
Antiarrhythmic Effects of Vernakalant in Human-Induced Pluripotent Stem Cell-Derived Cardiomyocytes from a Patient with Short QT Syndrome Type 1

 and
and {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Methods
2.1. Ethics Statement
2.2. Clinical Data
2.3. Generation of Human iPS Cells
2.4. Generation of hiPSC-CMs
2.5. Patch-Clamp
2.6. Measurement of Intracellular Calcium Transients
2.7. Drugs
2.8. Statistical Analysis
3. Results
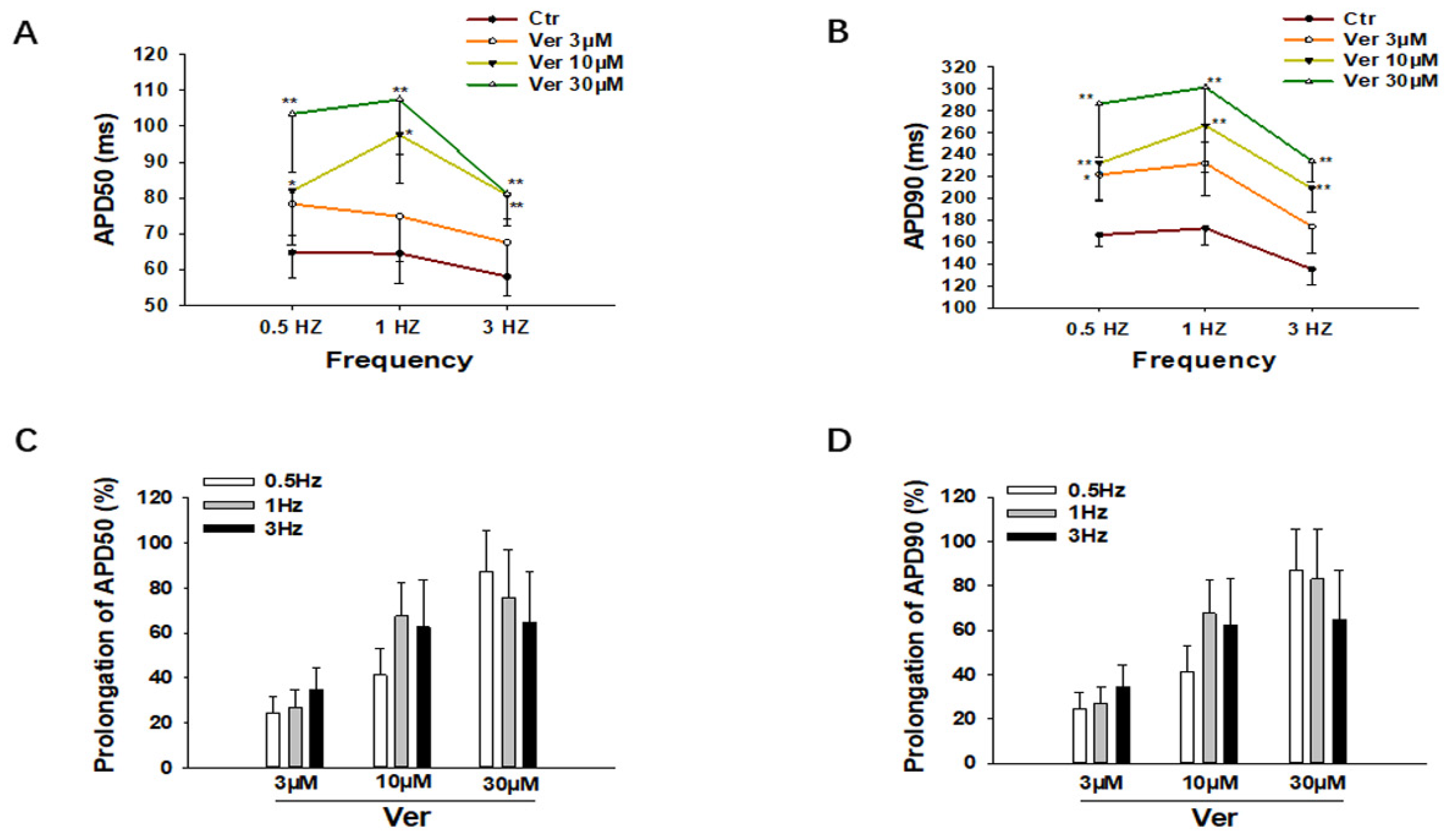
3.1. Effects of Vernakalant on Action Potentials (APs)
3.2. Vernakalant Reduced Arrhythmia-Like Events in SQTS1-hiPSC-CMs
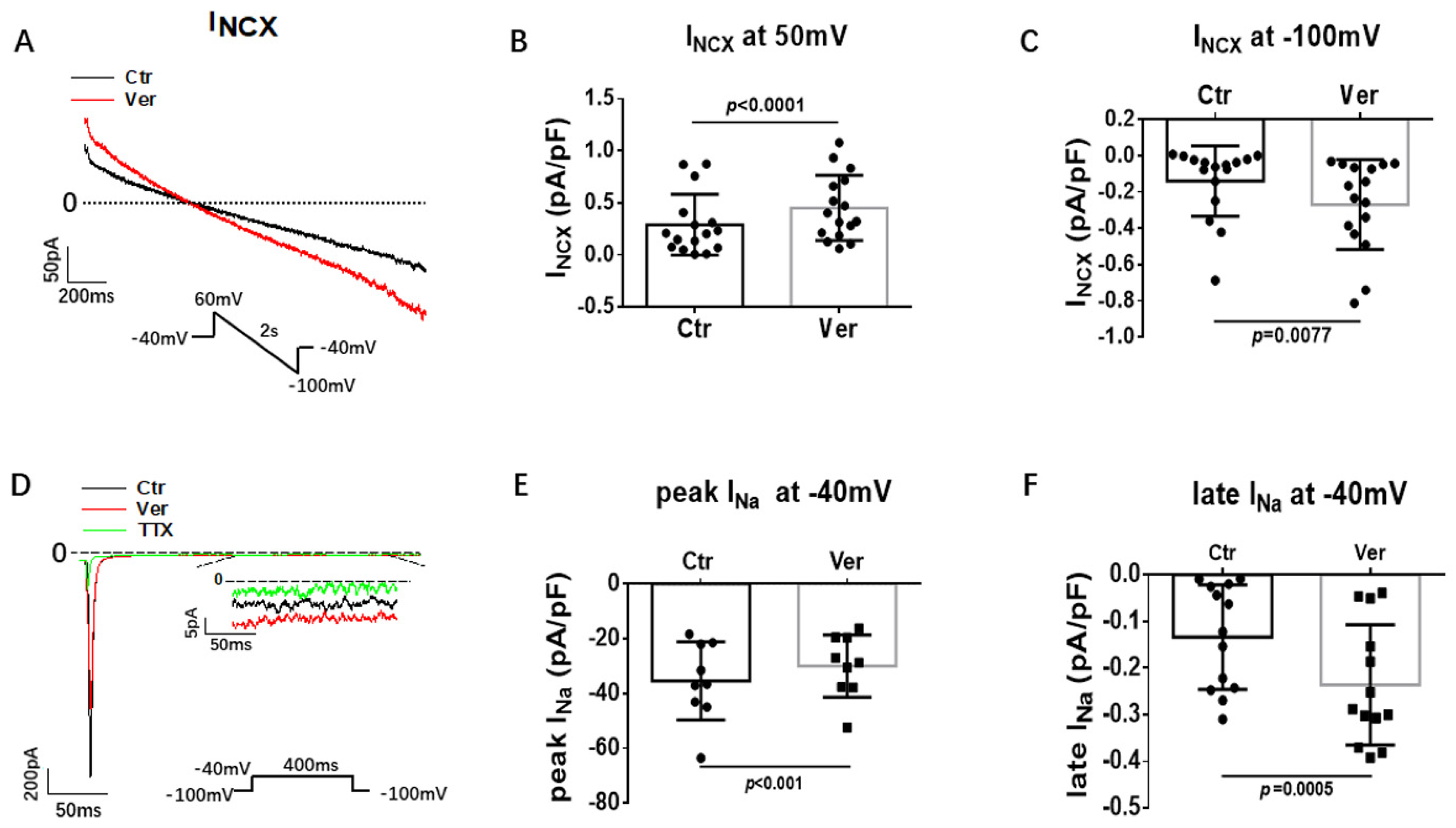
3.3. Effects of Vernakalant on Ion Channel Currents in hiPSC-CMs
4. Discussion
5. Conclusions
6. Study Limitations
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Gussak, I.; Brugada, P.; Brugada, J.; Wright, R.S.; Kopecky, S.L.; Chaitman, B.R.; Bjerregaard, P. Idiopathic short QT interval: A new clinical syndrome? Cardiology 2000, 94, 99–102. [Google Scholar] [CrossRef] [PubMed]
- Brugada, R.; Hong, K.; Dumaine, R.; Cordeiro, J.; Gaita, F.; Borggrefe, M.; Menendez, T.M.; Brugada, J.; Pollevick, G.D.; Wolpert, C.; et al. Sudden death associated with short-QT syndrome linked to mutations in HERG. Circulation 2004, 109, 30–35. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Campuzano, O.; Sarquella-Brugada, G.; Cesar, S.; Arbelo, E.; Brugada, J.; Brugada, R. Recent Advances in Short QT Syndrome. Front. Cardiovasc. Med. 2018, 5, 149. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Thorsen, K.; Dam, V.S.; Kjaer-Sorensen, K.; Pedersen, L.N.; Skeberdis, V.A.; Jurevičius, J.; Treinys, R.; Petersen, I.M.B.S.; Nielsen, M.S.; Oxvig, C.; et al. Loss-of-activity-mutation in the cardiac chloride-bicarbonate exchanger AE3 causes short QT syndrome. Nat. Commun. 2017, 8, 1696. [Google Scholar] [CrossRef] [PubMed]
- Priori, S.G.; Blomstrom-Lundqvist, C.; Mazzanti, A.; Blom, N.; Borggrefe, M.; Camm, J.; Elliott, P.M.; Fitzsimons, D.; Hatala, R.; Hindricks, G.; et al. 2015 ESC Guidelines for the management of patients with ventricular arrhythmias and the prevention of sudden cardiac death: The Task Force for the Management of Patients with Ventricular Arrhythmias and the Prevention of Sudden Cardiac Death of the European Society of Cardiology (ESC). Endorsed by: Association for European Paediatric and Congenital Cardiology (AEPC). Eur. Heart J. 2015, 36, 2793–2867. [Google Scholar]
- Giustetto, C.; Di Monte, F.; Wolpert, C.; Borggrefe, M.; Schimpf, R.; Sbragia, P.; Leone, G.; Maury, P.; Anttonen, O.; Haissaguerre, M.; et al. Short QT syndrome: Clinical findings and diagnostic-therapeutic implications. Eur. Heart J. 2006, 27, 2440–2447. [Google Scholar] [CrossRef] [Green Version]
- Mazzanti, A.; Kanthan, A.; Monteforte, N.; Memmi, M.; Bloise, R.; Novelli, V.; Miceli, C.; O’Rourke, S.; Borio, G.; Zienciuk-Krajka, A.; et al. Novel insight into the natural history of short QT syndrome. J. Am. Coll. Cardiol. 2014, 63, 1300–1308. [Google Scholar] [CrossRef] [Green Version]
- Abriel, H.; Rougier, J.S. beta-blockers in congenital short-QT syndrome as ion channel blockers. J. Cardiovasc. Electrophysiol. 2013, 24, 1172–1174. [Google Scholar] [CrossRef]
- Mizobuchi, M.; Enjoji, Y.; Yamamoto, R.; Ono, T.; Funatsu, A.; Kambayashi, D.; Kobayashi, T.; Nakamura, S. Nifekalant and disopyramide in a patient with short QT syndrome: Evaluation of pharmacological effects and electrophysiological properties. Pacing Clin. Electrophysiol. 2008, 31, 1229–1232. [Google Scholar] [CrossRef]
- Giustetto, C.; Schimpf, R.; Mazzanti, A.; Scrocco, C.; Maury, P.; Anttonen, O.; Probst, V.; Blanc, J.-J.; Sbragia, P.; Dalmasso, P.; et al. Long-term follow-up of patients with short QT syndrome. J. Am. Coll. Cardiol. 2011, 58, 587–595. [Google Scholar] [CrossRef] [Green Version]
- Gaita, F.; Giustetto, C.; Bianchi, F.; Schimpf, R.; Haissaguerre, M.; Calò, L.; Brugada, R.; Antzelevitch, C.; Borggrefe, M.; Wolpert, C. Short QT syndrome: Pharmacological treatment. J. Am. Coll. Cardiol. 2004, 43, 1494–1499. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- McPate, M.J.; Duncan, R.S.; Hancox, J.C.; Witchel, H.J. Pharmacology of the short QT syndrome N588K-hERG K+ channel mutation: Differential impact on selected class I and class III antiarrhythmic drugs. Br. J. Pharmacol. 2008, 155, 957–966. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wolpert, C.; Schimpf, R.; Giustetto, C.; Antzelevitch, C.; Cordeiro, J.; Dumaine, R.; Brugada, R.; Hong, K.; Bauersfeld, U.; Gaita, F.; et al. Further insights into the effect of quinidine in short QT syndrome caused by a mutation in HERG. J. Cardiovasc. Electrophysiol. 2005, 16, 54–58. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- El-Battrawy, I.; Lan, H.; Cyganek, L.; Zhao, Z.; Li, X.; Buljubasic, F.; Lang, S.; Yücel, G.; Sattler, K.; Zimmermann, W.; et al. Modeling Short QT Syndrome Using Human-Induced Pluripotent Stem Cell-Derived Cardiomyocytes. J. Am. Heart Assoc. 2018, 7, e007394. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Frommeyer, G.; Ellermann, C.; Dechering, D.G.; Kochhäuser, S.; Bögeholz, N.; Güner, F.; Leitz, P.; Pott, C.; Eckardt, L. Ranolazine and Vernakalant Prevent Ventricular Arrhythmias in an Experimental Whole-Heart Model of Short QT Syndrome. J. Cardiovasc. Electrophysiol. 2016, 27, 1214–1219. [Google Scholar] [CrossRef]
- Association World Medical. World Medical Association Declaration of Helsinki: Ethical principles for medical research involving human subjects. JAMA 2013, 310, 2191–2194. [Google Scholar] [CrossRef] [Green Version]
- Cyganek, L.; Tiburcy, M.; Sekeres, K.; Gerstenberg, K.; Bohnenberger, H.; Lenz, C.; Henze, S.; Stauske, M.; Salinas, G.; Zimmermann, W.-H.; et al. Deep phenotyping of human induced pluripotent stem cell-derived atrial and ventricular cardiomyocytes. JCI Insight 2018, 3, e99941. [Google Scholar] [CrossRef] [Green Version]
- Chen, S.; Liu, S.; Purerfellner, H. Vernakalant as a Novel Anti-Arrhythmic Agent for Converting of Atrial Fibrillation, Molecular Mechanism, Updated Clinical Efficacy, and Future Development. Curr. Pharm. Des. 2015, 21, 4400–4408. [Google Scholar] [CrossRef]
- Dorian, P.; Pinter, A.; Mangat, I.; Korley, V.; Cvitkovic, S.S.; Beatch, G.N. The effect of vernakalant (RSD1235), an investigational antiarrhythmic agent, on atrial electrophysiology in humans. J. Cardiovasc. Pharmacol. 2007, 50, 35–40. [Google Scholar] [CrossRef]
- Fedida, D.; Orth, P.M.; Chen, J.Y.; Lin, S.; Plouvier, B.; Jung, G.; Ezrin, A.M.; Beatch, G.N. The mechanism of atrial antiarrhythmic action of RSD1235. J. Cardiovasc. Electrophysiol. 2005, 16, 1227–1238. [Google Scholar] [CrossRef]
- Vizzardi, E.; Salghetti, F.; Bonadei, I.; Gelsomino, S.; Lorusso, R.; D’Aloia, A.; Curnis, A. A new antiarrhythmic drug in the treatment of recent-onset atrial fibrillation: Vernakalant. Cardiovasc. Ther. 2013, 31, e55–e62. [Google Scholar] [CrossRef] [PubMed]
- Kossaify, A. Vernakalant in Atrial Fibrillation: A Relatively New Weapon in the Armamentarium against an Old Enemy. Drug Target Insights 2019, 13, 1177392819861114. [Google Scholar] [CrossRef] [PubMed]
- Giles, W.R.; Imaizumi, Y. Comparison of potassium currents in rabbit atrial and ventricular cells. J. Physiol. 1988, 405, 123–145. [Google Scholar] [CrossRef] [PubMed]
- Ng, S.Y.; Wong, C.K.; Tsang, S.Y. Differential gene expressions in atrial and ventricular myocytes: Insights into the road of applying embryonic stem cell-derived cardiomyocytes for future therapies. Am. J. Physiol. Cell Physiol. 2010, 299, C1234–C1249. [Google Scholar] [CrossRef] [Green Version]
- Ravens, U.; Poulet, C.; Wettwer, E.; Knaut, M. Atrial selectivity of antiarrhythmic drugs. J. Physiol. 2013, 591, 4087–4097. [Google Scholar] [CrossRef] [Green Version]
- Hong, K.; Bjerregaard, P.; Gussak, I.; Brugada, R. Short QT syndrome and atrial fibrillation caused by mutation in KCNH2. J. Cardiovasc. Electrophysiol. 2005, 16, 394–396. [Google Scholar] [CrossRef]
- Zienciuk-Krajka, A.; Kukla, P.; Stec, S.; Raczak, G. Short QT syndrome presenting with atrial fibrillation and LV hypertrophy. Int. J. Cardiol. 2012, 156, e9–e10. [Google Scholar] [CrossRef]
- Shinnawi, R.; Shaheen, N.; Huber, I.; Shiti, A.; Arbel, G.; Gepstein, A.; Ballan, N.; Setter, N.; Tijsen, A.J.; Borggrefe, M.; et al. Modeling Reentry in the Short QT Syndrome With Human-Induced Pluripotent Stem Cell-Derived Cardiac Cell Sheets. J. Am. Coll. Cardiol. 2019, 73, 2310–2324. [Google Scholar] [CrossRef]
- Wettwer, E.; Christ, T.; Endig, S.; Rozmaritsa, N.; Matschke, K.; Lynch, J.J.; Pourrier, M.; Gibson, J.K.; Fedida, D.; Knaut, M.; et al. The new antiarrhythmic drug vernakalant: Ex vivo study of human atrial tissue from sinus rhythm and chronic atrial fibrillation. Cardiovasc. Res. 2013, 98, 145–154. [Google Scholar] [CrossRef] [Green Version]
- Skibsbye, L.; Poulet, C.; Diness, J.G.; Bentzen, B.H.; Yuan, L.; Kappert, U.; Matschke, K.; Wettwer, E.; Ravens, U.; Grunnet, M.; et al. Small-conductance calcium-activated potassium (SK) channels contribute to action potential repolarization in human atria. Cardiovasc. Res. 2014, 103, 156–167. [Google Scholar] [CrossRef] [Green Version]
- Zhang, X.D.; Timofeyev, V.; Li, N.; Myers, R.E.; Zhang, D.-M.; Singapuri, A.; Lau, V.C.; Bond, C.T.; Adelman, J.; Lieu, D.K.; et al. Critical roles of a small conductance Ca2+-activated K+ channel (SK3) in the repolarization process of atrial myocytes. Cardiovasc. Res. 2014, 101, 317–325. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sutanto, H.; Laudy, L.; Clerx, M.; Dobrev, D.; Crijns, H.J.; Heijman, J. Maastricht antiarrhythmic drug evaluator (MANTA): A computational tool for better understanding of antiarrhythmic drugs. Pharmacol. Res. 2019, 148, 104444. [Google Scholar] [CrossRef] [PubMed]
- Liang, P.; Sallam, K.; Wu, H.; Li, Y.; Itzhaki, I.; Garg, P.; Zhang, Y.; Termglichan, V.; Lan, F.; Gu, M.; et al. Patient-Specific and Genome-Edited Induced Pluripotent Stem Cell-Derived Cardiomyocytes Elucidate Single-Cell Phenotype of Brugada Syndrome. J. Am. Coll. Cardiol. 2016, 68, 2086–2096. [Google Scholar] [CrossRef] [PubMed]
- Blaustein, M.P.; Lederer, W.J. Sodium/calcium exchange: Its physiological implications. Physiol. Rev. 1999, 79, 763–854. [Google Scholar] [CrossRef]
- Tykocki, N.R.; Jackson, W.F.; Watts, S.W. Reverse-mode Na+/Ca2+ exchange is an important mediator of venous contraction. Pharmacol. Res. 2012, 66, 544–554. [Google Scholar] [CrossRef] [Green Version]







Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Xu, Q.; Huang, X.; Meng, Z.; Li, Y.; Zhong, R.; Li, X.; Cyganek, L.; El-Battrawy, I.; Akin, I.; Zhou, X.; et al. Antiarrhythmic Effects of Vernakalant in Human-Induced Pluripotent Stem Cell-Derived Cardiomyocytes from a Patient with Short QT Syndrome Type 1. J. Cardiovasc. Dev. Dis. 2022, 9, 112. https://0-doi-org.brum.beds.ac.uk/10.3390/jcdd9040112
Xu Q, Huang X, Meng Z, Li Y, Zhong R, Li X, Cyganek L, El-Battrawy I, Akin I, Zhou X, et al. Antiarrhythmic Effects of Vernakalant in Human-Induced Pluripotent Stem Cell-Derived Cardiomyocytes from a Patient with Short QT Syndrome Type 1. Journal of Cardiovascular Development and Disease. 2022; 9(4):112. https://0-doi-org.brum.beds.ac.uk/10.3390/jcdd9040112
Chicago/Turabian StyleXu, Qiang, Xuemei Huang, Zenghui Meng, Yingrui Li, Rujia Zhong, Xin Li, Lukas Cyganek, Ibrahim El-Battrawy, Ibrahim Akin, Xiaobo Zhou, and et al. 2022. "Antiarrhythmic Effects of Vernakalant in Human-Induced Pluripotent Stem Cell-Derived Cardiomyocytes from a Patient with Short QT Syndrome Type 1" Journal of Cardiovascular Development and Disease 9, no. 4: 112. https://0-doi-org.brum.beds.ac.uk/10.3390/jcdd9040112