanical design in arteries. JEB 1999, 202, 3305–3313. [Google Scholar] [CrossRef] [PubMed]
- Gosline, J.M.; Shadwick, R.E. The mechanical properties of fin whale arteries are explained by novel connective tissue designs. JEB 1996, 199, 985–997. [Google Scholar] [CrossRef]
- Smodlaka, H.; Khamas, W.; Tkalcic, S.; Golub, T.; Palmer, L. Histological assessment of selected blood vessels of the phocid seals (northern elephant and harbour seals). Anat. Histol. Embryol. 2010, 39, 178–185. [Google Scholar] [CrossRef] [PubMed]
- London, G.M.; Guerin, A.P. Influence of arterial pulse and reflected waves on blood pressure and cardiac function. Am. Heart. J. 1999, 138, 220–224. [Google Scholar] [CrossRef]
- Mompeó, B.; Pérez, L.; Fernández, A.; Saavedra, P.; Rivero, M.; Arbelo, M.; Arregui, M.; Suárez Santana, C.; Bernaldo de Quiros, Y. Morphological structure of the aortic wall in three Delphinid species with shallow or intermediate diving habits: Evidence for diving adaptation. J. Morphol. 2020, 281, 377–387. [Google Scholar] [CrossRef]
- Perrin, W.F.; Würsig, B.; Thewissen, J.G.M. Encyclopedia of Marine Mammals, 2nd ed.; Academic Press: Boston, TX, USA, 2009; pp. 255–259. [Google Scholar]
- Tejedor-Fuentes, M.L. Aportaciones a la Osteología y sus Anomalías Asociadas en los Cetáceos Menores del Archipiélago Canario. Ph.D. Thesis, Universidad de Las Palmas de Gran Canaria, Las Palmas, Spain, 2016. [Google Scholar]
- Sánchez-Sierra Campillo, C. Definición de Categorías de Edad Para las Especies Stenella frontalis y Stenella coeruloalba en Base a Parámetros Morfológicos y Madurez Gonadal. Master’s Thesis, Universidad de Las Palmas de Gran Canaria, Las Palmas, Spain, 2018. [Google Scholar]
- Geraci, J.R.; Lounsbury, V.J. Marine Mammals Ashore: A Field Guide for Strandings, 2nd ed.; National Aquarium in Baltimore: Baltimore, MD, USA, 2005. [Google Scholar]
- Bernaldo de Quiros, Y.; González-Díaz, O.; Arbelo, M.; Sierra, E.; Sacchini, S.; Fernández, A. Decompression vs decomposition: Distribution, amount, and gas composition of bubbles in stranded marine mammals. Front. Physiol. 2012, 3, 177. [Google Scholar]
- Kuiken, T.; García-Hartmann, M. Dissection techniques and tissue sampling. In: Stralsund ed. In Proceedings of the First ECS Workshop on Cetacean Pathology, Newsletter No. 17-Special Issue, Leiden, The Netherlands, 13–14 September 1991. [Google Scholar]
- Sokolis, D.P.; Boudoulas, H.; Kavantzas, N.G.; Kostomitsopoulos, N.; Agapitos, E.V.; Karayannacos, P.E. A morphometric study of the structural characteristics of the aorta in pigs using an image analysis method. Anat. Histol. Embryol. 2002, 31, 21–30. [Google Scholar] [CrossRef] [PubMed]
- Wolinsky, H.; Glagov, S.A. Lamellar unit of aortic medial structure and function in mammals. Cir. Res. 1967, 20, 99–111. [Google Scholar] [CrossRef] [PubMed]
- Wolinsky, H.; Glagov, S. Comparison of abdominal and thoracic aortic medial structure in mammals. Deviation of man from the usual pattern. Circ. Res. 1969, 6, 677–686. [Google Scholar] [CrossRef]
- Sokolis, D.P.; Boudoulas, H.; Karayannacos, P.E. Segmental differences of aortic function and composition: Clinical implications. Hellenic J. Cardiol. 2008, 49, 145–154. [Google Scholar] [PubMed]
- Tonar, Z.; Kubíková, T.; Prior, C.; Demjém, E.; Liska, V.; Králícková, M.; Witter, K. Segmental and age differences in the elastin network, collagen and smooth muscle phenotype in the tunica media of the porcine aorta. Ann. Anat. 2015, 201, 79–90. [Google Scholar] [CrossRef] [PubMed]
- Clark, J.M.; Glagov, S. The transmural organization of the arterial media; the lamellar unit revisited. Atherosclerosis 1985, 5, 19–34. [Google Scholar] [CrossRef] [PubMed]
- Dingemans, R.P.; Teeling, P.; Lagendijk, J.H.; Becker, A.E. Extracellular matrix of the human aortic media: An ultrastructural histochemical and immunohistochemical study of the adult aortic media. Anat. Rec. 2000, 258, 1–14. [Google Scholar] [CrossRef]
- Moreno, P.R.; Purushothaman, K.R.; Zias, E.; Sanz, J.; Fuster, V. Neovascularization in Human Atherosclerosis. Curr. Mol. Med. 2006, 6, 457–477. [Google Scholar] [CrossRef] [PubMed]
- Tonar, Z.; Tomášek, P.; Loskot, P.; Janáček, J.; Králíčková, M.; Witter, K. Vasa vasorum in the tunica media and tunica adventitia of the porcine aorta. Ann. Anat. 2016, 205, 22–36. [Google Scholar] [CrossRef]
- Geiringer, E. Intimal vascularization and atherosclerosis. J. Pathol. Bacteriol. 1951, 63, 201–211. [Google Scholar] [CrossRef]
- Teloni, V.; Johnson, P.M.; Miller, J.O.P.; Madsen, T.P. Shallow food for deep divers: Dynamic foraging behavior of male sperm whales in a high latitude habitat. J. Exp. Mar. Biol. Ecol. 2008, 354, 119–131. [Google Scholar] [CrossRef]
- Watkins, W.A.; Daher, M.A.; DiMarzio, N.A. Sperm whale dives tracked by radio tag telemetry. Mar. Mammal. Sci. 2002, 18, 55–68. [Google Scholar] [CrossRef]
- Culik, B.M. Review of small cetaceans: Distribution, behavior, migration and threats. In Marine Mammal Action Plan/Regional Seas Reports and Studies No. 177; UNEP/CMS Secretariat: Bonn, Germany, 2004. [Google Scholar]
- Bloodworth, B.E.; Odell, D.K. Kogia breviceps (cetacea: Kogiidae). Mamm. Species 2008, 819, 1–12. [Google Scholar] [CrossRef]
- Alves, F.; Dinis, A.; Ribeiro, C.; Nicolau, C.; Kaufmann, M.; Fortuna, C.M.; Freitas, L. Daytime dive characteristics from six short-finned pilot whales Globicephala macrorhynchus off Madeira Island Arquipélago. Life Mar. Sci. 2013, 31, 1–8. [Google Scholar]
- Shearer, J.M.; Quick, N.J.; Cioffi, W.R.; Baird, R.W.; Webster, D.L.; Foley, H.J.; Swaim, Z.T.; Waples, D.M.; Bell, J.T.; Read, A.J. Diving behavior of Cuvier’s beaked whales (Ziphius cavirostris) off Cape Hatteras, North Carolina. R. Soc. Open Sci. 2019, 6, 181728. [Google Scholar] [CrossRef] [PubMed]
- Baird, R.W.; Webster, D.L.; McSweeney, D.J.; Ligon, A.D.; Schorr, G.S.; Barlow, J. Diving behavior of Cuvier’s (Ziphius cavirostris) and Blainville’s (Mesoplodon densirostris) beaked whales in Hawai’i. Can. J. Zool. 2006, 84, 1120–1128. [Google Scholar] [CrossRef]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| FAMILY | SPECIES | M | F | U | T | LIFE STAGES | N | LENGHT | WEIGHT |
|---|---|---|---|---|---|---|---|---|---|
| ZIPHIIDAE | Mesoplodon densirostris (De Blainville, 1817) | 2 | 1 | 1 | 4 | unknown | 1 | 401.6 ± 79.3 | 645.6 ± 324.4 |
| young | 1 | ||||||||
| adult | 2 | ||||||||
| Mesoplodon Europeaus (Gervais, 1855) | 1 | 1 | calf | 1 | |||||
| Mesoplodon Bidens (Sowerby, 1804) | 1 | 1 | adult | 1 | |||||
| Ziphius cavirostris (Cuvier, 1823) | 2 | 1 | 1 | 4 | unknown | 1 | |||
| young | 2 | ||||||||
| adult | 1 | ||||||||
| PHYSETERIDAE | Physeter macrocephalus, (Linnaeus 1758) | 3 | 1 | 4 | calf | 1 | 776.2 ± 214.6 | 5375 ± 2750 | |
| young | 1 | ||||||||
| adult | 2 | ||||||||
| KOGIIDAE | Kogia breviceps (De Blainville, 1828) | 3 | 4 | 7 | calf | 4 | 177.4 ± 52.8 | 107.1 ± 112.6 | |
| young | 2 | ||||||||
| adult | 1 | ||||||||
| Kogia Sima(Owen, 1866) | 1 | 1 | calf | 1 | |||||
| DELPHINIDAE | Globicephal macrorhynchus (Gray, 1846) | 3 | 2 | 5 | young | 3 | 276.8 ± 95 | 340.1 ± 325.7 | |
| adult | 2 | ||||||||
| Grampus Griseus (Cuvier, 1812) | 1 | 3 | 4 | calf | 3 | ||||
| young | 1 | ||||||||
| SUMMARY | 15 | 15 | 3 | 31 | unknown | 2 | p = 0.000 * | p = 0.000 * | |
| calf | 10 | ||||||||
| young | 11 | ||||||||
| adult | 10 |
| Antibody | Manufacturer | RRID | Host | Type | Clone | Antigen Retrieval | Dilution |
|---|---|---|---|---|---|---|---|
| Anti- Actin | Sigma–Aldrich, St. Louis, MO, USA Cat# A5228 | AB_262054 | mouse | Monoclonal | Anti-actin α smooth-muscle antibody-1A4 | 0.1% trypsin (a) | 1:2000 (b) |
| Anti-Factor VIII | Zymed laboratories Inc, San Francisco CA94080 Cat# 18-0018 | AB_86604 | rabbit | Polyclonal | 0.1% trypsin (a) | 1:100 (b) |
| ASCENDING AORTA | THORACIC AORTA | ABDOMINAL AORTA | |||||||
|---|---|---|---|---|---|---|---|---|---|
| IM THICKNESS (μm) | |||||||||
| Family | n | Mean ± SD | Min–Max | N | Mean ± SD | Min–Max | n | Mean ± SD | Min–Max |
| Ziphiidae | 5 | 3079.8 ± 1202.1 | 1697.5–4919.8 | 9 | 2015.4 ± 1082.2 | 856.5–3837.1 | 7 | 1069.5 ±333.8 | 765.1–1775.8 |
| Kogiidae | 2 | 1834.1 ± 632.6 | 1386.8–2281.5 | 7 | 1758.5 ± 1170.7 | 488.1–3844.3 | 2 | 627.6 ± 292.4 | 420.8– 834 |
| Physeteridae | 1 | 11043.3 | - | 4 | 6445.7 ± 2547.2 | 4170.1–9782.6 | 0 | - | - |
| Deep-diving Delphinidae | 3 | 3124.1 ± 2132.5 | 1119.8–5365.1 | 7 | 2426 ± 1057.9 | 1209.9–4372.9 | 4 | 1306.5 ± 692.7 | 845.1–2330.9 |
| MEDIA THICKNESS (μm) | |||||||||
| Ziphiidae | 5 | 3008.8 ± 1173.3 | 1643.3–4790.3 | 9 | 1951.7 ± 1094.8 | 829.6–3763.5 | 7 | 1027.7 ± 322.9 | 733.3–1716.4 |
| Kogiidae | 2 | 1735.4 ± 599.1 | 1311.8–2159.1 | 7 | 1679.7 ± 1104.3 | 441.4–3571.8 | 2 | 597.6 ± 283.4 | 397.2–798 |
| Physeteridae | 1 | 10996.5 | - | 4 | 6379.8 ± 2573.1 | 4097.4–9751.2 | 0 | - | - |
| Deep-diving Delphinidae | 3 | 3059.5 ± 2099.8 | 1108.5–5281.8 | 7 | 2303.8 ± 999.6 | 1133.7–4151.1 | 4 | 1269.6 ± 668.4 | 815.7–2258,2 |
| INTIMA THICKNESS (μm) | |||||||||
| Ziphiidae | 5 | 71 ± 40.2 | 19.4–129.4 | 9 | 60.1 ± 41.8 | 8.2–147.8 | 7 | 41.8 ± 14.4 | 28.6–62.5 |
| Kogiidae | 2 | 98.7 ± 33.5 | 74.9–122.4 | 7 | 108.6 ± 107.5 | 31.8–272.4 | 2 | 29.9 ± 8.9 | 23.6–36.3 |
| Physeteridae | 1 | 46.7 | - | 4 | 66.1 ± 31.1 | 28.9–102.6 | 0 | - | - |
| Deep-diving Delphinidae | 3 | 64.5 ± 46.7 | 11.2–99 | 7 | 100.4 ± 59.8 | 46.8–221.8 | 4 | 36.9 ± 26.1 | 10.3–72.7 |
| ELASTIC THICKNESS (μm) | |||||||||
| Ziphiidae | 5 | 5.4 ± 1.7 | 3.2–7.9 | 9 | 4.5 ± 1.4 | 3.1–7.2 | 7 | 3.9 ± 1.1 | 2.26–5 |
| Kogiidae | 2 | 5.2 ± 0.3 | 4.9–5.5 | 7 | 4.1± 0.8 | 2.6–5.3 | 2 | 4.7 ± 0.4 | 4.4–5 |
| Physeteridae | 1 | 3.7 | - | 4 | 6.1 ± 1.4 | 4.5–7.9 | 0 | - | - |
| Deep-diving Delphinidae | 3 | 5.1 ± 0.9 | 4.1–6.1 | 7 | 5.4 ± 0.9 | 4.1–6.8 | 4 | 4.3 ± 0.2 | 4–4.6 |
| LAMELLAR UNIT THICKNESS (μm) | |||||||||
| Ziphiidae | 5 | 22 ± 4.7 | 16.8–26.9 | 9 | 20,05 ± 4.5 | 13.8–27.3 | 7 | 19.4 ± 6.4 | 9.7–26.2 |
| Kogiidae | 2 | 17.8 ± 3.3 | 15.4–20.1 | 7 | 21,3 ± 3.1 | 16.5–25.3 | 2 | 18.6 ± 2.2 | 17–20.2 |
| Physeteridae | 1 | 19.2 | - | 4 | 25. 5 ± 5 | 22.3–32.9 | 0 | - | - |
| Deep-diving Delphinidae | 3 | 16.7 ± 4.2 | 13.5–21.6 | 7 | 22.1 ± 5.7 | 13.1–27.5 | 4 | 19.69 ± 1.6 | 17.4–20.9 |
| NUMBER OF LAMELLAR UNITS | |||||||||
| Ziphiidae | 5 | 135 ± 38.3 | 97.5–182.4 | 9 | 102.4 ± 62.9 | 38–224.7 | 7 | 57.9 ± 24.3 | 36.8–104.2 |
| Kogiidae | 2 | 102.3 ± 52.5 | 65.1–139.5 | 7 | 72.2 ± 44,4 | 21.3–140.7 | 2 | 31.4 ± 11.3 | 23.3–39.4 |
| Physeteridae | 1 | 570.9 | - | 4 | 256.6 ± 121.6 | 169.7–436.7 | 0 | - | - |
| Deep-diving Delphinidae | 3 | 174.3 ± 89.8 | 73–244.2 | 7 | 120.6 ± 93.8 | 58.2–317.4 | 4 | 67.1 ± 42.4 | 39.1–129.9 |
| IM THICKNESS | Thoracic/Ascending Aorta | Abdominal/Ascending Aorta |
|---|---|---|
| Ziphiidae | 65% | 34.7% |
| Kogiidae | 95% | 34.2% |
| Physeteridae | 58.3% | - |
| Deep-diving Delphinidae | 77.6% | 41.8% |
| MEDIA THICKNESS | Thoracic/ascending aorta | Abdominal/Ascending aorta |
| Ziphiidae | 64% | 34,1% |
| Kogiidae | 96.7% | 34.4% |
| Physeteridae | 58% | |
| Deep-diving Delphinidae | 75.3% | 41.4% |
| LAMELLAR UNITS | Thoracic/ascending aorta | Abdominal/Ascending aorta |
| Ziphiidae | 75.8 | 42.8% |
| Kogiidae | 70.5% | 30.6% |
| Physeteridae | 44.9% | - |
| Deep-diving Delphinidae | 62.2% | 39.6% |
| N | Mean ± SD | Minimun–Maximun | |
|---|---|---|---|
| Intima thickness μm | 51 | 69.8 ± 55.9 | 8.2–272.4 |
| Elastic thickness μm | 51 | 4.7 ± 1.2 | 2.2–7.9 |
| Lamellar unit thickness μm | 51 | 20.6 ± 4.7 | 9.7–32.9 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Mompeó, B.; Sacchini, S.; Quintana, M.d.P.; Rivero, M.; Consoli, F.; Fernández, A.; Bernaldo de Quirós, Y. Morphological Structure of the Aortic Wall in Deep Diving Cetacean Species: Evidence for Diving Adaptation. Vet. Sci. 2022, 9, 424. https://0-doi-org.brum.beds.ac.uk/10.3390/vetsci9080424
Mompeó B, Sacchini S, Quintana MdP, Rivero M, Consoli F, Fernández A, Bernaldo de Quirós Y. Morphological Structure of the Aortic Wall in Deep Diving Cetacean Species: Evidence for Diving Adaptation. Veterinary Sciences. 2022; 9(8):424. https://0-doi-org.brum.beds.ac.uk/10.3390/vetsci9080424
Chicago/Turabian StyleMompeó, Blanca, Simona Sacchini, María del Pino Quintana, Miguel Rivero, Francesco Consoli, Antonio Fernández, and Yara Bernaldo de Quirós. 2022. "Morphological Structure of the Aortic Wall in Deep Diving Cetacean Species: Evidence for Diving Adaptation" Veterinary Sciences 9, no. 8: 424. https://0-doi-org.brum.beds.ac.uk/10.3390/vetsci9080424