From Stripes to a Beating Heart: Early Cardiac Development in Zebrafish


{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction: Zebrafish to Study Earliest Heart Development
2. Earliest Developmental Steps towards Heart Formation
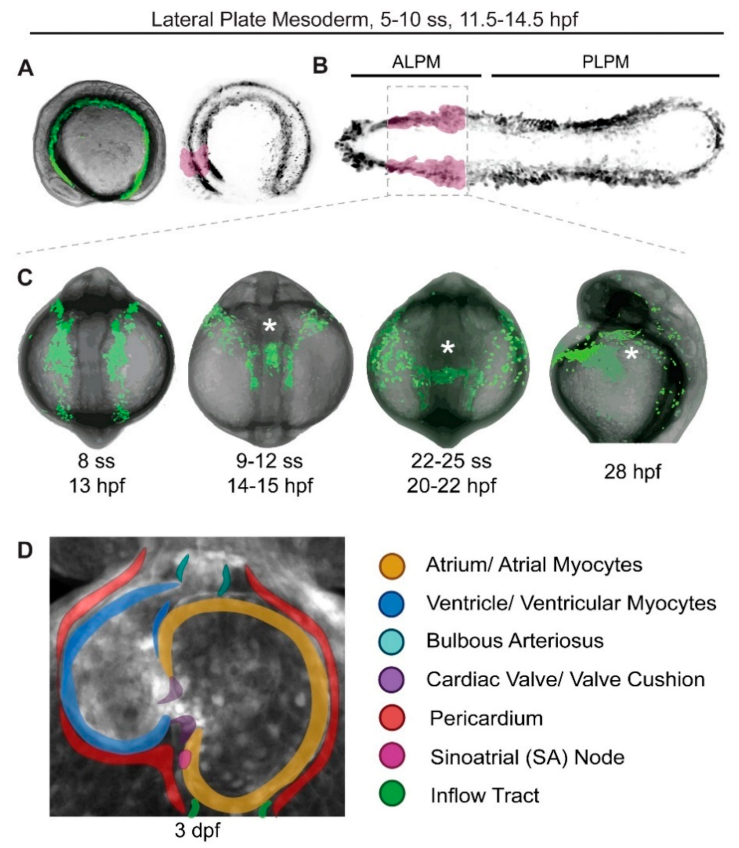
3. Labeling and Live Imaging of Heart Progenitors
4. Heart Formation from the First and Second Heart Fields
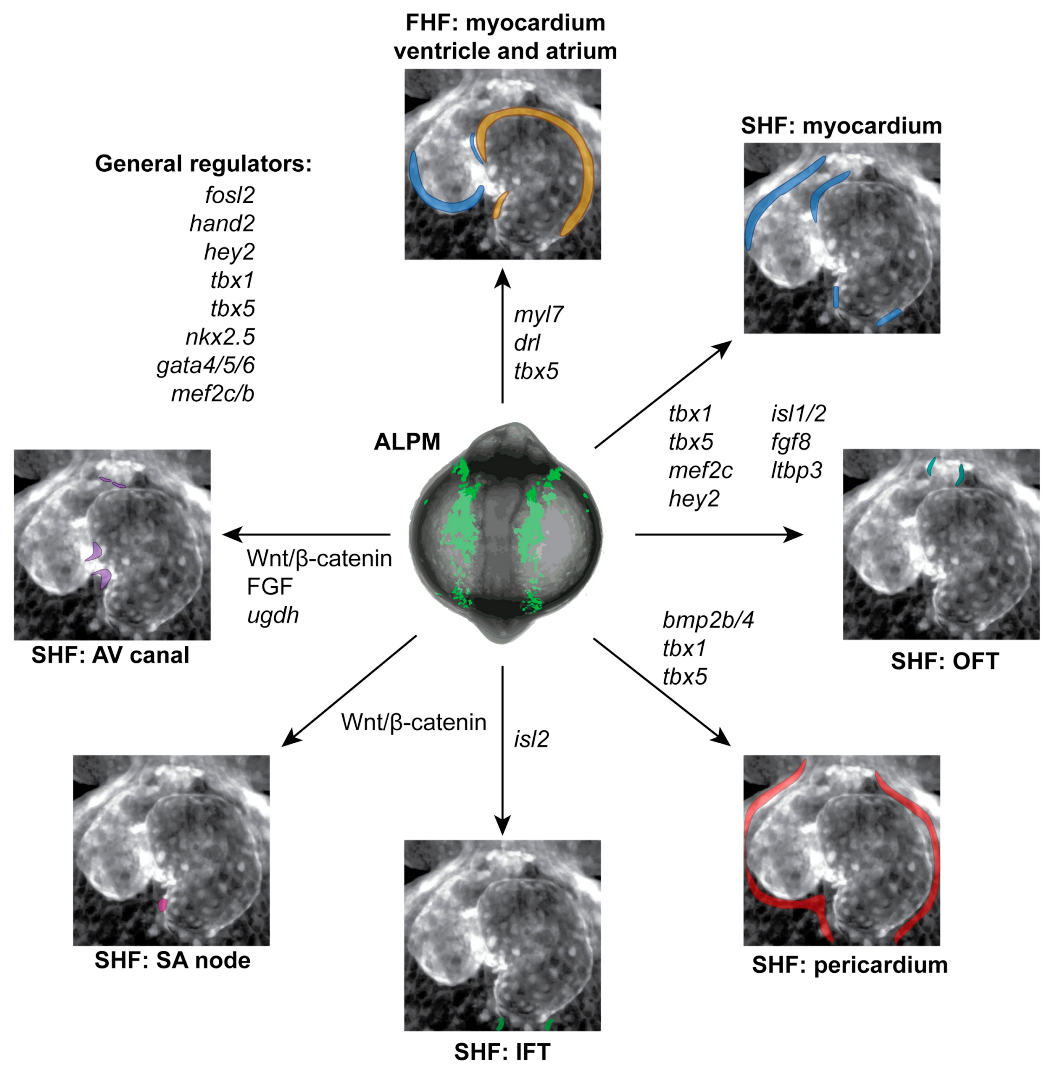
5. Developmental Signaling in Early Heart Development
6. Connecting Individual Players to Developmental Interactions
7. The Cardiopharyngeal Field as Context for Heart Formation
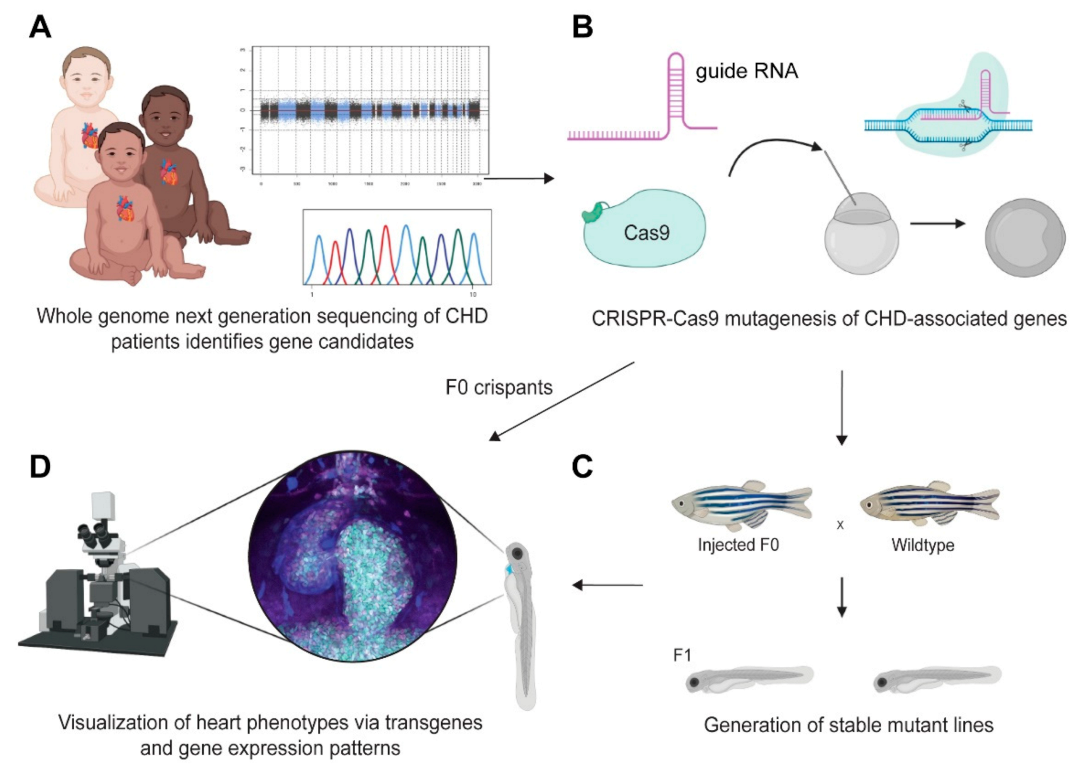
8. Modeling Early Causes of Congenital Heart Disease in Zebrafish
9. Conclusions and Future Challenges
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Lieschke, G.J.; Currie, P.D. Animal models of human disease: Zebrafish swim into view. Nat. Rev. Genet. 2007, 8, 353–367. [Google Scholar] [CrossRef]
- Meyers, J.R. Zebrafish: Development of a Vertebrate Model Organism. Curr. Protoc. Essent. Lab. Tech. 2018. [Google Scholar] [CrossRef] [Green Version]
- Kelly, R.G. The Second Heart Field. In Current Topics in Developmental Biology; Elsevier: Amsterdam, The Netherlands, 2012; Volume 100, pp. 33–65. [Google Scholar]
- Christoffels, V.; Jensen, B. Cardiac Morphogenesis: Specification of the Four-Chambered Heart. Cold Spring Harb. Perspect. Biol. 2020, a037143. [Google Scholar] [CrossRef] [PubMed]
- Stainier, D.Y.R. Zebrafish genetics and vertebrate heart formation. Nat. Rev. Genet. 2001, 2, 39–48. [Google Scholar] [CrossRef]
- Bakkers, J. Zebrafish as a model to study cardiac development and human cardiac disease. Cardiovasc. Res. 2011, 91, 279–288. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Stainier, D.Y.; Fouquet, B.; Chen, J.N.; Warren, K.S.; Weinstein, B.M.; Meiler, S.E.; Mohideen, M.A.; Neuhauss, S.C.; Solnica-Krezel, L.; Schier, A.F.; et al. Mutations affecting the formation and function of the cardiovascular system in the zebrafish embryo. Development 1996, 123, 285–292. [Google Scholar]
- Alexander, J.; Rothenberg, M.; Henry, G.L.; Stainier, D.Y. Casanova plays an early and essential role in endoderm formation in zebrafish. Dev. Biol. 1999, 215, 343–357. [Google Scholar] [CrossRef] [Green Version]
- Dickmeis, T.; Mourrain, P.; Saint-Etienne, L.; Fischer, N.; Aanstad, P.; Clark, M.; Strähle, U.; Rosa, F. A crucial component of the endoderm formation pathway, CASANOVA, is encoded by a novel sox-related gene. Genes Dev. 2001, 15. [Google Scholar] [CrossRef] [Green Version]
- Osborne, N.; Brand-Arzamendi, K.; Ober, E.A.; Jin, S.W.; Verkade, H.; Holtzman, N.G.; Yelon, D.; Stainier, D.Y.R. The Spinster Homolog, Two of Hearts, Is Required for Sphingosine 1-Phosphate Signaling in Zebrafish. Curr. Biol. 2008, 18, 1882–1888. [Google Scholar] [CrossRef] [Green Version]
- Just, S.; Raphel, L.; Berger, I.M.; Bühler, A.; Keßler, M.; Rottbauer, W. Tbx20 Is an Essential Regulator of Embryonic Heart Growth in Zebrafish. PLoS ONE 2016, 11, e0167306. [Google Scholar] [CrossRef] [PubMed]
- Garrity, D.M.; Childs, S.; Fishman, M.C. The heartstrings mutation in zebrafish causes heart/fin Tbx5 deficiency syndrome. Development 2002, 129, 4635–4645. [Google Scholar]
- Yelon, D.; Ticho, B.; Halpern, M.E.; Ruvinsky, I.; Ho, R.K.; Silver, L.M.; Stainier, D.Y.R. The bHLH transcription factor hand2 plays parallel roles in zebrafish heart and pectoral fin development. Development 2000, 127, 2573–2582. [Google Scholar] [PubMed]
- Stalsberg, H.; DeHaan, R.L. The precardiac areas and formation of the tubular heart in the chick embryo. Dev. Biol. 1969, 19, 128–159. [Google Scholar] [CrossRef]
- Stainier, D.Y.; Lee, R.K.; Fishman, M.C. Cardiovascular development in the zebrafish. I. Myocardial fate map and heart tube formation. Development 1993, 119, 31–40. [Google Scholar]
- Liu, F.; Bhang, S.H.; Arentson, E.; Sawada, A.; Kim, C.K.; Kang, I.; Yu, J.; Sakurai, N.; Kim, S.H.; Yoo, J.J.W.; et al. Enhanced hemangioblast generation and improved vascular repair and regeneration from embryonic stem cells by defined transcription factors. Stem Cell Reports 2013, 1, 166–182. [Google Scholar] [CrossRef] [Green Version]
- Rawles, M.E. The Heart-Forming Areas of the Early Chick Blastoderm. Physiol. Zool. 1943, 16, 22–43. [Google Scholar] [CrossRef]
- Kaufman, M.H.; Navaratnam, V. Early differentiation of the heart in mouse embryos. J. Anat. 1981, 133, 235–246. [Google Scholar] [PubMed]
- Prummel, K.D.; Nieuwenhuize, S.; Mosimann, C. The lateral plate mesoderm. Development 2020, 147. [Google Scholar] [CrossRef] [PubMed]
- Haack, T.; Abdelilah-Seyfried, S. The force within: Endocardial development, mechanotransduction and signalling during cardiac morphogenesis. Development 2016, 143, 373–386. [Google Scholar] [CrossRef] [Green Version]
- Khodiyar, V.K.; Howe, D.; Talmud, P.J.; Breckenridge, R.; Lovering, R.C. From zebrafish heart jogging genes to mouse and human orthologs: Using Gene Ontology to investigate mammalian heart development. F1000Research 2013. [Google Scholar] [CrossRef]
- Chen, J.N.; van Eeden, F.J.M.; Warren, K.S.; Chin, A.; Nüsslein-Volhard, C.; Haffter, P.; Fishman, M.C. Left-right pattern of cardiac BMP4 may drive asymmetry of the heart in zebrafish. Development 1997, 124, 4373–4382. [Google Scholar] [PubMed]
- Grimes, D.T.; Patterson, V.L.; Luna-Arvizu, G.; Schottenfeld-Roames, J.; Irons, Z.H.; Burdine, R.D. Left-right asymmetric heart jogging increases the robustness of dextral heart looping in zebrafish. Dev. Biol. 2020, 459, 79–86. [Google Scholar] [CrossRef] [PubMed]
- Mosimann, C.; Panáková, D.; Werdich, A.A.; Musso, G.; Burger, A.; Lawson, K.L.; Carr, L.A.; Nevis, K.R.; Sabeh, M.K.; Zhou, Y.; et al. Chamber identity programs drive early functional partitioning of the heart. Nat. Commun. 2015, 6. [Google Scholar] [CrossRef] [Green Version]
- Pan, Y.A.; Freundlich, T.; Weissman, T.A.; Schoppik, D.; Wang, X.C.; Zimmerman, S.; Ciruna, B.; Sanes, J.R.; Lichtman, J.W.; Schier, A.F. Zebrabow: Multispectral cell labeling for cell tracing and lineage analysis in zebrafish. Development 2013, 140, 2835–2846. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Beis, D.; Bartman, T.; Jin, S.-W.W.; Scott, I.C.; D’Amico, L.A.; Ober, E.A.; Verkade, H.; Frantsve, J.; Field, H.A.; Wehman, A.; et al. Genetic and cellular analyses of zebrafish atrioventricular cushion and valve development. Development 2005, 132, 4193–4204. [Google Scholar] [CrossRef] [Green Version]
- Lorenzale, M.; López-Unzu, M.A.; Rodríguez, C.; Fernández, B.; Durán, A.C.; Sans-Coma, V. The anatomical components of the cardiac outflow tract of chondrichthyans and actinopterygians. Biol. Rev. 2018, 93, 1604–1619. [Google Scholar] [CrossRef] [PubMed]
- Grimes, A.C.; Kirby, M.L. The outflow tract of the heart in fishes: Anatomy, genes and evolution. J. Fish Biol. 2009, 74, 983–1036. [Google Scholar] [CrossRef]
- Tessadori, F.; van Weerd, J.H.; Burkhard, S.B.; Verkerk, A.O.; de Pater, E.; Boukens, B.J.; Vink, A.; Christoffels, V.M.; Bakkers, J. Identification and Functional Characterization of Cardiac Pacemaker Cells in Zebrafish. PLoS ONE 2012. [Google Scholar] [CrossRef] [Green Version]
- Ferretti, E.; Hadjantonakis, A.-K. Mesoderm specification and diversification: From single cells to emergent tissues. Curr. Opin. Cell Biol. 2019, 61, 110–116. [Google Scholar] [CrossRef]
- Abu-Issa, R.; Kirby, M.L. Heart field: From mesoderm to heart tube. Annu. Rev Cell Dev. Biol. 2007, 23, 45–68. [Google Scholar] [CrossRef]
- Prummel, K.D.; Hess, C.; Nieuwenhuize, S.; Parker, H.J.; Rogers, K.W.; Kozmikova, I.; Racioppi, C.; Brombacher, E.C.; Czarkwiani, A.; Knapp, D.; et al. A conserved regulatory program initiates lateral plate mesoderm emergence across chordates. Nat. Commun. 2019, 10, 3857. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Keegan, B.R.; Meyer, D.; Yelon, D. Organization of cardiac chamber progenitors in the zebrafish blastula. Development 2004, 131, 3081–3091. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Schoenebeck, J.J.; Keegan, B.R.; Yelon, D. Vessel and blood specification override cardiac potential in anterior mesoderm. Dev. Cell 2007, 13, 254–267. [Google Scholar] [CrossRef] [Green Version]
- Waxman, J.S.; Keegan, B.R.; Roberts, R.W.; Poss, K.D.; Yelon, D. Hoxb5b acts downstream of retinoic acid signaling in the forelimb field to restrict heart field potential in zebrafish. Dev. Cell 2008, 15, 923–934. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lescroart, F.; Wang, X.; Lin, X.; Swedlund, B.; Gargouri, S.; Sànchez-Dànes, A.; Moignard, V.; Dubois, C.; Paulissen, C.; Kinston, S.; et al. Defining the earliest step of cardiovascular lineage segregation by single-cell RNA-seq. Science 2018, eaao4174. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Costello, I.; Pimeisl, I.-M.; Dräger, S.; Bikoff, E.K.; Robertson, E.J.; Arnold, S.J. The T-box transcription factor Eomesodermin acts upstream of Mesp1 to specify cardiac mesoderm during mouse gastrulation. Nat. Cell Biol. 2011, 13, 1084–1091. [Google Scholar] [CrossRef] [Green Version]
- Satou, Y.; Imai, K.S.; Satoh, N. The ascidian Mesp gene specifies heart precursor cells. Development 2004, 131, 2533–2541. [Google Scholar] [CrossRef] [Green Version]
- Saga, Y.; Miyagawa-Tomita, S.; Takagi, A.; Kitajima, S.; Miyazaki, J.I.; Inoue, T. MesP1 is expressed in the heart precursor cells and required for the formation of a single heart tube. Development 1999, 126, 3437–3447. [Google Scholar]
- Deshwar, A.R.; Onderisin, J.C.; Aleksandrova, A.; Yuan, X.; Burrows, J.T.A.; Scott, I.C. Mespaa can potently induce cardiac fates in zebrafish. Dev. Biol. 2016, 418, 17–27. [Google Scholar] [CrossRef]
- Yabe, T.; Hoshijima, K.; Yamamoto, T.; Takada, S. Quadruple zebrafish mutant reveals different roles of Mesp genes in somite segmentation between mouse and zebrafish. Development 2016, 143. [Google Scholar] [CrossRef] [Green Version]
- Harvey, R.P. Patterning the vertebrate heart. Nat. Rev. Genet. 2002, 3, 544–556. [Google Scholar] [CrossRef]
- Lyons, I.; Parsons, L.M.; Hartley, L.; Li, R.; Andrews, J.E.; Robb, L.; Harvey, R.P. Myogenic and morphogenetic defects in the heart tubes of murine embryos lacking the homeo box gene Nkx2-5. Genes Dev. 1995, 9, 1654–1666. [Google Scholar] [CrossRef] [Green Version]
- Tanaka, M.; Chen, Z.; Bartunkova, S.; Yamasaki, N.; Izumo, S. The cardiac homeobox gene Csx/Nkx2.5 lies genetically upstream of multiple genes essential for heart development. Development 1999, 126, 1269–1280. [Google Scholar]
- Lee, K.H.; Xu, Q.; Breitbart, R.E. A new tinman-related gene, nkx2.7, anticipates the expression of nkx2.5 and nkx2.3 in zebrafish heart and pharyngeal endoderm. Dev. Biol. 1996. [Google Scholar] [CrossRef] [Green Version]
- Azpiazu, N.; Frasch, M. tinman and bagpipe: Two homeo box genes that determine cell fates in the dorsal mesoderm of Drosophila. Genes Dev. 1993, 7, 1325–1340. [Google Scholar] [CrossRef]
- Harvey, R.P. REVIEW NK-2 Homeobox Genes and Heart Development. Dev. Biol. 1996, 178, 203–216. [Google Scholar] [CrossRef] [Green Version]
- Hinits, Y.; Pan, L.; Walker, C.; Dowd, J.; Moens, C.B.; Hughes, S.M. Zebrafish Mef2ca and Mef2cb are essential for both first and second heart field cardiomyocyte differentiation. Dev. Biol. 2012, 369, 199–210. [Google Scholar] [CrossRef] [Green Version]
- Ticho, B.S.; Stainier, D.Y.R.; Fishman, M.C.; Breitbart, R.E. Three zebrafish MEF2 genes delineate somitic and cardiac muscle development in wild-type and mutant embryos. Mech. Dev. 1996, 59, 205–218. [Google Scholar] [CrossRef]
- Bahary, N.; Goishi, K.; Stuckenholz, C.; Weber, G.; Leblanc, J.; Schafer, C.A.; Berman, S.S.; Klagsbrun, M.; Zon, L.I. Duplicate VegfA genes and orthologues of the KDR receptor tyrosine kinase family mediate vascular development in the zebrafish. Blood 2007, 110, 3627–3636. [Google Scholar] [CrossRef] [PubMed]
- Jin, H.; Xu, J.; Qian, F.; Du, L.; Tan, C.Y.; Lin, Z.; Peng, J.; Wen, Z. The 5’ zebrafish scl promoter targets transcription to the brain, spinal cord, and hematopoietic and endothelial progenitors. Dev. Dyn. 2006, 235, 60–67. [Google Scholar] [CrossRef] [PubMed]
- Lieschke, G.J.; Oates, A.C.; Paw, B.H.; Thompson, M.A.; Hall, N.E.; Ward, A.C.; Ho, R.K.; Zon, L.I.; Layton, J.E. Zebrafish SPI-1 (PU.1) marks a site of myeloid development independent of primitive erythropoiesis: Implications for axial patterning. Dev. Biol. 2002, 246, 274–295. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lints, T.J.; Parsons, L.M.; Hartley, L.; Lyons, I.; Harvey, R.P. Nkx-2.5: A novel murine homeobox gene expressed in early heart progenitor cells and their myogenic descendants. Development 1993, 119, 969. [Google Scholar]
- Reifers, F.; Walsh, E.C.; Léger, S.; Stainier, D.Y.R.; Brand, M.; Leger, S.; Stainier, D.Y.R.; Brand, M. Induction and differentiation of the zebrafish heart requires fibroblast growth factor 8 (fgf8/acerebellar). Development 2000, 127, 225–235. [Google Scholar] [PubMed]
- Herbomel, P.; Thisse, B.; Thisse, C. Ontogeny and behaviour of early macrophages in the zebrafish embryo. Development 1999, 126, 3735–3745. [Google Scholar]
- Bussmann, J.; Bakkers, J.; Schulte-Merker, S. Early endocardial morphogenesis requires Scl/Tal1. PLoS Genet. 2007, 3, e140. [Google Scholar] [CrossRef]
- Misfeldt, A.M.; Boyle, S.C.; Tompkins, K.L.; Bautch, V.L.; Labosky, P.A.; Baldwin, H.S. Endocardial cells are a distinct endothelial lineage derived from Flk1+ multipotent cardiovascular progenitors. Dev Biol 2009, 333, 78–89. [Google Scholar] [CrossRef] [Green Version]
- Vogeli, K.M.; Jin, S.W.; Martin, G.R.; Stainier, D.Y. A common progenitor for haematopoietic and endothelial lineages in the zebrafish gastrula. Nature 2006, 443, 337–339. [Google Scholar] [CrossRef]
- Miura, G.I.; Yelon, D. A Guide to Analysis of Cardiac Phenotypes in the Zebrafish Embryo; Academic Press: Cambridge, MA, USA, 2011; Volume 101. [Google Scholar]
- Yelon, D.; Horne, S.A.; Stainier, D.Y.R. Restricted Expression of Cardiac Myosin Genes Reveals Regulated Aspects of Heart Tube Assembly in Zebrafish. Dev. Biol. 1999, 214, 23–37. [Google Scholar] [CrossRef] [Green Version]
- Berdougo, E.; Coleman, H.; Lee, D.H.; Stainier, D.Y.R.; Yelon, D. Mutation of weak atrium/atrial myosin heavy chain disrupts atrial function and influences ventricular morphogenesis in zebrafish. Development 2003, 130, 6121–6129. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rohr, S.; Otten, C.; Abdelilah-Seyfried, S. Asymmetric Involution of the Myocardial Field Drives Heart Tube Formation in Zebrafish. Circ. Res. 2008, 102, e12–e19. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kikuchi, Y.; Agathon, A.; Alexander, J.; Thisse, C.; Waldron, S.; Yelon, D.; Thisse, B.; Stainier, D.Y. casanova encodes a novel Sox-related protein necessary and sufficient for early endoderm formation in zebrafish. Genes Dev. 2001, 15. [Google Scholar] [CrossRef] [Green Version]
- Alexander, J.; Stainier, D.Y. A molecular pathway leading to endoderm formation in zebrafish. Curr. Biol. 1999, 9, 1147–1157. [Google Scholar] [CrossRef] [Green Version]
- Kikuchi, Y.; Trinh, L.A.; Reiter, J.F.; Alexander, J.; Yelon, D.; Stainier, D.Y. The zebrafish bonnie and clyde gene encodes a Mix family homeodomain protein that regulates the generation of endodermal precursors. Genes Dev. 2000, 14, 1279–1289. [Google Scholar] [PubMed]
- Bloomekatz, J.; Singh, R.; Prall, O.W.; Dunn, A.C.; Vaughan, M.; Loo, C.-S.; Harvey, R.P.; Yelon, D. Platelet-derived growth factor (PDGF) signaling directs cardiomyocyte movement toward the midline during heart tube assembly. eLife 2017, 6, e21172. [Google Scholar] [CrossRef] [Green Version]
- Burkhard, S.B.; Bakkers, J. Spatially resolved RNA-sequencing of the embryonic heart identifies a role for Wnt/β-catenin signaling in autonomic control of heart rate. eLife 2018, 7, e31515. [Google Scholar] [CrossRef] [PubMed]
- Guerra, A.; Germano, R.F.; Stone, O.; Arnaout, R.; Guenther, S.; Ahuja, S.; Uribe, V.; Vanhollebeke, B.; Stainier, D.Y.; Reischauer, S. Distinct myocardial lineages break atrial symmetry during cardiogenesis in zebrafish. eLife 2018, 7, e32833. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liu, J.; Bressan, M.; Hassel, D.; Huisken, J.; Staudt, D.; Kikuchi, K.; Poss, K.D.; Mikawa, T.; Stainier, D.Y.R. A dual role for ErbB2 signaling in cardiac trabeculation. Development 2010, 137, 3867–3875. [Google Scholar] [CrossRef] [Green Version]
- Liu, J.; Stainier, D.Y.R. Tbx5 and Bmp Signaling Are Essential for Proepicardium Specification in Zebrafish. Circ. Res. 2010, 106, 1818. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Serluca, F.C. Development of the proepicardial organ in the zebrafish. Dev. Biol. 2008, 315, 18–27. [Google Scholar] [CrossRef] [Green Version]
- Grimes, A.C.; Stadt, H.A.; Shepherd, I.T.; Kirby, M.L. Solving an enigma: Arterial pole development in the zebrafish heart. Dev. Biol. 2006, 290, 265–276. [Google Scholar] [CrossRef] [Green Version]
- Peshkovsky, C.; Totong, R.; Yelon, D. Dependence of cardiac trabeculation on neuregulin signaling and blood flow in zebrafish. Dev. Dyn. 2011, 240, 446–456. [Google Scholar] [CrossRef]
- Scherz, P.J.; Huisken, J.; Sahai-Hernandez, P.; Stainier, D.Y.R. High-speed imaging of developing heart valves reveals interplay of morphogenesis and function. Development 2008, 135, 1179–1187. [Google Scholar] [CrossRef] [Green Version]
- Staudt, D.W.; Liu, J.; Thorn, K.S.; Stuurman, N.; Liebling, M.; Stainier, D.Y.R. High-resolution imaging of cardiomyocyte behavior reveals two distinct steps in ventricular trabeculation. Development 2014, 141, 585–593. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Peralta, M.; González-Rosa, J.M.; Marques, I.J.; Mercader, N. The Epicardium in the Embryonic and Adult Zebrafish. J. Dev. Biol. 2014, 2, 101–116. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Huisken, J.; Stainier, D.Y.R. Selective plane illumination microscopy techniques in developmental biology. Development 2009, 136, 1963–1975. [Google Scholar] [CrossRef] [Green Version]
- Kwan, K.M.; Fujimoto, E.; Grabher, C.; Mangum, B.D.; Hardy, M.E.; Campbell, D.S.; Parant, J.M.; Yost, H.J.; Kanki, J.P.; Chien, C. Bin The Tol2kit: A multisite gateway-based construction kit for Tol2 transposon transgenesis constructs. Dev. Dyn. 2007, 236, 3088–3099. [Google Scholar] [CrossRef] [PubMed]
- Villefranc, J.A.; Amigo, J.; Lawson, N.D. Gateway compatible vectors for analysis of gene function in the zebrafish. Dev Dyn. 2007, 236, 3077–3087. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mosimann, C.; Kaufman, C.K.; Li, P.; Pugach, E.K.; Tamplin, O.J.; Zon, L.I. Ubiquitous transgene expression and Cre-based recombination driven by the ubiquitin promoter in zebrafish. Development 2011, 138, 169–177. [Google Scholar] [CrossRef] [Green Version]
- Hans, S.; Kaslin, J.; Freudenreich, D.; Brand, M. Temporally-controlled site-specific recombination in zebrafish. PLoS ONE 2009, 4, e4640. [Google Scholar] [CrossRef] [Green Version]
- Kawakami, K. Transposon tools and methods in zebrafish. Dev. Dyn. 2005, 234, 244–254. [Google Scholar] [CrossRef]
- Sánchez-Iranzo, H.; Galardi-Castilla, M.; Minguillón, C.; Sanz-Morejón, A.; González-Rosa, J.M.J.M.; Felker, A.; Ernst, A.; Guzmán-Martínez, G.; Mosimann, C.; Mercader, N. Tbx5a lineage tracing shows cardiomyocyte plasticity during zebrafish heart regeneration. Nat. Commun. 2018, 9, 428. [Google Scholar] [CrossRef] [Green Version]
- Prummel, K.D.; Crowell, H.L.; Nieuwenhuize, S.; Brombacher, E.C.; Daetwyler, S.; Soneson, C.; Kresoja-Rakic, J.; Ronner, M.; Kocere, A.; Ernst, A.; et al. Hand2 delineates mesothelium progenitors and is reactivated in mesothelioma. bioRxiv 2020. [Google Scholar] [CrossRef]
- Devine, W.P.; Wythe, J.D.; George, M.; Koshiba-Takeuchi, K.; Bruneau, B.G. Early patterning and specification of cardiac progenitors in gastrulating mesoderm. eLife 2014, 3, e03848. [Google Scholar] [CrossRef] [PubMed]
- Yuan, X.; Song, M.; Devine, P.; Bruneau, B.G.; Scott, I.C.; Wilson, M.D. Heart enhancers with deeply conserved regulatory activity are established early in zebrafish development. Nat. Commun. 2018, 9, 4977. [Google Scholar] [CrossRef] [Green Version]
- Huang, C.J.; Tu, C.T.; Hsiao, C.D.; Hsieh, F.J.; Tsai, H.J. Germ-line transmission of a myocardium-specific GFP transgene reveals critical regulatory elements in the cardiac myosin light chain 2 promoter of zebrafish. Dev. Dyn. 2003, 228, 30–40. [Google Scholar] [CrossRef] [PubMed]
- Zhou, Y.; Cashman, T.J.T.J.; Nevis, K.R.K.R.; Obregon, P.; Carney, S.A.S.A.; Liu, Y.; Gu, A.; Mosimann, C.; Sondalle, S.; Peterson, R.E.R.E.; et al. Latent TGF-beta binding protein 3 identifies a second heart field in zebrafish. Nature 2011, 474, 645–648. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Felker, A.; Prummel, K.D.; Merks, A.M.; Mickoleit, M.; Brombacher, E.C.; Huisken, J.; Panáková, D.; Mosimann, C. Continuous addition of progenitors forms the cardiac ventricle in zebrafish. Nat. Commun. 2018, 9. [Google Scholar] [CrossRef] [Green Version]
- Lawson, N.D.; Weinstein, B.M. In vivo imaging of embryonic vascular development using transgenic zebrafish. Dev. Biol. 2002, 248, 307–318. [Google Scholar] [CrossRef] [Green Version]
- Mably, J.D.; Mohideen, M.A.P.K.; Burns, C.G.; Chen, J.N.; Fishman, M.C. Heart of glass Regulates the Concentric Growth of the Heart in Zebrafish. Curr. Biol. 2003, 13, 2138–2147. [Google Scholar] [CrossRef]
- de Pater, E.; Clijsters, L.; Marques, S.R.; Lin, Y.-F.F.; Garavito-Aguilar, Z.V.; Yelon, D.; Bakkers, J. Distinct phases of cardiomyocyte differentiation regulate growth of the zebrafish heart. Development 2009, 136, 1633–1641. [Google Scholar] [CrossRef] [Green Version]
- Lazic, S.; Scott, I.C. Mef2cb regulates late myocardial cell addition from a second heart field-like population of progenitors in zebrafish. Dev. Biol. 2011, 354, 123–133. [Google Scholar] [CrossRef] [PubMed]
- Kikuchi, K.; Holdway, J.E.; Major, R.J.; Blum, N.; Dahn, R.D.; Begemann, G.; Poss, K.D. Short Article Retinoic Acid Production by Endocardium and Epicardium Is an Injury Response Essential for Zebrafish Heart Regeneration. Dev. Cell 2011, 20, 397–404. [Google Scholar] [CrossRef] [Green Version]
- Yin, C.; Kikuchi, K.; Hochgreb, T.; Poss, K.D.; Stainier, D.Y.R. Hand2 regulates extracellular matrix remodeling essential for gut-looping morphogenesis in zebrafish. Dev. Cell 2010, 18, 973–984. [Google Scholar] [CrossRef] [Green Version]
- Buckingham, M.; Meilhac, S.; Zaffran, S. Building the mammalian heart from two sources of myocardial cells. Nat. Rev. Genet. 2005, 6, 826–835. [Google Scholar] [CrossRef] [PubMed]
- Stolfi, A.; Gainous, T.B.; Young, J.J.; Mori, A.; Levine, M.; Christiaen, L. Early chordate origins of the vertebrate second heart field. Science 2010, 329, 565–568. [Google Scholar] [CrossRef] [Green Version]
- Zaffran, S.; Kelly, R.G. New developments in the second heart field. Differentiation 2012, 84, 17–24. [Google Scholar] [CrossRef]
- Black, B.L. Transcriptional pathways in second heart field development. Semin. Cell Dev. Biol. 2007, 18, 67–76. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hami, D.; Grimes, A.C.; Tsai, H.J.; Kirby, M.L. Zebrafish cardiac development requires a conserved secondary heart field. Development 2011, 138, 2389–2398. [Google Scholar] [CrossRef] [Green Version]
- Ivanovitch, K.; Temiño, S.; Torres, M. Live imaging of heart tube development in mouse reveals alternating phases of cardiac differentiation and morphogenesis. eLife 2017, 6. [Google Scholar] [CrossRef] [PubMed]
- McDole, K.; Guignard, L.; Amat, F.; Berger, A.; Malandain, G.; Royer, L.A.; Turaga, S.C.; Branson, K.; Keller, P.J. In Toto Imaging and Reconstruction of Post-Implantation Mouse Development at the Single-Cell Level. Cell 2018, 175, 859–876. [Google Scholar] [CrossRef] [Green Version]
- Kidokoro, H.; Yonei-Tamura, S.; Tamura, K.; Schoenwolf, G.C.; Saijoh, Y. The heart tube forms and elongates through dynamic cell rearrangement coordinated with foregut extension. Development 2018, 145. [Google Scholar] [CrossRef] [Green Version]
- Kaplan, N.; Razy-Krajka, F.; Christiaen, L. Regulation and evolution of cardiopharyngeal cell identity and behavior: Insights from simple chordates. Curr. Opin. Genet. Dev. 2015, 32, 119–128. [Google Scholar] [CrossRef] [Green Version]
- Simoes, F.C.; Peterkin, T.; Patient, R.; Simões, F.C.; Peterkin, T.; Patient, R. Fgf differentially controls cross-antagonism between cardiac and haemangioblast regulators. Development 2011, 138, 3235–3245. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dobrzycki, T.; Lalwani, M.; Telfer, C.; Monteiro, R.; Patient, R. The roles and controls of GATA factors in blood and cardiac development. IUBMB Life 2020, 72, 39–44. [Google Scholar] [CrossRef]
- Marques, S.R.; Lee, Y.; Poss, K.D.; Yelon, D. Reiterative roles for FGF signaling in the establishment of size and proportion of the zebrafish heart. Dev. Biol. 2008, 321, 397–406. [Google Scholar] [CrossRef] [Green Version]
- Rasouli, S.J.; El-Brolosy, M.; Tsedeke, A.T.; Bensimon-Brito, A.; Ghanbari, P.; Maischein, H.M.; Kuenne, C.; Stainier, D.Y. The flow responsive transcription factor Klf2 is required for myocardial wall integrity by modulating Fgf signaling. eLife 2018, 7. [Google Scholar] [CrossRef]
- Pradhan, A.; Zeng, X.-X.I.; Sidhwani, P.; Marques, S.R.; George, V.; Targoff, K.L.; Chi, N.C.; Yelon, D. FGF signaling enforces cardiac chamber identity in the developing ventricle. Development 2017, 144, 1328–1338. [Google Scholar] [CrossRef] [Green Version]
- Gibb, N.; Lazic, S.; Yuan, X.; Deshwar, A.R.; Leslie, M.; Wilson, M.D.; Scott, I.C. Hey2 regulates the size of the cardiac progenitor pool during vertebrate heart development. Development 2018. [Google Scholar] [CrossRef] [Green Version]
- Rydeen, A.A.B.; Waxman, J.J.S.; Bruneau, B.; Bruneau, B.; Buckingham, M.; Meilhac, S.; Zaffran, S.; Abu-Issa, R.; Kirby, M.; Lazic, S.; et al. Cyp26 Enzymes Facilitate Second Heart Field Progenitor Addition and Maintenance of Ventricular Integrity. PLOS Biol. 2016, 14, e2000504. [Google Scholar] [CrossRef]
- Perl, E.; Waxman, J.S. Reiterative mechanisms of retinoic acid signaling during vertebrate heart development. J. Dev. Biol. 2019, 2019, 11. [Google Scholar] [CrossRef] [Green Version]
- Sorrell, M.R.J.; Waxman, J.S. Restraint of Fgf8 signaling by retinoic acid signaling is required for proper heart and forelimb formation. Dev. Biol. 2011. [Google Scholar] [CrossRef] [Green Version]
- Schier, A.F.; Talbot, W.S. Molecular genetics of axis formation in zebrafish. Annu. Rev. Genet. 2005, 39, 561–613. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lombardo, V.A.; Heise, M.; Moghtadaei, M.; Bornhorst, D.; Männer, J.; Abdelilah-Seyfried, S. Morphogenetic control of zebrafish cardiac looping by Bmp signaling. Development 2019, 146. [Google Scholar] [CrossRef] [PubMed]
- Lenhart, K.F.; Holtzman, N.G.; Williams, J.R.; Burdine, R.D. Integration of nodal and BMP signals in the heart requires FoxH1 to create left-right differences in cell migration rates that direct cardiac asymmetry. PLoS Genet. 2013, 9, e1003109. [Google Scholar] [CrossRef] [Green Version]
- Andrés-Delgado, L.; Galardi-Castilla, M.; Münch, J.; Peralta, M.; Ernst, A.; González-Rosa, J.M.; Tessadori, F.; Santamaría, L.; Bakkers, J.; Vermot, J.; et al. Notch and Bmp signaling pathways act coordinately during the formation of the proepicardium. Dev. Dyn. 2020, 249, 1455–1469. [Google Scholar] [CrossRef]
- Fischer, P.; Chen, H.; Pacho, F.; Rieder, D.; Kimmel, R.A.; Meyer, D. FoxH1 represses miR-430 during early embryonic development of zebrafish via non-canonical regulation. BMC Biol. 2019, 17, 61. [Google Scholar] [CrossRef]
- Nelson, A.C.; Cutty, S.J.; Niini, M.; Stemple, D.L.; Flicek, P.; Houart, C.; Bruce, A.E.E.; Wardle, F.C. Global identification of Smad2 and Eomesodermin targets in zebrafish identifies a conserved transcriptional network in mesendoderm and a novel role for Eomesodermin in repression of ectodermal gene expression. BMC Biol. 2014, 12, 81. [Google Scholar] [CrossRef] [Green Version]
- Pogoda, H.M.; Solnica-Krezel, L.; Driever, W.; Meyer, D. The zebrafish forkhead transcription factor FoxH1/Fast1 is a modulator of nodal signaling required for organizer formation. Curr. Biol. 2000, 10, 1041–1049. [Google Scholar] [CrossRef] [Green Version]
- Guner-Ataman, B.; Paffett-Lugassy, N.; Adams, M.S.; Nevis, K.R.; Jahangiri, L.; Obregon, P.; Kikuchi, K.; Poss, K.D.; Burns, C.E.; Burns, C.G. Zebrafish second heart field development relies on progenitor specification in anterior lateral plate mesoderm and nkx2.5 function. Development 2013, 140, 1353–1363. [Google Scholar] [CrossRef] [Green Version]
- Ueno, S.; Weidinger, G.; Osugi, T.; Kohn, A.D.; Golob, J.L.; Pabon, L.; Reinecke, H.; Moon, R.T.; Murry, C.E. Biphasic role for Wnt/β-catenin signaling in cardiac specification in zebrafish and embryonic stem cells. Proc. Natl. Acad. Sci. USA 2007, 104, 9685–9690. [Google Scholar] [CrossRef] [Green Version]
- Bakre, M.M.; Hoi, A.; Mong, J.C.Y.; Koh, Y.Y.; Wong, K.Y.; Stanton, L.W. Generation of multipotential mesendodermal progenitors from mouse embryonic stem cells via sustained Wnt pathway activation. J. Biol. Chem. 2007, 282, 31703–31712. [Google Scholar] [CrossRef] [Green Version]
- Woll, P.S.; Morris, J.K.; Painschab, M.S.; Marcus, R.K.; Kohn, A.D.; Biechele, T.L.; Moon, R.T.; Kaufman, D.S. Wnt signaling promotes hematoendothelial cell development from human embryonic stem cells. Blood 2008, 111, 122–131. [Google Scholar] [CrossRef] [Green Version]
- Gessert, S.; Kühl, M.; Kühl, M. The multiple phases and faces of wnt signaling during cardiac differentiation and development. Circ. Res. 2010, 107, 186–199. [Google Scholar] [CrossRef] [PubMed]
- Cohen, E.D.; Tian, Y.; Morrisey, E.E. Wnt signaling: An essential regulator of cardiovascular differentiation, morphogenesis and progenitor self-renewal. Development 2008, 135, 789. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mandal, A.; Holowiecki, A.; Song, Y.C.; Waxman, J.S. Wnt signaling balances specification of the cardiac and pharyngeal muscle fields. Mech. Dev. 2017, 143, 32–41. [Google Scholar] [CrossRef] [PubMed]
- Verhoeven, M.C.; Haase, C.; Christoffels, V.M.; Weidinger, G.; Bakkers, J. Wnt signaling regulates atrioventricular canal formation upstream of BMP and Tbx2. Birth Defects Res. Part A Clin. Mol. Teratol. 2011, 91, 435–440. [Google Scholar] [CrossRef]
- Hurlstone, A.F.L.; Haramis, A.-P.G.; Wienholds, E.; Begthel, H.; Korving, J.; van Eeden, F.; Cuppen, E.; Zivkovic, D.; Plasterk, R.H.A.; Clevers, H. The Wnt/β-catenin pathway regulates cardiac valve formation. Nature 2003, 425, 633–637. [Google Scholar] [CrossRef] [PubMed]
- Matsui, T.; Raya, Á.; Kawakami, Y.; Callol-Massot, C.; Capdevila, J.; Rodríguez-Esteban, C.; Belmonte, J.C.I. Noncanonical Wnt signaling regulates midline convergence of organ primordia during zebrafisn development. Genes Dev. 2005. [Google Scholar] [CrossRef] [Green Version]
- Panakova, D.; Werdich, A.A.; Macrae, C.A. Wnt11 patterns a myocardial electrical gradient through regulation of the L-type Ca(2+) channel. Nature 2010, 466, 874–878. [Google Scholar] [CrossRef] [Green Version]
- Huang, P.; Schier, A.F. Dampened Hedgehog signaling but normal Wnt signaling in zebrafish without cilia. Development 2009. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Schilling, T.F.; Concordet, J.P.; Ingham, P.W. Regulation of left-right asymmetries in the zebrafish by Shh and BMP4. Dev. Biol. 1999. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Thomas, N.A.; Koudijs, M.; van Eeden, F.J.M.; Joyner, A.L.; Yelon, D. Hedgehog signaling plays a cell-autonomous role in maximizing cardiac developmental potential. Development 2008, 135, 3789–3799. [Google Scholar] [CrossRef] [Green Version]
- Wong, K.S.; Rehn, K.; Palencia-Desai, S.; Kohli, V.; Hunter, W.; Uhl, J.D.; Rost, M.S.; Sumanas, S. Hedgehog signaling is required for differentiation of endocardial progenitors in zebrafish. Dev. Biol. 2012. [Google Scholar] [CrossRef] [Green Version]
- Sugimoto, K.; Hui, S.P.; Sheng, D.Z.; Kikuchi, K. Dissection of zebrafish shha function using site-specific targeting with a Cre-dependent genetic switch. eLife 2017. [Google Scholar] [CrossRef] [Green Version]
- Zhou, L.; Liu, J.; Xiang, M.; Olson, P.; Guzzetta, A.; Zhang, K.; Moskowitz, I.P.; Xie, L. Gata4 potentiates second heart field proliferation and Hedgehog signaling for cardiac septation. Proc. Natl. Acad. Sci. USA 2017, 201605137. [Google Scholar] [CrossRef] [Green Version]
- Stennard, F.A.; Harvey, R.P. T-box Transcription Factors and Their Roles in Regulatory Hierarchies in the Developing Heart; The Company of Biologists Ltd.: Cambridge, UK, 2005; Volume 132, pp. 4897–4910. [Google Scholar]
- Nevis, K.; Obregon, P.; Walsh, C.; Guner-Ataman, B.; Burns, C.G.; Burns, C.E. Tbx1 is required for second heart field proliferation in zebrafish. Dev. Dyn. 2013, 242, 550–559. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Piotrowski, T.; Ahn, D.; Schilling, T.F.; Nair, S.; Ruvinsky, I.; Geisler, R.; Rauch, G.-J.; Haffter, P.; Zon, L.I.; Zhou, Y.; et al. The zebrafish van gogh mutation disrupts tbx1, which is involved in the DiGeorge deletion syndrome in humans. Development 2003, 130, 5043–5052. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Naiche, L.A.; Harrelson, Z.; Kelly, R.G.; Papaioannou, V.E. T-box genes in vertebrate development. Annu. Rev. Genet. 2005, 39, 219–239. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, Z.; Baldini, A. Manipulation of endogenous regulatory elements and transgenic analyses of the Tbx1 gene. Mamm. Genome 2010, 21, 556–564. [Google Scholar] [CrossRef] [Green Version]
- Song, Y.C.; Dohn, T.E.; Rydeen, A.B.; Nechiporuk, A.V.; Waxman, J.S. HDAC1-mediated repression of the retinoic acid-responsive gene ripply3 promotes second heart field development. PLoS Genet. 2019. [Google Scholar] [CrossRef]
- Mori, A.D.; Bruneau, B.G. TBX5 mutations and congenital heart disease: Holt-Oram syndrome revealed. Curr. Opin. Cardiol. 2004, 19, 211–215. [Google Scholar] [CrossRef]
- Ahn, D.G.; Kourakis, M.J.; Rohde, L.A.; Silver, L.M.; Ho, R.K. T-box gene tbx5 is essential for formation of the pectoral limb bud. Nature 2002, 417, 754–758. [Google Scholar] [CrossRef]
- Parrie, L.E.; Renfrew, E.M.; Wal, A.V.; Mueller, R.L.; Garrity, D.M. Zebrafish tbx5 paralogs demonstrate independent essential requirements in cardiac and pectoral fin development. Dev. Dyn. 2013, 242, 485–502. [Google Scholar] [CrossRef]
- Chiavacci, E.; Cremisi, F.; Guzzolino, E.; Cellerino, A.; Baumgart, M.; D’aurizio, R.; Pitto, L. miR-19a replacement rescues cardiac and fin defects in zebrafish model of holt-oram syndrome. Cardiovasc. Res. 2014, 103. [Google Scholar] [CrossRef] [Green Version]
- Holtzinger, A.; Evans, T. Gata5 and Gata6 are functionally redundant in zebrafish for specification of cardiomyocytes. Dev. Biol. 2007, 312, 613–622. [Google Scholar] [CrossRef] [Green Version]
- Jiang, Y.; Drysdale, T.A.; Evans, T. A Role for GATA-4/5/6 in the Regulation of Nkx2.5 Expression with Implications for Patterning of the Precardiac Field. Dev. Biol. 1999, 216, 57–71. [Google Scholar] [CrossRef] [Green Version]
- Reiter, J.F.; Alexander, J.; Rodaway, A.; Yelon, D.; Patient, R.; Holder, N.; Stainier, D.Y. Gata5 is required for the development of the heart and endoderm in zebrafish. Genes Dev. 1999, 13, 2983–2995. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sam, J.; Mercer, E.J.; Torregroza, I.; Banks, K.M.; Evans, T. Specificity, redundancy and dosage thresholds among gata4/5/6 genes during zebrafish cardiogenesis. Biol. Open 2020, 9. [Google Scholar] [CrossRef]
- Lou, X.; Deshwar, A.R.; Crump, J.G.; Scott, I.C. Smarcd3b and Gata5 promote a cardiac progenitor fate in the zebrafish embryo. Development 2011, 138, 3113–3123. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dodou, E.; Verzi, M.P.; Anderson, J.P.; Xu, S.-M.; Black, B.L. Mef2c is a direct transcriptional target of ISL1 and GATA factors in the anterior heart field during mouse embryonic development. Development 2004, 131, 3931–3942. [Google Scholar] [CrossRef] [Green Version]
- Arceci, R.J.; King, A.A.; Simon, M.C.; Orkin, S.H.; Wilson, D.B. Mouse GATA-4: A retinoic acid-inducible GATA-binding transcription factor expressed in endodermally derived tissues and heart. Mol. Cell. Biol. 1993, 13, 2235–2246. [Google Scholar] [CrossRef] [Green Version]
- Molkentin, J.D.; Antos, C.; Mercer, B.; Taigen, T.; Miano, J.M.; Olson, E.N. Direct Activation of a GATA6 Cardiac Enhancer by Nkx2.5: Evidence for a Reinforcing Regulatory Network of Nkx2.5 and GATA Transcription Factors in the Developing Heart. Dev. Biol. 2000, 217, 301–309. [Google Scholar] [CrossRef]
- Stainier, D.Y.R. A glimpse into the molecular entrails of endoderm formation. Genes Dev. 2002, 16, 893–907. [Google Scholar] [CrossRef] [Green Version]
- Song, M.; Yuan, X.; Racioppi, C.; Leslie, M.; Aleksandrova, A.; Christiaen, L.; Wilson, M.D.; Scott, I.C. GATA4/5/6 family transcription factors are conserved determinants of cardiac versus pharyngeal mesoderm fate. bioRxiv 2020. [Google Scholar] [CrossRef]
- Evans, A.L.; Faial, T.; Gilchrist, M.J.; Down, T.; Vallier, L.; Pedersen, R.A.; Wardle, F.C.; Smith, J.C. Genomic targets of Brachyury (T) in differentiating mouse embryonic stem cells. PLoS ONE 2012, 7, e33346. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- George, V.; Colombo, S.; Targoff, K.L. An early requirement for nkx2.5 Ensures first and Second heart field ventricular identity and cardiac function into adulthood. Dev. Biol. 2014. [Google Scholar] [CrossRef] [Green Version]
- Targoff, K.L.; Colombo, S.; George, V.; Schell, T.; Kim, S.-H.; Solnica-Krezel, L.; Yelon, D. Nkx genes are essential for maintenance of ventricular identity. Development 2013, 140, 4203–4213. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Holowiecki, A.; Linstrum, K.; Ravisankar, P.; Chetal, K.; Salomonis, N.; Waxman, J.S. Pbx4 limits heart size and fosters arch artery formation by partitioning second heart field progenitors and restricting proliferation. Development 2020, 147. [Google Scholar] [CrossRef]
- Holland, N.D.; Venkatesh, T.V.; Holland, L.Z.; Jacobs, D.K.; Bodmer, R. AmphiNk2-tin, an amphioxus homeobox gene expressed in myocardial progenitors: Insights into evolution of the vertebrate heart. Dev. Biol. 2003, 255, 128–137. [Google Scholar] [CrossRef] [Green Version]
- Newman, C.S.; Krieg, P.A. tinman-related genes expressed during heart development inXenopus. Dev. Genet. 1998, 22, 230–238. [Google Scholar] [CrossRef]
- Cai, C.-L.L.; Liang, X.; Shi, Y.; Chu, P.-H.H.; Pfaff, S.L.; Chen, J.; Evans, S. Isl1 identifies a cardiac progenitor population that proliferates prior to differentiation and contributes a majority of cells to the heart. Dev. Cell 2003, 5, 877–889. [Google Scholar] [CrossRef] [Green Version]
- Kwon, C.; Qian, L.; Cheng, P.; Nigam, V.; Arnold, J.; Srivastava, D. A regulatory pathway involving Notch1/β-catenin/Isl1 determines cardiac progenitor cell fate. Nat. Cell Biol. 2009, 11, 951–957. [Google Scholar] [CrossRef] [Green Version]
- Nathan, E.; Monovich, A.; Tirosh-Finkel, L.; Harrelson, Z.; Rousso, T.; Rinon, A.; Harel, I.; Evans, S.M.; Tzahor, E. The contribution of Islet1-expressing splanchnic mesoderm cells to distinct branchiomeric muscles reveals significant heterogeneity in head muscle development. Development 2008, 135, 647–657. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kelly, R.G.; Buckingham, M.E.; Moorman, A.F. Heart fields and cardiac morphogenesis. Cold Spring Harb. Perspect. Med. 2014, 4. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Witzel, H.R.R.; Jungblut, B.; Choe, C.P.P.; Crump, J.G.G.; Braun, T.; Dobreva, G. The LIM protein Ajuba restricts the second heart field progenitor pool by regulating Isl1 activity. Dev. Cell 2012, 23, 58–70. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Glasauer, S.M.K.; Neuhauss, S.C.F. Whole-genome duplication in teleost fishes and its evolutionary consequences. Mol. Genet. Genomics 2014, 289, 1045–1060. [Google Scholar] [CrossRef] [Green Version]
- Witzel, H.R.; Cheedipudi, S.; Gao, R.; Stainier, D.Y.R.; Dobreva, G.D. Isl2b regulates anterior second heart field development in zebrafish. Sci. Rep. 2017, 7, 41043. [Google Scholar] [CrossRef]
- Caputo, L.; Witzel, H.R.; Kolovos, P.; Cheedipudi, S.; Looso, M.; Mylona, A.; van IJcken, W.F.J.; Laugwitz, K.-L.; Evans, S.M.; Braun, T.; et al. The Isl1/Ldb1 Complex Orchestrates Genome-wide Chromatin Organization to Instruct Differentiation of Multipotent Cardiac Progenitors. Cell Stem Cell 2015, 17, 287–299. [Google Scholar] [CrossRef] [Green Version]
- Colombo, S.; De Sena-Tomaś, C.; George, V.; Werdich, A.A.; Kapur, S.; Macrae, C.A.; Targoff, K.L. Nkx genes establish second heart field cardiomyocyte progenitors at the arterial pole and pattern the venous pole through isl1 repression. Development 2018. [Google Scholar] [CrossRef] [Green Version]
- Meilhac, S.M.; Esner, M.; Kelly, R.G.; Nicolas, J.-F.; Buckingham, M.E. The Clonal Origin of Myocardial Cells in Different Regions of the Embryonic Mouse Heart. Dev. Cell 2004, 6, 685–698. [Google Scholar] [CrossRef] [Green Version]
- Gans, C.; Northcutt, R.G. Neural Crest and the Origin of Vertebrates: A New Head. Science 1983, 220, 268–273. [Google Scholar] [CrossRef]
- Diogo, R.; Kelly, R.G.; Christiaen, L.; Levine, M.; Ziermann, J.M.; Molnar, J.L.; Noden, D.M.; Tzahor, E. A new heart for a new head in vertebrate cardiopharyngeal evolution. Nature 2015, 520, 466–473. [Google Scholar] [CrossRef] [Green Version]
- Tirosh-Finkel, L.; Elhanany, H.; Rinon, A.; Tzahor, E. Mesoderm progenitor cells of common origin contribute to the head musculature and the cardiac outflow tract. Development 2006, 133, 1943–1953. [Google Scholar] [CrossRef] [Green Version]
- Bothe, I.; Dietrich, S. The molecular setup of the avian head mesoderm and its implication for craniofacial myogenesis. Dev. Dyn. 2006, 235, 2845–2860. [Google Scholar] [CrossRef] [Green Version]
- Kokubo, N.; Matsuura, M.; Onimaru, K.; Tiecke, E.; Kuraku, S.; Kuratani, S.; Tanaka, M. Mechanisms of heart development in the Japanese lamprey, Lethenteron japonicum. Evol. Dev. 2010, 12, 34–44. [Google Scholar] [CrossRef]
- Onimaru, K.; Shoguchi, E.; Kuratani, S.; Tanaka, M. Development and evolution of the lateral plate mesoderm: Comparative analysis of amphioxus and lamprey with implications for the acquisition of paired fins. Dev. Biol. 2011, 359, 124–136. [Google Scholar] [CrossRef] [Green Version]
- Sauka-Spengler, T.; Le Mentec, C.; Lepage, M.; Mazan, S. Embryonic expression of Tbx1, a DiGeorge syndrome candidate gene, in the lamprey Lampetra fluviatilis. Gene Expr. Patterns 2002, 2, 99–103. [Google Scholar] [CrossRef]
- Tiecke, E.; Matsuura, M.; Kokubo, N.; Kuraku, S.; Kusakabe, R.; Kuratani, S.; Tanaka, M. Identification and developmental expression of two Tbx1/10-related genes in the agnathan Lethenteron japonicum. Dev. Genes Evol. 2007, 217, 691–697. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, W.; Niu, X.; Stuart, T.; Jullian, E.; Mauck, W.M.; Kelly, R.G.; Satija, R.; Christiaen, L. A single-cell transcriptional roadmap for cardiopharyngeal fate diversification. Nat. Cell Biol. 2019, 21, 674–686. [Google Scholar] [CrossRef] [PubMed]
- Wang, H.; Holland, P.W.H.; Takahashi, T. Gene profiling of head mesoderm in early zebrafish development: Insights into the evolution of cranial mesoderm. Evodevo 2019, 10, 14. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tolkin, T.; Christiaen, L. Development and evolution of the ascidian cardiogenic mesoderm. Curr. Top. Dev. Biol. 2012, 100, 107–142. [Google Scholar] [CrossRef] [PubMed]
- Paffett-Lugassy, N.; Singh, R.; Nevis, K.R.; Guner-Ataman, B.; O’Loughlin, E.; Jahangiri, L.; Harvey, R.P.; Burns, C.G.; Burns, C.E. Heart field origin of great vessel precursors relies on nkx2.5-mediated vasculogenesis. Nat. Cell Biol. 2013, 15, 1362–1369. [Google Scholar] [CrossRef] [Green Version]
- Choudhry, P.; Trede, N.S.; Oskarsdottir, S.; Vujic, M.; Fasth, A.; Jerome, L.; Papaioannou, V.; Lindsay, E.; Vitelli, F.; Su, H.; et al. DiGeorge syndrome gene tbx1 functions through wnt11r to regulate heart looping and differentiation. PLoS ONE 2013, 8, e58145. [Google Scholar] [CrossRef] [Green Version]
- Van Handel, B.; Montel-Hagen, A.; Sasidharan, R.; Nakano, H.; Ferrari, R.; Boogerd, C.J.; Schredelseker, J.; Wang, Y.; Hunter, S.; Org, T.; et al. Scl represses cardiomyogenesis in prospective hemogenic endothelium and endocardium. Cell 2012, 150, 590–605. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Guner-Ataman, B.; González-Rosa, J.M.; Shah, H.N.; Butty, V.L.; Jeffrey, S.; Abrial, M.; Boyer, L.A.; Burns, C.G.; Burns, C.E. Failed Progenitor Specification Underlies the Cardiopharyngeal Phenotypes in a Zebrafish Model of 22q11.2 Deletion Syndrome. Cell Rep. 2018, 24, 1342–1354. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Razy-Krajka, F.; Gravez, B.; Kaplan, N.; Racioppi, C.; Wang, W.; Christiaen, L. An FGF-driven feed-forward circuit patterns the cardiopharyngeal mesoderm in space and time. eLife 2018, 7. [Google Scholar] [CrossRef] [PubMed]
- Davidson, B.; Shi, W.; Beh, J.; Christiaen, L.; Levine, M. FGF signaling delineates the cardiac progenitor field in the simple chordate, Ciona intestinalis. Genes Dev. 2006, 20, 2728–2738. [Google Scholar] [CrossRef] [Green Version]
- Burger, A.; Lindsay, H.; Felker, A.; Hess, C.; Anders, C.; Chiavacci, E.; Zaugg, J.; Weber, L.M.L.M.; Catena, R.; Jinek, M.; et al. Maximizing mutagenesis with solubilized CRISPR-Cas9 ribonucleoprotein complexes. Development 2016, 143, 2025–2037. [Google Scholar] [CrossRef] [Green Version]
- Gagnon, J.A.; Valen, E.; Thyme, S.B.; Huang, P.; Ahkmetova, L.; Pauli, A.; Montague, T.G.; Zimmerman, S.; Richter, C.; Schier, A.F.; et al. Efficient mutagenesis by Cas9 protein-mediated oligonucleotide insertion and large-scale assessment of single-guide RNAs. PLoS ONE 2014, 9, e98186. [Google Scholar] [CrossRef]
- Jao, L.-E.; Wente, S.R.; Chen, W. Efficient multiplex biallelic zebrafish genome editing using a CRISPR nuclease system. Proc. Natl. Acad. Sci. USA 2013, 110, 13904–13909. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Trubiroha, A.; Gillotay, P.; Giusti, N.; Gacquer, D.; Libert, F.; Lefort, A.; Haerlingen, B.; De Deken, X.; Opitz, R.; Costagliola, S.; et al. A Rapid CRISPR/Cas-based Mutagenesis Assay in Zebrafish for Identification of Genes Involved in Thyroid Morphogenesis and Function. Science 2018, 8, 5647. [Google Scholar] [CrossRef]
- Wu, R.S.; Lam, I.I.; Clay, H.; Duong, D.N.; Deo, R.C.; Coughlin, S.R. A Rapid Method for Directed Gene Knockout for Screening in G0 Zebrafish. Dev. Cell 2018, 46, 112–125. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Metzker, M.L. Sequencing technologies-the next generation. Nat. Rev. Genet. 2010, 11, 31–46. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Blue, G.M.; Winlaw, D. Next Generation Sequencing in Congenital Heart Disease: Gene Discovery and Clinical Application. J. Next Gener. Seq. Appl. 2015, 2. [Google Scholar] [CrossRef] [Green Version]
- Wang, L.; Zhang, Y.; Zhou, T.; Fu, Y.F.; Du, T.T.; Jin, Y.; Chen, Y.; Ren, C.G.; Peng, X.L.; Deng, M.; et al. Functional characterization of Lmo2-Cre transgenic zebrafish. Dev. Dyn. 2008, 237, 2139–2146. [Google Scholar] [CrossRef] [PubMed]
- Chiavacci, E.; Kirchgeorg, L.; Felker, A.; Burger, A.; Mosimann, C. Early frameshift alleles of zebrafish tbx5a that fail to develop the heartstrings phenotype. Matters 2017, 4, e201703000011. [Google Scholar]
- Siu, B.L.; Niimura, H.; Osborne, J.A.; Fatkin, D.; MacRae, C.; Solomon, S.; Benson, D.W.; Seidman, J.G.; Seidman, C.E. Familial dilated cardiomyopathy locus maps to chromosome 2q31. Circulation 1999, 99, 1022–1026. [Google Scholar] [CrossRef]
- Gerull, B.; Gramlich, M.; Atherton, J.; McNabb, M.; Trombitás, K.; Sasse-Klaassen, S.; Seidman, J.G.; Seidman, C.; Granzier, H.; Labeit, S.; et al. Mutations of TTN, encoding the giant muscle filament titin, cause familial dilated cardiomyopathy. Nat. Genet. 2002, 30, 201–204. [Google Scholar] [CrossRef] [PubMed]
- Xu, X.; Meiler, S.E.; Zhong, T.P.; Mohideen, M.; Crossley, D.A.; Burggren, W.W.; Fishman, M.C. Cardiomyopathy in zebrafish due to mutation in an alternatively spliced exon of titin. Nat. Genet. 2002, 30, 205–209. [Google Scholar] [CrossRef]
- Smith, K.A.; Joziasse, I.C.; Chocron, S.; van Dinther, M.; Guryev, V.; Verhoeven, M.C.; Rehmann, H.; van der Smagt, J.J.; Doevendans, P.A.; Cuppen, E.; et al. Dominant-Negative ALK2 Allele Associates with Congenital Heart Defects. Circulation 2009, 119, 3062–3069. [Google Scholar] [CrossRef] [Green Version]
- Walsh, E.C.; Stainier, D.Y. UDP-Glucose Dehydrogenase Required for Cardiac Valve Formation in Zebrafish. Science 2001, 293, 1670–1673. [Google Scholar] [CrossRef]
- Peterson, R.T.; Shaw, S.Y.; Peterson, T.A.; Milan, D.J.; Zhong, T.P.; Schreiber, S.L.; MacRae, C.A.; Fishman, M.C. Chemical suppression of a genetic mutation in a zebrafish model of aortic coarctation. Nat. Biotechnol. 2004, 22, 595–599. [Google Scholar] [CrossRef]
- Basson, C.T.; Bachinsky, D.R.; Lin, R.C.; Levi, T.; Elkins, J.A.; Soults, J.; Grayzel, D.; Kroumpouzou, E.; Traill, T.A.; Leblanc-Straceski, J.; et al. Mutations in human cause limb and cardiac malformation in Holt-Oram syndrome. Nat. Genet. 1997, 15, 30–35. [Google Scholar] [CrossRef]
- Bruneau, B.G.; Logan, M.; Davis, N.; Levi, T.; Tabin, C.J.; Seidman, J.G.; Seidman, C.E. Chamber-specific cardiac expression of Tbx5 and heart defects in Holt- Oram syndrome. Dev. Biol. 1999, 211, 100–108. [Google Scholar] [CrossRef] [Green Version]
- Yi Li, Q.; Newbury-Ecob, R.A.; Terrett, J.A.; Wilson, D.I.; Curtis, A.R.J.; Ho Yi, C.; Gebuhr, T.; Bullen, P.J.; Robson, S.C.; Strachan, T.; et al. Holt-Oram syndrome is caused by mutations in TBX5, a member of the Brachyury (T) gene family. Nat. Genet. 1997, 15, 21–29. [Google Scholar] [CrossRef] [PubMed]
- Papangeli, I.; Scambler, P. The 22q11 deletion: DiGeorge and velocardiofacial syndromes and the role of TBX1. Wiley Interdiscip. Rev. Dev. Biol. 2013, 2, 393–403. [Google Scholar] [CrossRef]
- Yagi, H.; Furutani, Y.; Hamada, H.; Sasaki, T.; Asakawa, S.; Minoshima, S.; Ichida, F.; Joo, K.; Kimura, M.; Imamura, S.; et al. Role of TBX1 in human del22q11.2 syndrome. Lancet 2003, 362, 1366–1373. [Google Scholar] [CrossRef]
- Piotrowski, T.; Schilling, T.F.; Brand, M.; Jiang, Y.J.; Heisenberg, C.P.; Beuchle, D.; Grandel, H.; van Eeden, F.J.; Furutani-Seiki, M.; Granato, M.; et al. Jaw and branchial arch mutants in zebrafish II: Anterior arches and cartilage differentiation. Development 1996, 123, 345–356. [Google Scholar] [PubMed]
- Christiansen, J.; Dyck, J.D.; Elyas, B.G.; Lilley, M.; Bamforth, J.S.; Hicks, M.; Sprysak, K.A.; Tomaszewski, R.; Haase, S.M.; Vicen-Wyhony, L.M.; et al. Chromosome 1q21.1 Contiguous Gene Deletion Is Associated with Congenital Heart Disease. Circ. Res. 2004, 94, 1429–1435. [Google Scholar] [CrossRef] [Green Version]
- Zaidi, S.; Choi, M.; Wakimoto, H.; Ma, L.; Jiang, J.; Overton, J.D.; Romano-Adesman, A.; Bjornson, R.D.; Breitbart, R.E.; Brown, K.K.; et al. De novo mutations in histone-modifying genes in congenital heart disease. Nature 2013, 498, 220–223. [Google Scholar] [CrossRef] [Green Version]
- Greenway, S.C.; Pereira, A.C.; Lin, J.C.; Depalma, S.R.; Samuel, J.; Mesquita, S.M.; Ergul, E.; Conta, J.R.; Korn, J.M.; Steven, A.; et al. De Novo Copy Number Variants Identify New Genes and Loci in Isolated, Sporadic Tetralogy of Fallot. Nat. Genet. 2009, 41, 931–935. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cantù, C.; Felker, A.; Zimmerli, D.; Prummel, K.D.; Cabello, E.M.; Chiavacci, E.; Méndez-Acevedo, K.M.; Kirchgeorg, L.; Burger, S.; Ripoll, J.; et al. Mutations in Bcl9 and Pygo genes cause congenital heart defects by tissue-specific perturbation of Wnt/β-catenin signaling. Genes Dev. 2018, 32, 1443–1458. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tessadori, F.; Roessler, H.I.; Savelberg, S.M.C.; Chocron, S.; Kamel, S.M.; Duran, K.J.; van Haelst, M.M.; van Haaften, G.; Bakkers, J. Effective CRISPR/Cas9-based nucleotide editing in zebrafish to model human genetic cardiovascular disorders. Dis. Model. Mech. 2018, 11, dmm035469. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jahangiri, L.; Sharpe, M.; Novikov, N.; González-Rosa, J.M.; Borikova, A.; Nevis, K.; Paffett-Lugassy, N.; Zhao, L.; Adams, M.; Guner-Ataman, B.; et al. The AP-1 transcription factor component Fosl2 potentiates the rate of myocardial differentiation from the zebrafish second heart field. Development 2016, 143, 113–122. [Google Scholar] [CrossRef] [Green Version]
- Santos-Ledo, A.; Washer, S.; Dhanaseelan, T.; Eley, L.; Alqatani, A.; Chrystal, P.W.; Papoutsi, T.; Henderson, D.J.; Chaudhry, B. Alternative splicing of jnk1a in zebrafish determines first heart field ventricular cardiomyocyte numbers through modulation of hand2 expression. PLOS Genet. 2020, 16, e1008782. [Google Scholar] [CrossRef]
- Aanhaanen, W.T.J.; Moorman, A.F.M.; Christoffels, V.M. Origin and development of the atrioventricular myocardial lineage: Insight into the development of accessory pathways. Birth Defects Res. Part. A Clin. Mol. Teratol. 2011, 91, 565–577. [Google Scholar] [CrossRef]
- Dobrzynski, H.; Boyett, M.R.; Anderson, R.H. New Insights into Pacemaker Activity. Circulation 2007, 115, 1921–1932. [Google Scholar] [CrossRef] [Green Version]
- Ren, J.; Han, P.; Ma, X.; Farah, E.N.; Bloomekatz, J.; Zeng, X.-X.I.; Zhang, R.; Swim, M.M.; Witty, A.D.; Knight, H.G.; et al. Canonical Wnt5b Signaling Directs Outlying Nkx2.5+ Mesoderm into Pacemaker Cardiomyocytes. Dev. Cell 2019, 50, 729–743. [Google Scholar] [CrossRef]
- Gillis, J.A.; Dahn, R.D.; Shubin, N.H. Shared developmental mechanisms pattern the vertebrate gill arch and paired fin skeletons. Proc. Natl. Acad. Sci. USA 2009, 106, 5720–5724. [Google Scholar] [CrossRef] [Green Version]
- Li, Y.; Klena, N.T.; Gabriel, G.C.; Liu, X.; Kim, A.J.; Lemke, K.; Chen, Y.; Chatterjee, B.; Devine, W.; Damerla, R.R.; et al. Global genetic analysis in mice unveils central role for cilia in congenital heart disease. Nature 2015, 521, 520–524. [Google Scholar] [CrossRef] [Green Version]
- MacRae, C.A.; Seidman, C.E. Closing the Genotype-Phenotype Loop for Precision Medicine. Circulation 2017, 136, 1492–1494. [Google Scholar] [CrossRef] [PubMed]



Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kemmler, C.L.; Riemslagh, F.W.; Moran, H.R.; Mosimann, C. From Stripes to a Beating Heart: Early Cardiac Development in Zebrafish. J. Cardiovasc. Dev. Dis. 2021, 8, 17. https://0-doi-org.brum.beds.ac.uk/10.3390/jcdd8020017
Kemmler CL, Riemslagh FW, Moran HR, Mosimann C. From Stripes to a Beating Heart: Early Cardiac Development in Zebrafish. Journal of Cardiovascular Development and Disease. 2021; 8(2):17. https://0-doi-org.brum.beds.ac.uk/10.3390/jcdd8020017
Chicago/Turabian StyleKemmler, Cassie L., Fréderike W. Riemslagh, Hannah R. Moran, and Christian Mosimann. 2021. "From Stripes to a Beating Heart: Early Cardiac Development in Zebrafish" Journal of Cardiovascular Development and Disease 8, no. 2: 17. https://0-doi-org.brum.beds.ac.uk/10.3390/jcdd8020017