Microglial Response to Aspergillus flavus and Candida albicans: Implications in Endophthalmitis


1
Jhaveri Microbiology Centre, Brien Holden Eye Research Centre, L. V. Prasad Eye Institute, Hyderabad, Telangana 500034, India
2
Manipal Academy of Higher Education, Manipal, Karnataka 576104, India
3
Kallam Anji Reddy Molecular Genetics Laboratory, Jhaveri Microbiology Centre, Brien Holden Eye Research Centre, L. V. Prasad Eye Institute, Hyderabad, Telangana 500034, India
4
Department of Ophthalmology, Visual and Anatomical Sciences, Wayne State University, Detroit, MI 48202, USA
*
Author to whom correspondence should be addressed.
J. Fungi 2020, 6(3), 162; https://0-doi-org.brum.beds.ac.uk/10.3390/jof6030162
Submission received: 30 July 2020
/
Revised: 3 September 2020
/
Accepted: 4 September 2020
/
Published: 5 September 2020
Abstract
:Aspergillus flavus is the most common etiology of fungal endophthalmitis in India, while Candida albicans is the causative agent in the West. In this study, we determined the role of microglial cells in evoking an inflammatory response following an infection with A. flavus and C. albicans strains isolated from patients with endophthalmitis. Microglia (CHME-3) cells were infected with A. flavus and C. albicans and the expression of Toll-Like Receptors (TLRs), cytokines and Matrix metalloproteinases (MMPs) were assessed at various time intervals. A. flavus infected cells induced higher expressions of TLR-1, -2, -5, -6, -7 and -9 and cytokines such as IL-1α, IL-6, IL-8, IL-10 and IL-17. In contrast, C. albicans infected microglia induced only TLR-2 along with the downregulation of IL-10 and IL-17. The expression of MMP-9 (Matrix metalloproteinase-9) was however upregulated in both A. flavus and C. albicans infected microglia. These results indicate that microglial cells have the ability to incite an innate response towards endophthalmitis causing fungal pathogens via TLRs and inflammatory mediators. Moreover, our study highlights the differential responses of microglia towards yeast vs. filamentous fungi.
1. Introduction
Fungal endophthalmitis is a rare clinical entity globally, however, in tropical countries such as India and China, this incidence varies from 17–30% of the total cases seen and represents a significant problem, associated with a delayed diagnosis and poor prognosis [1]. The spectrum of fungal agents causing endophthalmitis depends on the clinical setting and the geographical location. While Candida sp. (yeast) is the most frequent pathogen reported from the Western countries such as the USA and Europe, in tropical countries such as India, the etiology of fungal pathogens includes Aspergillus sp. mainly along with Alternaria, Fusarium, and Curvularia species [1,2,3,4]. Fungal endophthalmitis due to yeast (Candida albicans) are usually endogenous and differ from filamentous fungi (Aspergillus or Fusarium sp.) as treatment is often successful, whereas mold endophthalmitis is usually exogenous and often results in permanent loss of vision. The major mechanism responsible for increased virulence in clinical Candida isolates in addition to viral coinfections is also the overexpression of plasma membrane efflux pumps and the use of P-glycoprotein pump inhibitor associated drugs for the enhancement of antifungal activity is now being proposed [5]. A previous study in our laboratory showed a high rate of detection of fungal pathogens in nearly 50% of our culture-negative samples by a Next Generation Sequencing (NGS) analysis and was identified to be predominantly Aspergillus sp. [3], which suggests that fungal endophthalmitis might be more prevalent than previously believed, especially in the developing countries. In series from India [4], the outcome was unfavorable in more than 50% of the patients with fungal endophthalmitis despite providing intensive therapy and the administration of intravitreal steroids to promote a faster clearance of inflammation remains controversial [6]. The pathogenesis of fungal endophthalmitis involves complex host pathogen interactions resulting in severe intraocular inflammation, and retinal tissue damage. The immune response is critical to the survival of all pathogenic organisms, and along with phagocyting the pathogen, it also facilitates the repair and regeneration of inflamed tissue [7]. Possible virulence factors in fungal endophthalmitis depend largely on the virulence of the organism. Although there are similar characteristics between various fungi and their immune response, each fungus has varying virulence mechanisms resulting in differential immune responses [8]. Therefore, the personalized manipulation of the inflammatory response could offer strategies to control or prevent exacerbations in those diseases.
Microglia, which represents the resident mononuclear phagocyte population present in the central nervous system (CNS), are among the potential retinal glial cells involved in antimicrobial defense [9] that respond to local injury and infectious agents. Under pathological conditions, microglia along with macrophages release inflammatory mediators that control the neuroinflammatory response via an enhanced recruitment of peripheral tissue dendritic cells, neutrophils and lymphocytes [10]. The role of microglia along with Muller glial cells in the initiation of the innate response to live pathogens during endophthalmitis has already been reported earlier, [9,11], however, despite the increasing prevalence of fungal identification in the intraocular fluids from endophthalmitis patients, the effect of fungal infection on the inflammatory response in microglia is poorly defined. Understanding the available information on how microglial cells react and influence disease progression in fungal infections is important and warrants further investigation to mitigate the number of clinical cases of patients with fungal endophthalmitis not responsive to standard therapy [12]. Additionally, a previous 25 year study at our institute found the filamentous fungus Aspergillus flavus (A. flavus) as one of the most clinically relevant fungal pathogens [13] which is a less studied organism compared to its related and well known human pathogen, A. fumigatus [14,15,16]. The purpose of the present study was to study the hypothesis that microglia cells would have a differential inflammatory response to infection by C. albicans and A. flavus isolates.
2. Materials and Methods
2.1. Ethics Statement
All procedures were approved by the Institutional Review Board of the L V Prasad Eye Institute, dated 11th September 2018 (LEC 09-18-124).
2.2. Fungal Isolates
The fungal isolates used in the study were clinical strains of C. albicans (L-614/2017) and A. flavus (L-416/2018) isolated from patients diagnosed clinically with postoperative fungal endophthalmitis and were operated and treated at our institute, which is a tertiary referral center in South India. The demographic details of these patients are given in Table 1. Following a detailed biomicroscopic examination including a retinal examination, these underwent pars plana vitrectomy and the vitreous sample was collected and transported to the microbiology laboratory immediately, where it was cultured on appropriate bacterial and fungal media. All media were incubated at 37 °C while the fungal media were incubated at 25 °C for 2 weeks. While Candida albicans was identified by ViTEK 2, Aspergillus flavus was identified based on their colony characteristics and microscopic features of its conidia. The A. flavus isolate was stocked and preserved in water at room temperature, while the C. albicans isolate was preserved on Sabouraud dextrose agar slopes at room temperature.
2.3. Generation of Fungal Preparations
C. albicans was cultured in Sabouraud Dextrose broth at 150 rpm under 37 °C for 24 h. The cells were then counted in a haemocytometer after isolating them by centrifugation at 4 °C for 3000× g for 15 min. Similarly, A. flavus was cultured on SDA and conidia were harvested after 5 days at 37 °C by scraping into phosphate-buffered saline (PBS, Sigma, St. Louis, MO, USA). The mix was then filtered using a muslin cloth to avoid hyphal contamination and washed in PBS at 8000 rpm for 10 min. The pellet was then resuspended in PBS for enumeration as described earlier.
2.4. In Vitro Culture of Microglia Cells (CHME-3) and Fungal Cocultivation
Human microglial cells (CHME-3) [17] were maintained and serially passaged in a cellular grade culture flask (Nunc EasYFlask 75 cm2) in DMEM (Gibco™, Waltham, MA, USA) with 10% fetal bovine serum (FBS, Gibco™) and 1% antibiotic cocktail and maintained at 37 °C and 5% carbon dioxide in a humidified incubator. CHME-3 cells were grown to a confluence of 1.2 × 106 cells mL−1 in 6-well plates in antibiotics and serum-free DMEM for 16 to 18 h prior to fungal stimulation. The cells were then cocultured with Candida albicans or Aspergillus flavus strains with an MOI of 5:1 at 37 °C in 5% CO2. The uninfected CHME-3 cells were cultured under the same medium conditions. Upon the completion of incubation, at various time points (6–24 h), the culture supernatant were assessed for growth and viability as described previously [18]. The culture supernatants were filtered using a 0.33 μm membrane filter to remove conidia and debris. The samples (both supernatants and cells) were stored at −80 °C prior to RNA extraction and the Enzyme-Linked Immunosorbent Assay (ELISA).
2.5. Total RNA Extraction and Quantitative Real-Time Polymerase Chain Reaction
Total RNA was extracted from the CHME-3 cells using the Qiagen RNeasy Mini kits and cDNA was synthesized using 1 µg of total RNA by Maxima first-strand cDNA synthesis kit (SuperScript; Verso). The cDNA was then amplified using gene-specific PCR primers for human Toll-Like Receptors (TLRs) (TLR1–TLR7 and TLR9), cytokines/chemokine (IL-1α, IL-1β, IL-6, IL-8, TNF-α, IL-10, IL-17, M-CSF), Matrix Metalloproteinases (MMPs) (MMP-2, MMP-9) and TIMP-1 (Tissue Inhibitor of Metalloproteinases) using SYBR Green (DyNAmo Flash SYBR Green qPCR Kit, Thermo Scientific, Waltham, MA, USA) on the Real-Time PCR system (Applied Biosystems Qauntstudio3 system). The real-time primers are listed in Supplementary Table S1. The quantification of gene expression was determined via the comparative ΔΔCT method. Gene expression in the test samples was normalized to the endogenous control β-actin, and was reported as the fold change relative to uninfected controls.
2.6. Enzyme-Linked Immunosorbent Assay (ELISA)
Interleukin-6 (sensitivity < 0.11 pg/mL), IL-8 (sensitivity < 0.13 pg/mL), IL-10 (sensitivity < 0.56 pg/mL), and GM-CSF (sensitivity < 0.35 pg/mL) concentrations in cell culture supernatant were determined using the MAGPIX Milliplex kit (Merck) following the manufacturer’s protocol. Untreated cells were taken as a negative control. Additionally, the MMP-9 concentration in culture supernatants was also measured using a sandwich ELISA (Sigma Aldrich, Merk) and read using a BT 2000 Microkinetics Reader at 450 nm. A standard curve was constructed to quantify the MMP-9 with a detection range of 8–6000 pg/mL.
2.7. Immunofluorescence Assay
For immunocytochemistry staining, microglia cells were grown on coverslips and infected with A. flavus and C. albicans for the indicated time period. The cells were washed three times with phosphate-buffered saline (PBS) and fixed with 4% paraformaldehyde and blocked in 2.5% BSA for 1 h at room temperature followed by an incubation with IL-17, and IL-1β (Abcam, Cambridge, UK) (1:200 dilution) antibodies overnight at 4 °C. Following the removal of the primary antibodies, the cells were then extensively washed with PBS and incubated with Invitrogen Alexa Fluor 594 goat anti-rabbit IgG secondary antibody prepared in 1% BSA in PBS at a dilution of 1:400 for 1 h at room temperature and mounted using Fluoroshield™ medium and visualized under a fluorescence microscope (Carl Ziess AXIO 503 monoscope A1). The level of fluorescence intensity were quantified using Image J version 1.44 software (National Institutes of Health, USA). The corrected total cell fluorescence was calculated from the regions of interest (ROIs) selected and the mean fluorescence values were plotted after subtracting the background value [19].
2.8. Heatmap and Network Analysis for Differentially Expressed Genes (DEGs)
A gene expression heatmap was constructed employing Differentially Expressed Genes (DEGs) in human microglial cells challenged with A. flavus and C. albicans at the desired time point. DEGs with a >2-fold change in at least three data sets were selected and a clustered heatmap was plotted using the online tool heatmapper.ca [20]. The heatmap colors displayed a normalized expression value (zero meant normalization of log2 transformed expression). Genes with significant differential expression were used to generated protein–protein interaction network (PPI) using STRING (Search Tool for the Retrieval of Interacting Genes/Proteins 9.1) [21] (http://string-db.org/) database. The results of the analyses generated a gene–protein interaction network where the intensity of the edges reflected the strength of the interaction score.
2.9. Statistical Analysis
All the values were presented as the mean ± SE. The statistical analysis was performed using Student’s t-test (GraphPad Prism software version 5.0) for a comparison of two groups. The following symbols were used to indicate statistical significance: * p ≤ 0.05 and ** p ≤ 0.005. All experiments were performed at least three times unless indicated otherwise.
3. Results
3.1. Toll-Like Receptors’ (TLRs) Expression Profile in Response to Aspergillus flavus and Candida albicans in Microglia
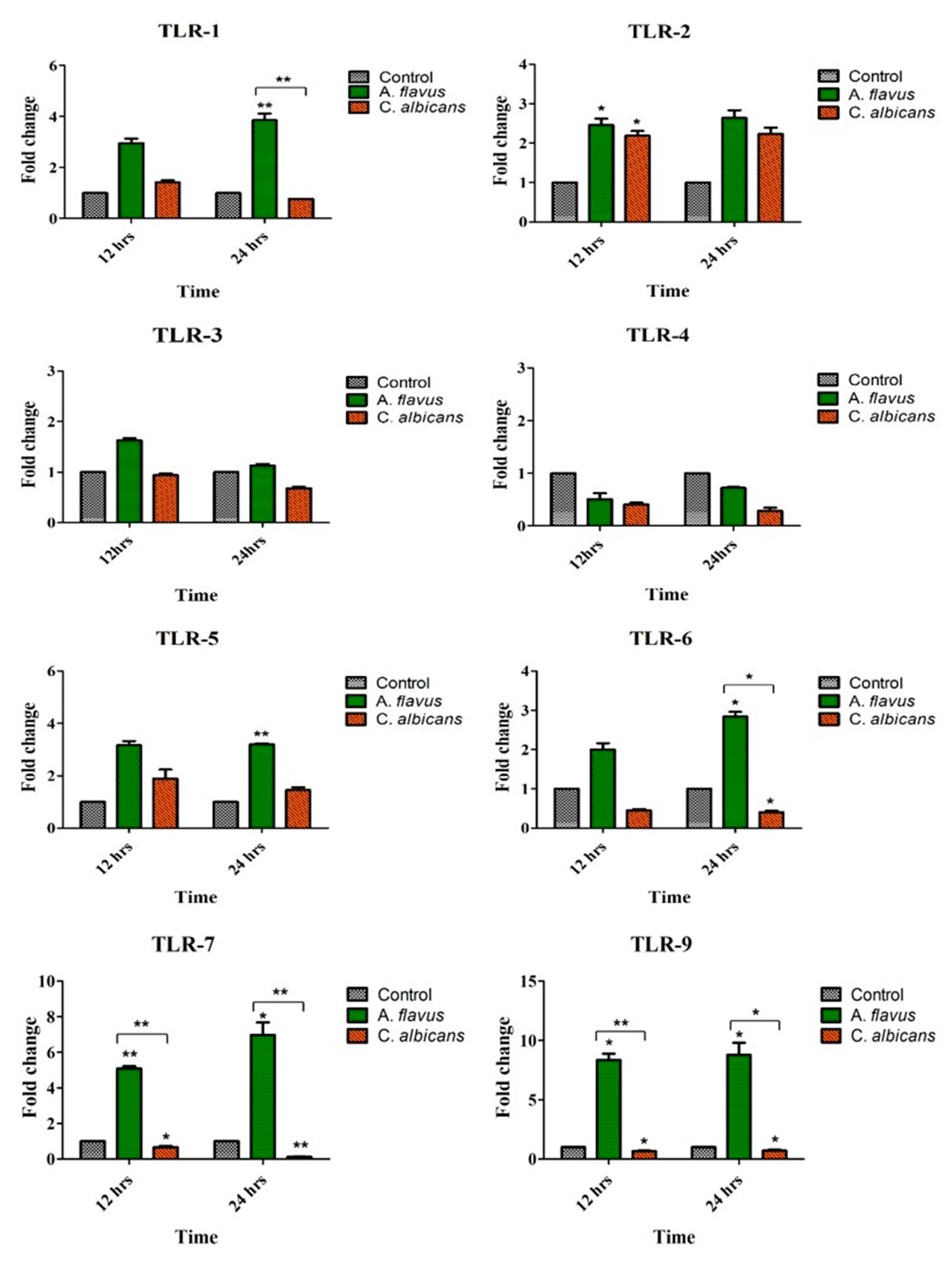
A. flavus challenged microglial cells demonstrated an increased expression of various TLRs compared to uninfected cells, which included TLR-1 (3.8-fold; p < 0.005), TLR-5 (3-fold; p < 0.005) and TLR-6 (2.8-fold; p < 0.05) at 24 h postinfection (p.i.). Additionally, the mRNA expression of TLR-7 (5-fold; p < 0.005) and TLR-9 (8-fold; p < 0.05) were overexpressed at 12 h itself, while TLR-2 was slightly upregulated (2.4-fold) by microglia cells infected with A. flavus at the same time point (Figure 1).
In comparison, C. albicans challenged microglial cells demonstrated an increased expression of only TLR-2 (2.2-fold) at 24 h in comparison to uninfected microglia cells (Figure 1). TLR-3 and TLR-4, however, did not show any expression in cells challenged with both the fungal pathogens. To understand if human microglial cells have a differential response to conidia (A. flavus) and yeast-like fungus (C. albicans), we found that TLR-1, TLR-6, TLR-7 and TLR-9 were highly expressed in cells infected with A. flavus but downregulated in microglial cells challenged with C. albicans (Figure 1).
3.2. Induction of Immune Mediators by Microglia Challenged with A. flavus and C. albicans
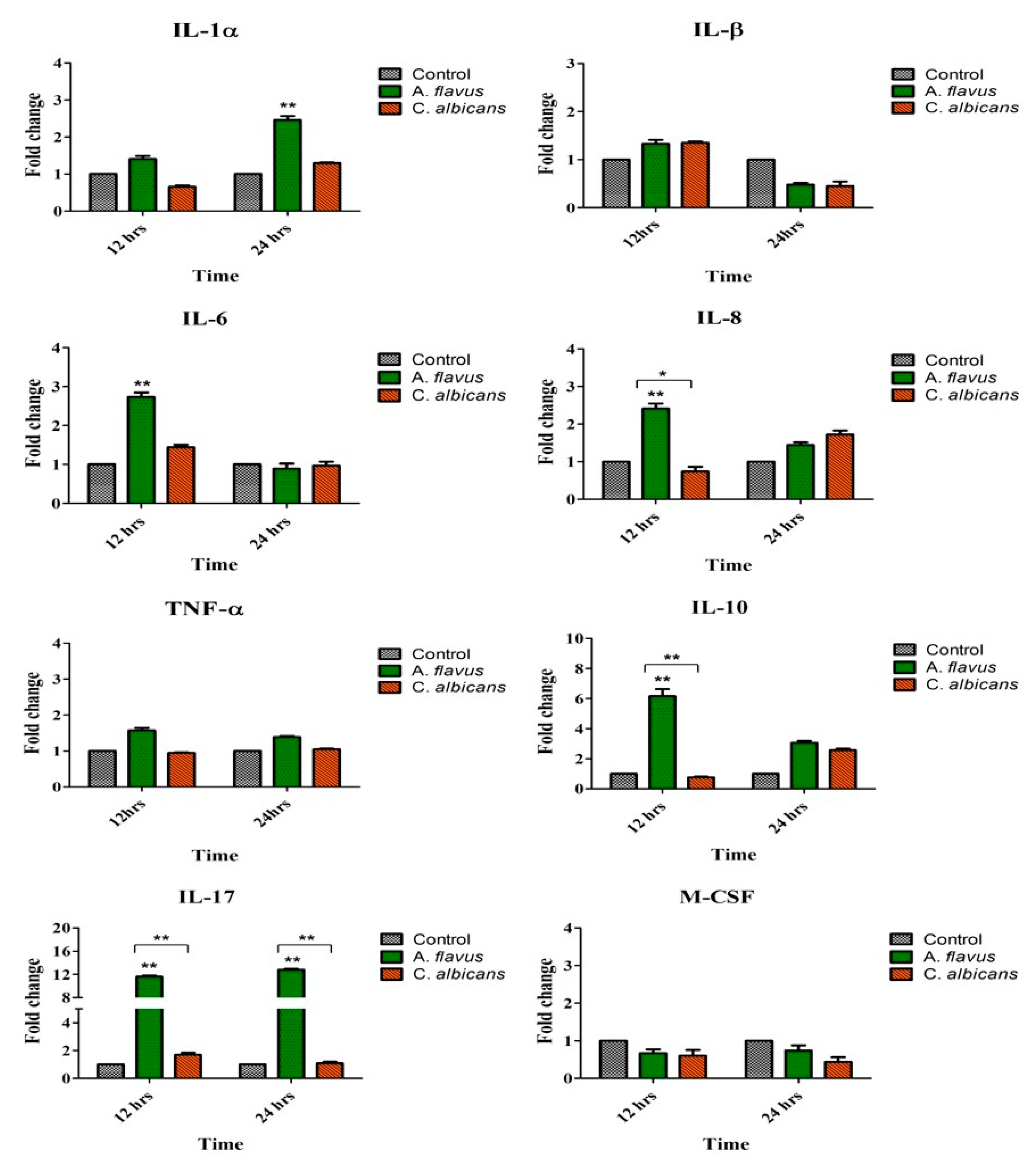
Similar to TLRs’ activation of microglial cells by A. flavus, there was an increased expression of proinflammatory mediators such as IL-6 (2.7-fold) and IL-8 (2.4-fold) at 12 h postinfection while IL-1α (2.4-fold) was expressed at 24 h p.i. compared to uninfected cells. Similarly anti-inflammatory cytokine IL-10 (6-fold) was also upregulated at 12 h p.i. (p < 0.005) along with IL-17 (11.6-fold; p < 0.005) which showed continued increased expression up to 24 h p.i. (12.7-fold, p < 0.005) (Figure 2). There was however, no significant expression of IL1β, TNF-α and M-CSF in microglia cells challenged with both A. flavus as well as C. albicans. Interestingly none of the immune mediators tested were expressed in cells infected with C. albicans. Comparing the expression between the two fungal pathogens, we found that microglial cells infected with A. flavus were significantly upregulated in IL-8 (p < 0.05), IL-10 (p < 0.005) and IL-17 (p < 0.005) at 12 h p.i. whereas in C. albicans infected cells these mediators were downregulated at the indicated time points (Figure 2).
3.3. Expression of Tissue Remodeling Enzymes by A. flavus and C. albicans Infected Microglia
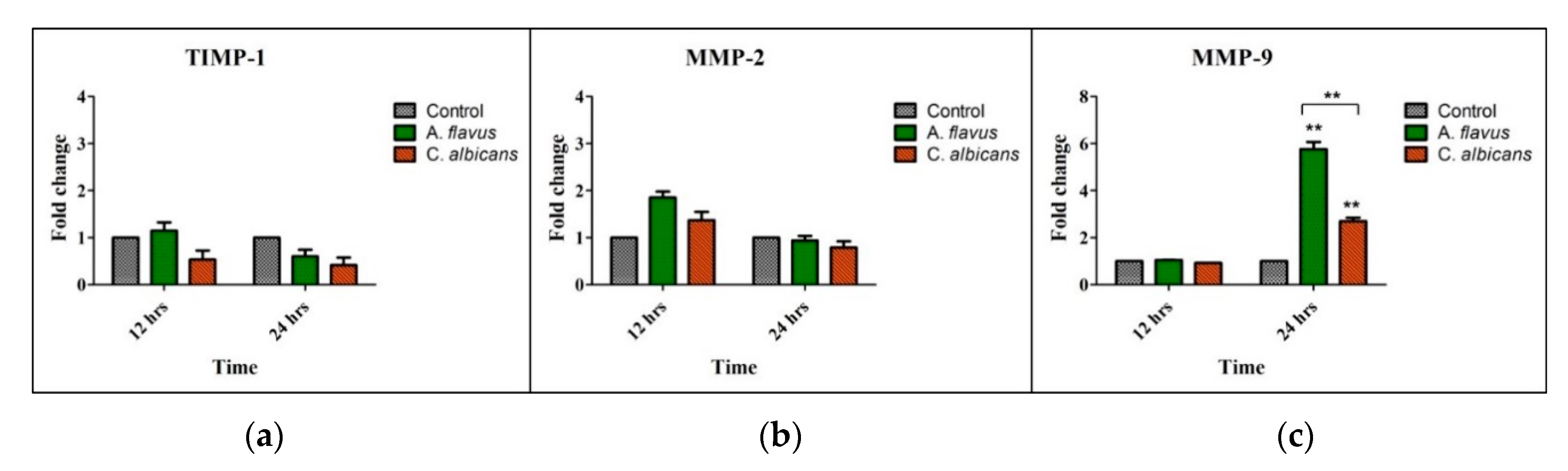
Matrix metalloproteinases’ (MMPs), such as MMP-2, MMP-9 and Tissue Inhibitor of metalloproteinases (TIMP-1), mRNA levels were assessed in CHME-3 cells challenged with A. flavus and C. albicans compared to uninfected cells and we observed an increased mRNA expression of only MMP-9 at 24 h p.i. (5.8-fold and 2.7-fold, respectively) (p < 0.005) (Figure 3c). However, the expression of MMP-9 in A. flavus infected cells was significantly higher (p < 0.005) than cells infected with C. albicans (Figure 3c). In comparison, MMP-2 and its inhibitor TIMP-1 had no significant expression in cells infected with both A. flavus and C. albicans (Figure 3a,b).
3.4. Protein Expression of Immune Mediators in Microglia Infected with Fungus
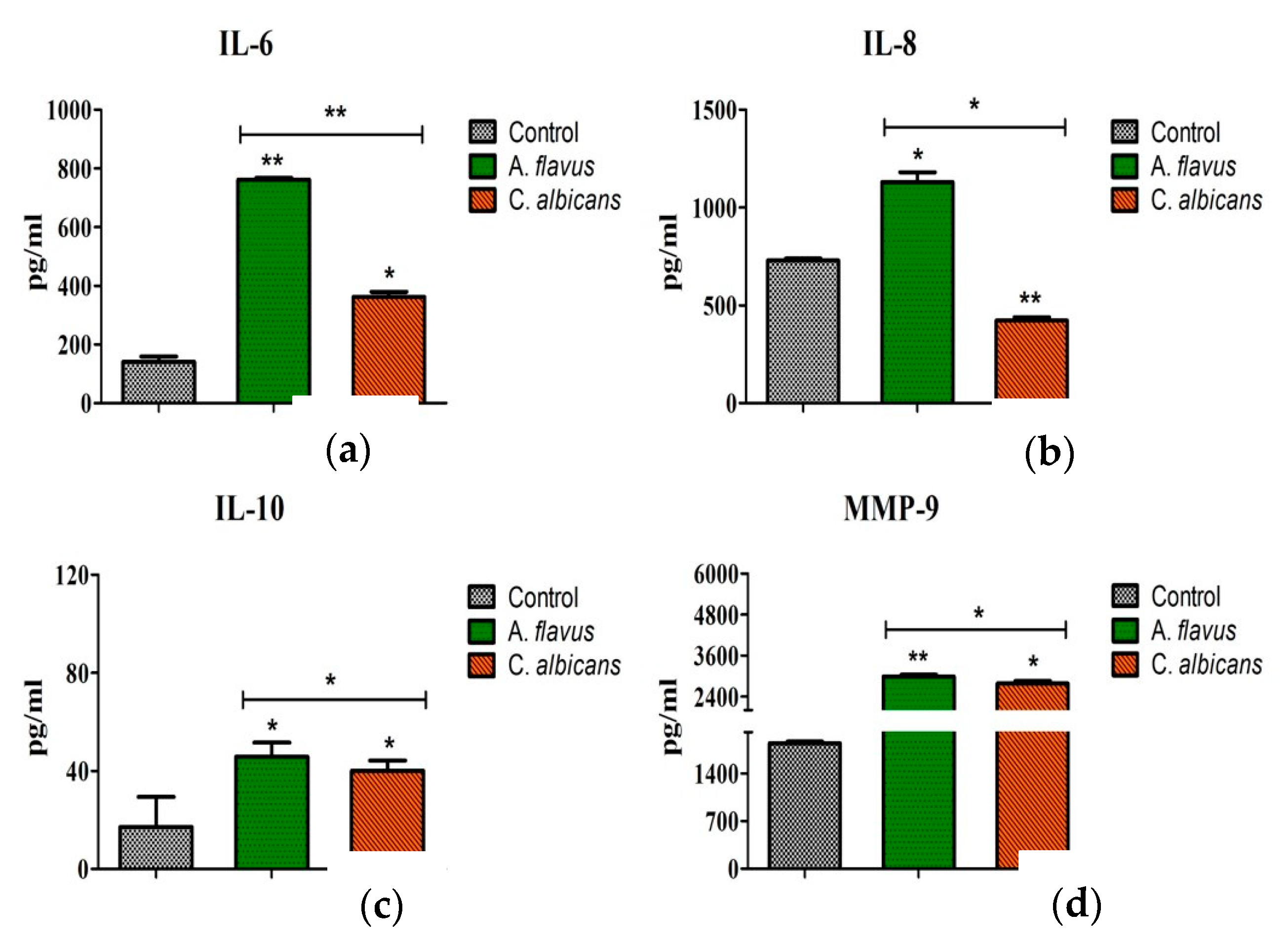
The translation of these mRNA transcripts was confirmed by assessing the protein levels of these immune mediators. Consistent with qRT-PCR data, significantly increased concentrations of IL-10 (46 pg/mL; p < 0.05), IL-6 (762 pg/mL; p < 0.005), and IL-8 (1130 pg/mL; p < 0.05) accumulated in the culture media of A. flavus stimulated CHME-3 cells at 24 h p.i. compared to C. albicans (IL-6 = 362 pg/mL; p < 0.05, IL-8 = 423 pg/mL; p < 0.005, IL-10 = 40 pg/mL; p < 0.05) (Figure 4a–c). The induction of MMP-9 levels in particular showed elevated levels in cells infected with both A. flavus and C. albicans (2993.66 ± 49.01 pg/mL vs. 2779.00 ± 74.22 pg/mL, p = 0.05) at 24 h postinfection. The protein concentrations of MMP-2 and MMP-9 were also assessed and MMP-2 was not detectable in the supernatant of both A. flavus and C. albicans challenged cells. MMP-9 however, showed a high expression in CHME-3 cells infected with A. flavus (2994 pg/mL ± 49; p < 0.005) compared to C. albicans (2779 pg/mL ± 74; p < 0.05) (Figure 4d).
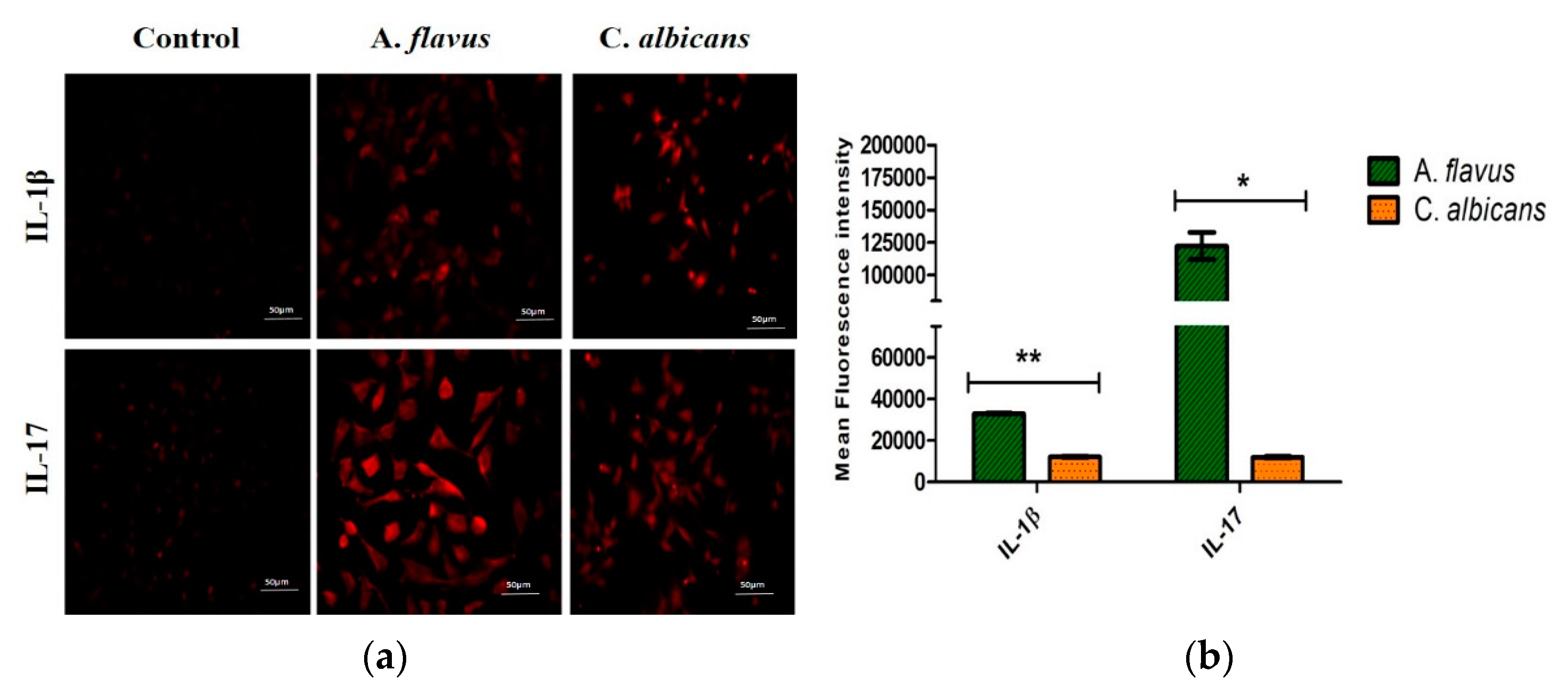
To confirm the mRNA expression observed for the remaining mediators, immunofluorescence staining was also carried out for the IL-17 and IL-1β expression in CHME-3 cells challenged with both A. flavus and C. albicans (Figure 5a), and the fluorescence intensities were compared to the uninfected cells at 12 h p.i. Similar to mRNA transcription levels, a higher protein expression of IL-17 was observed in cells infected with A. flavus compared to those infected with C. albicans. IL-1β however, showed a moderate expression in CHME-3 cells challenged with both A. flavus and C. albicans (Figure 5b).
3.5. Heatmap and PPI Network
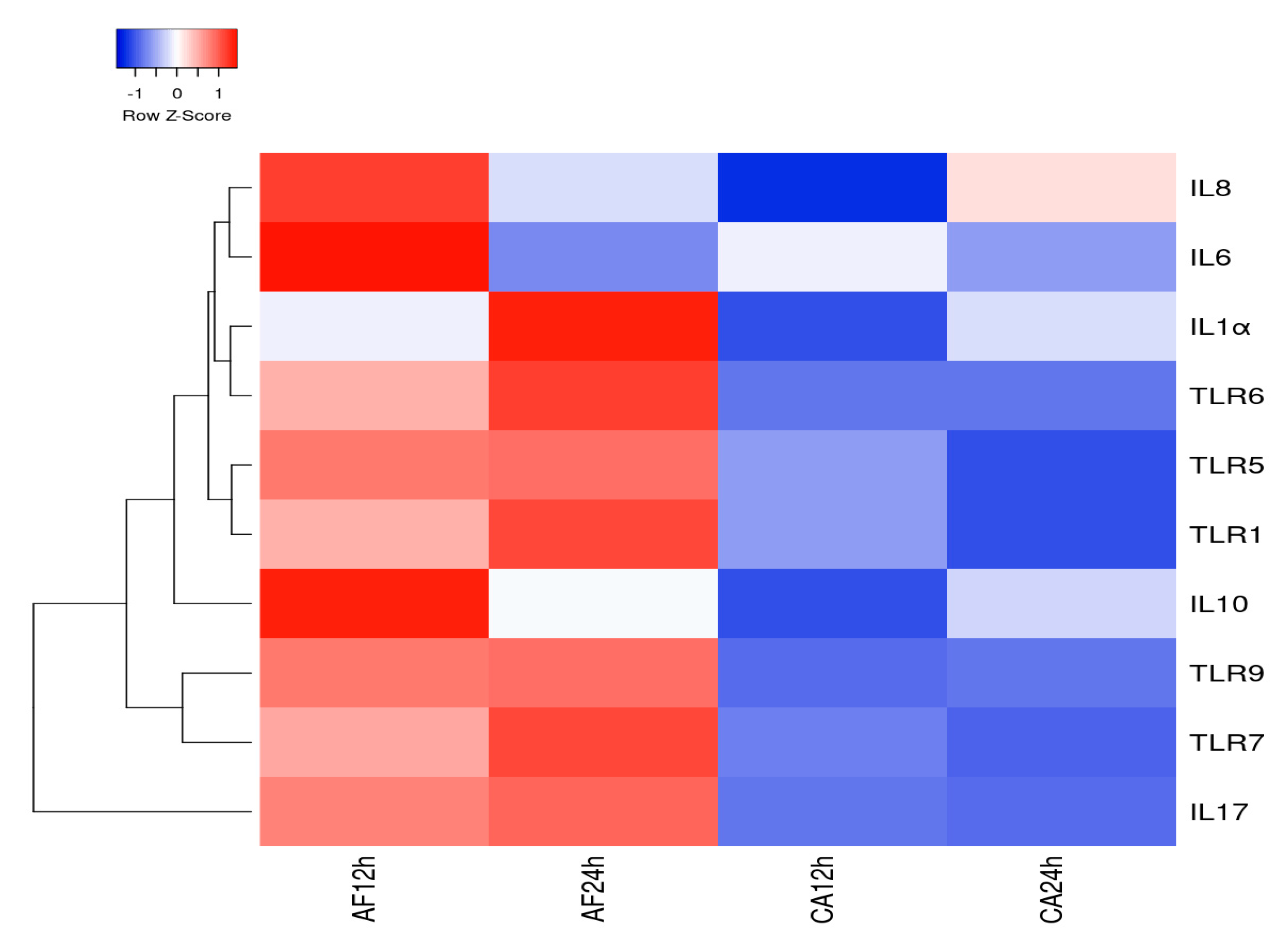
The heatmap analysis for 10 DEGs by human microglial cell infected with A. flavus and C. albicans demonstrates that the maximum gene expression was at 12 and 24 h by microglial cells challenged with A. flavus. Specifically, genes such as IL-6, IL-8 and IL-10 showed a high expression at 12 h; TLR-1, TLR-6 and IL-1α exhibited a high expression at 24 h in comparison to the other genes included in the analysis (Figure 6). However, the gene expression heatmap for C. albicans infected microglial cells revealed a lower expression level even at a higher MOI (10:1) in comparison to genes expressed by microglial cells infected with A. flavus except for IL-8 (Figure 6). Thus, there is a clear distinction in terms of the gene expression levels in microglial cells challenged with A. flavus vs. C. albicans. The hierarchical cluster analysis of real-time data was obtained using Heatmapper (http://www2.heatmapper.ca/) demonstrates that DEGs (differentially expressed genes) precisely distinguish human microglial cells (CHME-3) challenged with A. flavus from C. albicans.
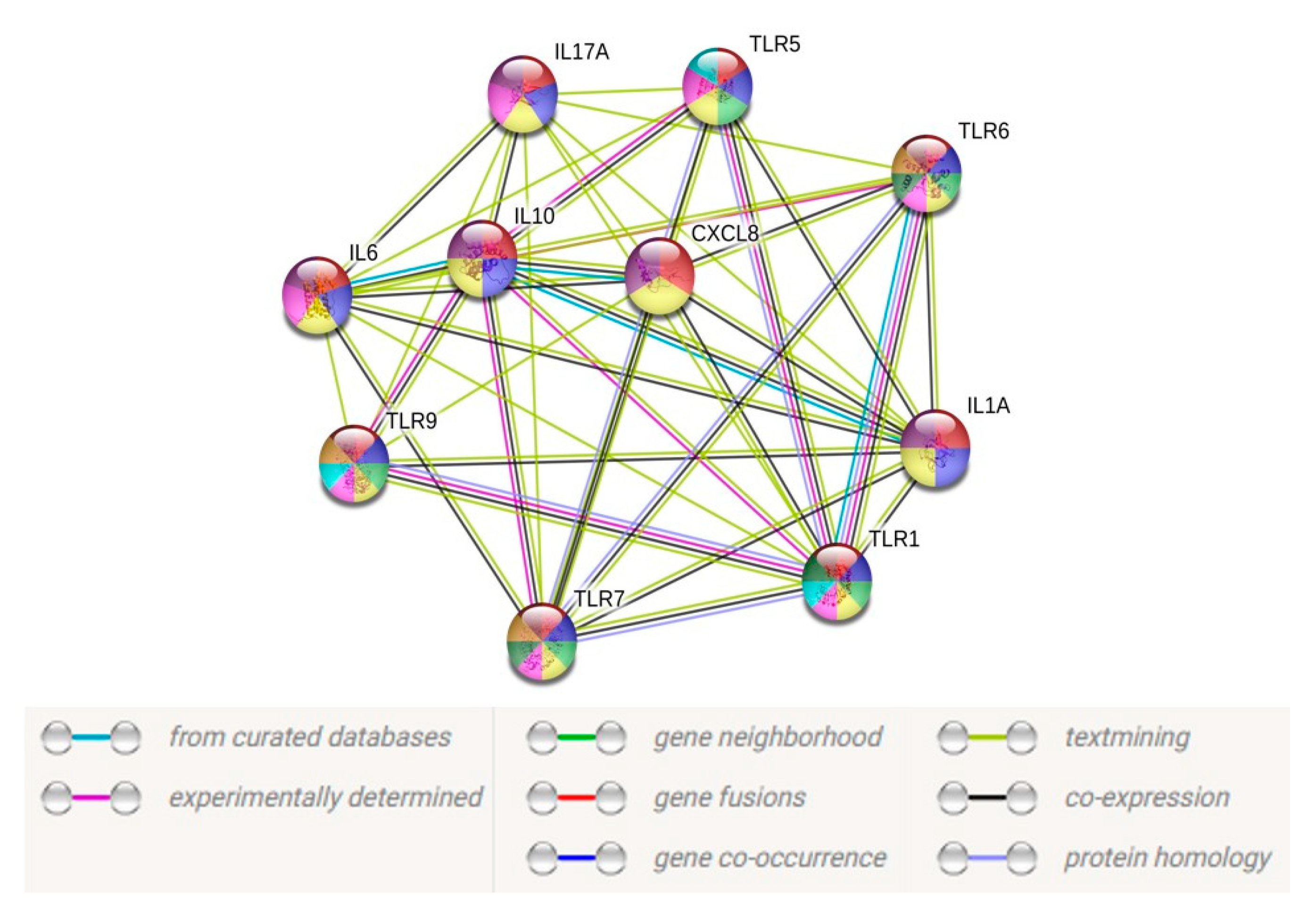
To unveil the functional facets associated with these TLRs and cytokines, we constructed a protein–protein interaction (PPI) network using the STRING database (https://string-db.org/) based on the transcript data obtained from the CHME-3 infection with A. flavus and C. albicans. Only 10 TLRs and/or cytokines with a fold change of >2-fold were utilized to construct the PPI network. By excluding the proteins without a quantified or differential expression, our resulting PPI network contained 10 nodes and 62 edges (Figure 7), with a p-value < 1.0 × 10−16, indicating a close interaction, while the PPI for CHME-3 infected with Candida sp. did not show any significant interaction due to less than five differentially expressed genes.
4. Discussion
Vision loss in fungal endophthalmitis is due to pathogen virulence factors and host-evoked inflammatory damage to the retina along with the timing and mode of interventions [22,23]. Moreover, the ocular toxicity associated with the current antifungal drugs [24], the emergence of drug resistance [25] and limited clinical experience, make the treatment of these infections challenging.
Restricted treatment options in turn alters patient management, and thus it is critical to develop better therapeutics, and to improve diagnostics and intervention strategies.
Over the last few decades, active research is being directed to understand the effect of fungal pathogens on cells as well as the host–pathogen interplay, including the secretion of cytokines and to study the cross-talk between the major signaling pathways [24,26]. We have previously demonstrated [9], using a C57BL/6 mouse model of bacterial endophthalmitis, that microglia plays an important role in initiating early innate responses to bacterial endophthalmitis and that both primary cells and the mouse-derived BV-2 microglia cell line respond the same way to infection. Continuing on from that report, we wanted to explore the responses to fungal strains isolated from patients with endophthalmitis in human microglial cell lines. Microglia are the resident mononuclear phagocytic cells and have been shown previously to play an important role in antifungal immunity in systemic fungal infections in mice by C. albicans, leading to retinal microglia activation, through the expression of TLRs [27]. However, Shah et al. [28] have reported that these microglia get activated due to the expression of dectin-1 via the recognition of fungal β-glucans, a major receptor present within fungal cell walls which in turn represses cytokine/chemokine production [29]. Hence, understanding the signaling pathways that govern the responses to both yeasts and molds in microglial cells is important. While Candida spp. remain the predominant cause of fungal endophthalmitis worldwide, in tropical countries such as India and China, Aspergillus spp. are the leading etiological agent of exogenous endophthalmitis [13]. However, the pathogenesis of Aspergillus endophthalmitis is not well studied, baring a report to evaluate the therapeutic efficacy of existing and newer antifungal agents in the eye of an animal model of A. fumigatus [24]. Keeping our focus on A. flavus, which has been described by us earlier as the most common etiology in our country [3,13], our study, for the first time, used microglia cells as a model for fungal endophthalmitis.
While the innate immune mechanisms of hosts infected with Candida albicans have already been studied, little is known about their role during endophthalmitis caused by filamentous fungi, A. flavus. In this study, we have shown that there would be a differential immune response to yeast and filamentous fungi by microglia cells. This difference in innate response among fungal pathogens is primarily speculated to involve cell wall components and their interaction with host cells and tissues, favoring fungal growth and allowing dissemination in the host [30]. The outer layer of the C. albicans cell wall is said to be packed with mannoproteins and cross-linked to β-1,6-glucans and, unlike Aspergillus spp., α-glucan is absent from Candida spp. cell walls [31]. C. albicans has also been hypothesized to initiate its own uptake by host cells via complement proteins or complement receptors as an alternative immune evasion strategy [32]. It has also been reported that these mannoproteins are glycosylphosphatidylinositol-modified during interactions with the host, allowing the activation of the immune response by masking the β-glucan layer and thereby leading to an ineffective activation of the host immune system [31]. Similarly, it has been reported that C. albicans tended to cause higher inflammatory cytokine and chemokine concentrations, along with increased neutrophils and inflammatory monocytes, compared with nonalbicans Candida infections [33]. Additionally, the MAPK signaling pathway was also demonstrated by Galan-Diéz et al. [30] to be involved in the process of β-glucan masking. Comparatively, the cell wall of Aspergillus sp. is a two-layered structure, consisting of a polymer of β-1,3-glucan, cross-linked to α-1,3-glucan, galactomannan, galactosaminogalactan and chitin, all of them covalently bound one to the other. Galactosaminogalactan, an adhesin that facilitates the binding of hyphae to macrophages, neutrophils, and platelets has been reported to be associated with masking cell wall β-glucans from recognition by dectin-1, decreased polymorphonuclear neutrophil apoptosis via an NK cell-dependent mechanism and ROS production. It is also required for stimulating IL-17 production by neutrophils as well as IL-1α, IL-12, and TNFα release as also seen in our study [32].
The presence of fungal-derived ligands following a direct recognition by Pattern Recognition Receptors (PRRs) through TLRs has also been reported, as has the involvement of TLR-2 and -4 in the mediation of host responses during systemic candidiasis as well as in corneal infections, which instigates the production of inflammatory cytokines [27,34,35]. Our results, however, show the expression of only TLR-2, on the surface of microglia by C. albicans which recognizes the carbohydrates such as mannose and β-glucans on the surface of C. albicans [34], while cells infected with A. flavus demonstrate then activation of several TLRs including TLR-1, -2, -5, -7 and TLR-9 have an important role in the initial interaction of this fungus with the host cells. While, TLR-9 is known to modulate the immune response towards conidia A. fumigatus, which in turn is specific for dectin-1 expression and the activation of IL-17 [36]. In addition, TLR-4 was also expressed by microglial cells infected with A. flavus at the 3 h time point (data not shown) but wasn’t expressed in cells infected with C. albicans. The initial expression of TLR-4 by A. flavus corroborates with reports that suggest only TLR-4-mediated proinflammatory signals (and not TLR-2 induced anti-inflammatory signals), are lost on Aspergillus germination to hyphae. Our findings also corroborated with studies showing initiation of an immune response via TLR signaling in the murine model of Aspergillus fumigatus endophthalmitis [37,38]. TLR-2 has been reported to be induced by unidentified ligands present on both hyphal and conidia forms of A. fumigatus, whereas ligands for only conidial forms are reported by TLR-4 [39,40]. Another study found the α-glucan of Candida sp. to play a major role in TLR2-mediated attenuation of the immune response, while TLR4-attenuation was caused by the presence of the α-, β-glucan, and galactomannan components of the Aspergillus cell wall [41]. Our findings are also in contrast to Koutsouras et al., who reported an induction of a proinflammatory response that is critical in the protection against fungi via TLR-4 signaling on the surface of microglial cells [10]. This could be due to the time points chosen in the study and perhaps an earlier time point could reveal the presence of TLR-4 activation. This indicates that TLRs discriminate between distinct fungal morphotypes [40].
In addition, Aspergillus hyphae, but not Aspergillus conidia, stimulated the production of IL-10 through TLR2-dependent mechanisms [40], as was also shown by the high expression of IL-10 by the 12 h time point. The conidia of A. flavus in turn lead to the activation of inflammatory responses by secreting proinflammatory cytokines, particularly, the cytokines IL-1α, IL-6, IL-8 and IL-17 as well as the anti-inflammatory cytokine IL-10. Comparatively, only IL-6 and IL-10 were expressed slightly in cells infected with C. albicans in our study. While there is limited knowledge on the modulation of anti-inflammatory processes by microglia in the setting of fungal infections, a previous study had shown that, an infection with C. albicans (in mice) induces immunosuppression by activating TLR-2, leading to the release of IL-10 [42]. Additionally, we noticed that A. flavus causes a downregulation in the levels of GM-CSF and IFN-γ in microglia cells, which was similar to the findings by Schneider et al. [43] who reported that A. fumigatus actively impairs the release of IFN-γ (and M-CSF) by NK cells. In contrast to the results obtained regarding the cytokine expression in the murine model of Aspergillus endophthalmitis [14], where the production of several inflammatory mediators including IL-1β, IL6, and CXCL2 were reported, we observed a nonsignificant downregulation of IFN-γ and IL-1β. Numerous observations that IL-17 contributes to fungal protection have been obtained from studies with experimental models of infection [44]. However, the exact role of IL-17 in controlling these fungi in humans is yet to be determined. Our study shows the overexpression of IL-17 in A. flavus infected cells but a downregulation in cells infected with C. albicans. Our observations of microglia cells’ impaired immune response when infected with C. albicans could be due to biofilm formation as described previously, which seems to protect C. albicans from microglial damage by impairment of fungal cell phagocytosis, the release of cytokines and chemokines, and NO production [42]. Although the phenotypic variations and activation of microglial cells are complicated, they are mainly said to be involved in anti-inflammatory and/or proinflammatory processes, depending on the infectious stimuli, and get polarized into multiple phenotypes or switch activity states [45]. Humans with a MyD88 signaling defect do not have a higher susceptibility to fungal infections [46]. However, TLR polymorphisms alters signaling pathways, thereby increasing the risk of fungal infections [47]. Some reports also suggests that A. fumigatus causes the activation of the TLR9/BTK/calcineurin/NFAT signaling pathway, causing the activation of several proinflammatory mediators, such as TNF-α, leading to early neutrophil recruitment, and fungal clearance [47]. A major limitation of our study is that it is an in vitro study with a human, immortalized CNS-derived microglial cell line, and while the characteristics of brain- and retinal-derived microglial cells are similar, the environments in which they reside as well as pathophysiological conditions might differ. It is clear that antigenic, structural, and transcriptional differences exist between cohorts of microglia [48]. IL-34 appears to be exclusive to retinal microglia and different levels of CD11c, CD11b, and TLR4, which have been shown in microglia cells of retinal origin. While Huang et al. reported that repopulated retinal microglia might share a similar origin as the brain [49], Hammond et al. compared the transcriptional data of retinal microglia to brain microglia and found a similar set of lineage-specific factors shared by both populations, suggesting that developing retina and brain microglia may be ontogenically similar [50]. Additionally, O’Koren et al. [51] have reported functional differences by niche and IL-34 dependency across two anatomically distinct locations in the retina during homeostasis. Consequently, their close communication with immune cells, in conjunction with their ability to tailor their function specifically to the surrounding milieu, we believe that differential inflammatory expression shown by our fungal isolates in brain microglial cells would be applicable in retinal microglial cells as well. Additionally, establishing pure primary retinal microglial cultures from human tissues is rather challenging and probably impossible, but considering the importance of Muller and microglial cell interactions in microglial cell signaling, it would also be of considerable importance to perform these studies with cocultures of microglial and Muller cells and future work could focus on this aspect.
5. Conclusions
In conclusion, we demonstrated that A. flavus infection initiates a host immune response by the activation of several TLRs, followed by an induction of inflammatory cytokines whereas C. albicans infection in microglia demonstrated an impaired level of proinflammatory cytokine release. Therefore, taking into account our results, we propose that microglia may be inefficient for the clearance of C. albicans from the retina during endophthalmitis. Conidia and yeasts have a differential immune response and understanding this and other mechanisms of inflammation will further our ability to initiate strategies of immunomodulation, to minimize the retinal damage associated with endophthalmitis. Thus, this study advances the role using immunomodulators such as natural extracts and essential oils [52,53,54] or phytochemicals in mitigating the impact of inflammatory molecules as some compounds have already been shown to be effective in the treatment of multidrug resistance infections [55]. Similarly, a commercial ophthalmic solution containing PVP-I 0.6% [56] and the more recently approved keratosept, containing hexamidine diisethionate 0.05%, used to aid in a faster re-epithelization, showed rapid antifungal activity against different Candida strains [57], and also represent promising compounds for adjuvant therapy. To the best of our knowledge, this is the first in vitro model of A. flavus induced infection and further detailed studies are required to understand the microglia interactions with the retina and thus contribute to better management of patients affected by Aspergillus endophthalmitis.
Supplementary Materials
The following are available online at https://0-www-mdpi-com.brum.beds.ac.uk/2309-608X/6/3/162/s1, Table S1: List of Human primers (TLRs, Cytokines/chemokine, MMPs and TIMP) used for qRT-PCR analysis.
Author Contributions
Conceptualization, J.J. methodology, J.J.; software, J.G.; validation, J.G., and P.N.; formal analysis, J.G.; investigation, J.G.; resources, I.K.; data curation, J.G., and P.N.; writing—original draft preparation, J.G.; writing—review and editing, J.J., I.K., and A.K.; visualization, J.J., and A.K.; supervision, J.J.; project administration, J.J.; funding acquisition, J.J. All authors have read and agreed to the published version of the manuscript.
Funding
This research was funded by grant from DST-SERB to J.J. (File Number: CRG/2019/004386) India.
Acknowledgments
Hyderabad Eye Research Foundation for funding support. The authors also thank Inderjeet Kaur for providing the human microglial cell line (CHME3) and Vivek Singh (CORE, L V Prasad Eye Institute, Hyderabad, India) for helping with a validation experiment using an immunofluorescence assay.
Conflicts of Interest
The authors declare no conflict of interest. The funders had no role in the design of the study; in the collection, analyses, or interpretation of data; in the writing of the manuscript, or in the decision to publish the results.
References
- Durand, M.L. Endophthalmitis. Clin. Microbiol. Infect. 2013, 19, 227–234. [Google Scholar] [CrossRef] [Green Version]
- Kunimoto, D.Y.; Das, T.; Sharma, S.; Jalali, S.; Majji, A.B.; Gopinathan, U.; Athmanathan, S.; Rao, T.N. Microbiologic spectrum and susceptibility of isolates: Part I. Postoperative endophthalmitis. Endophthalmitis Research Group. Am. J. Ophthalmol. 1999, 128, 240–242. [Google Scholar] [CrossRef]
- Gandhi, J.; Jayasudha, R.; Naik, P.; Sharma, S.; Dave, V.P.; Joseph, J. Targeted high-throughput sequencing identifies predominantly fungal pathogens in patients with clinically infectious, culture-negative endophthalmitis in South India. Microorganisms 2019, 7, 411. [Google Scholar] [CrossRef] [Green Version]
- Chakrabarti, A.; Shivaprakash, M.R.; Singh, R.; Tarai, B.; George, V.K.; Fomda, B.A.; Gupta, A. Fungal endophthalmitis: Fourteen years’ experience from a center in India. Retina 2008, 28, 1400–1407. [Google Scholar] [CrossRef]
- Usai, D.; Donadu, M.; Bua, A.; Molicotti, P.; Zanetti, S.; Piras, S.; Corona, P.; Ibba, R.; Carta, A. Enhancement of antimicrobial activity of pump inhibitors associating drugs. J. Infect. Dev. Ctries. 2019, 13, 162–164. [Google Scholar] [CrossRef] [Green Version]
- Bui, D.K.; Carvounis, P.E. Evidence for and against intravitreous corticosteroids in addition to intravitreous antibiotics for acute endophthalmitis. Int. Ophthalmol. Clin. 2014, 54, 215–224. [Google Scholar] [CrossRef] [PubMed]
- Driscoll, K.E. TNFalpha and MIP-2: Role in particle-induced inflammation and regulation by oxidative stress. Toxicol. Lett. 2000, 112–113, 177–183. [Google Scholar] [CrossRef]
- Hernandez-Chavez, M.J.; Perez-Garcia, L.A.; Nino-Vega, G.A.; Mora-Montes, H.M. Fungal strategies to evade the host immune recognition. J. Fungi (Basel) 2017, 3, 51. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kochan, T.; Singla, A.; Tosi, J.; Kumar, A. Toll-like receptor 2 ligand pretreatment attenuates retinal microglial inflammatory response but enhances phagocytic activity toward Staphylococcus aureus. Infect. Immun. 2012, 80, 2076–2088. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Koutsouras, G.W.; Ramos, R.L.; Martinez, L.R. Role of microglia in fungal infections of the central nervous system. Virulence 2017, 8, 705–718. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vallejo-Garcia, J.L.; Asencio-Duran, M.; Pastora-Salvador, N.; Vinciguerra, P.; Romano, M.R. Role of inflammation in endophthalmitis. Mediat. Inflamm. 2012, 2012, 196094. [Google Scholar] [CrossRef] [PubMed]
- Li, L.; Eter, N.; Heiduschka, P. The microglia in healthy and diseased retina. Exp. Eye Res. 2015, 136, 116–130. [Google Scholar] [CrossRef] [PubMed]
- Joseph, J.; Sontam, B.; Guda, S.J.M.; Gandhi, J.; Sharma, S.; Tyagi, M.; Dave, V.P.; Das, T. Trends in microbiological spectrum of endophthalmitis at a single tertiary care ophthalmic hospital in India: A review of 25 years. Eye (London) 2019, 33, 1090–1095. [Google Scholar] [CrossRef] [PubMed]
- Gupta, N.; Singh, P.K.; Revankar, S.G.; Chandrasekar, P.H.; Kumar, A. Pathobiology of Aspergillus fumigatus endophthalmitis in immunocompetent and immunocompromised mice. Microorganisms 2019, 7, 297. [Google Scholar] [CrossRef] [Green Version]
- Cramer, R.A.; Rivera, A.; Hohl, T.M. Immune responses against Aspergillus fumigatus: What have we learned? Curr. Opin. Infect. Dis. 2011, 24, 315–322. [Google Scholar] [CrossRef] [Green Version]
- Bozza, S.; Clavaud, C.; Giovannini, G.; Fontaine, T.; Beauvais, A.; Sarfati, J.; D’Angelo, C.; Perruccio, K.; Bonifazi, P.; Zagarella, S.; et al. Immune sensing of Aspergillus fumigatus proteins, glycolipids, and polysaccharides and the impact on Th immunity and vaccination. J. Immunol. 2009, 183, 2407–2414. [Google Scholar] [CrossRef] [Green Version]
- Dello Russo, C.; Cappoli, N.; Coletta, I.; Mezzogori, D.; Paciello, F.; Pozzoli, G.; Navarra, P.; Battaglia, A. The human microglial HMC3 cell line: Where do we stand? A systematic literature review. J. Neuroinflamm. 2018, 15, 259. [Google Scholar] [CrossRef] [Green Version]
- Meletiadis, J.; Meis, J.F.; Mouton, J.W.; Verweij, P.E. Analysis of growth characteristics of filamentous fungi in different nutrient media. J. Clin. Microbiol. 2001, 39, 478–484. [Google Scholar] [CrossRef] [Green Version]
- Jensen, E.C. Quantitative analysis of histological staining and fluorescence using ImageJ. Anat. Rec. 2013, 296, 378–381. [Google Scholar] [CrossRef]
- Babicki, S.; Arndt, D.; Marcu, A.; Liang, Y.; Grant, J.R.; Maciejewski, A.; Wishart, D.S. Heatmapper: Web-enabled heat mapping for all. Nucleic Acids Res. 2016, 44, W147–W153. [Google Scholar] [CrossRef]
- Szklarczyk, D.; Franceschini, A.; Kuhn, M.; Simonovic, M.; Roth, A.; Minguez, P.; Doerks, T.; Stark, M.; Muller, J.; Bork, P.; et al. The STRING database in 2011: Functional interaction networks of proteins, globally integrated and scored. Nucleic Acids Res. 2011, 39, D561–D568. [Google Scholar] [CrossRef] [PubMed]
- Durand, M.L. Bacterial and fungal endophthalmitis. Clin. Microbiol. Rev. 2017, 30, 597–613. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Essman, T.F.; Flynn, H.W., Jr.; Smiddy, W.E.; Brod, R.D.; Murray, T.G.; Davis, J.L.; Rubsamen, P.E. Treatment outcomes in a 10-year study of endogenous fungal endophthalmitis. Ophthalmic Surg. Lasers Imaging Retin. 1997, 28, 185–194. [Google Scholar]
- Guest, J.M.; Singh, P.K.; Revankar, S.G.; Chandrasekar, P.H.; Kumar, A. Isavuconazole for treatment of experimental fungal endophthalmitis caused by Aspergillus fumigatus. Antimicrob. Agents Chemother. 2018, 62. [Google Scholar] [CrossRef] [Green Version]
- Howard, S.J.; Cerar, D.; Anderson, M.J.; Albarrag, A.; Fisher, M.C.; Pasqualotto, A.C.; Laverdiere, M.; Arendrup, M.C.; Perlin, D.S.; Denning, D.W. Frequency and evolution of Azole resistance in Aspergillus fumigatus associated with treatment failure. Emerg. Infect. Dis. 2009, 15, 1068–1076. [Google Scholar] [CrossRef]
- Horn, F.; Heinekamp, T.; Kniemeyer, O.; Pollmacher, J.; Valiante, V.; Brakhage, A.A. Systems biology of fungal infection. Front. Microbiol. 2012, 3, 108. [Google Scholar] [CrossRef] [Green Version]
- Maneu, V.; Noailles, A.; Megias, J.; Gomez-Vicente, V.; Carpena, N.; Gil, M.L.; Gozalbo, D.; Cuenca, N. Retinal microglia are activated by systemic fungal infection. Investig. Ophthalmol. Vis. Sci. 2014, 55, 3578–3585. [Google Scholar] [CrossRef]
- Shah, V.B.; Huang, Y.; Keshwara, R.; Ozment-Skelton, T.; Williams, D.L.; Keshvara, L. Beta-glucan activates microglia without inducing cytokine production in Dectin-1-dependent manner. J. Immunol. 2008, 180, 2777–2785. [Google Scholar] [CrossRef] [Green Version]
- Shah, V.B.; Williams, D.L.; Keshvara, L. beta-Glucan attenuates TLR2- and TLR4-mediated cytokine production by microglia. Neurosci. Lett. 2009, 458, 111–115. [Google Scholar] [CrossRef] [Green Version]
- Galan-Diez, M.; Arana, D.M.; Serrano-Gomez, D.; Kremer, L.; Casasnovas, J.M.; Ortega, M.; Cuesta-Dominguez, A.; Corbi, A.L.; Pla, J.; Fernandez-Ruiz, E. Candida albicans beta-glucan exposure is controlled by the fungal CEK1-mediated mitogen-activated protein kinase pathway that modulates immune responses triggered through dectin-1. Infect. Immun. 2010, 78, 1426–1436. [Google Scholar] [CrossRef] [Green Version]
- Garcia-Rubio, R.; de Oliveira, H.C.; Rivera, J.; Trevijano-Contador, N. The fungal cell wall: Candida, Cryptococcus, and Aspergillus species. Front. Microbiol. 2019, 10, 2993. [Google Scholar] [CrossRef] [PubMed]
- Singh, D.K.; Toth, R.; Gacser, A. Mechanisms of pathogenic Candida species to evade the host complement attack. Front. Cell. Infect. Microbiol. 2020, 10, 94. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Abe, M.; Kinjo, Y.; Ueno, K.; Takatsuka, S.; Nakamura, S.; Ogura, S.; Kimura, M.; Araoka, H.; Sadamoto, S.; Shinozaki, M.; et al. Differences in ocular complications between Candida albicans and non-albicans Candida infection analyzed by epidemiology and a mouse ocular candidiasis model. Front. Microbiol. 2018, 9, 2477. [Google Scholar] [CrossRef] [PubMed]
- Romani, L. Immunity to fungal infections. Nat. Rev. Immunol. 2004, 4, 1–23. [Google Scholar] [CrossRef]
- Yang, I.; Han, S.J.; Kaur, G.; Crane, C.; Parsa, A.T. The role of microglia in central nervous system immunity and glioma immunology. J. Clin. Neurosci. 2010, 17, 6–10. [Google Scholar] [CrossRef] [Green Version]
- Ramaprakash, H.; Ito, T.; Standiford, T.J.; Kunkel, S.L.; Hogaboam, C.M. Toll-like receptor 9 modulates immune responses to Aspergillus fumigatus conidia in immunodeficient and allergic mice. Infect. Immun. 2009, 77, 108–119. [Google Scholar] [CrossRef] [Green Version]
- Bourgeois, C.; Kuchler, K. Fungal pathogens-a sweet and sour treat for toll-like receptors. Front. Cell. Infect. Microbiol. 2012, 2, 142. [Google Scholar] [CrossRef] [Green Version]
- Netea, M.G.; Gow, N.A.; Munro, C.A.; Bates, S.; Collins, C.; Ferwerda, G.; Hobson, R.P.; Bertram, G.; Hughes, H.B.; Jansen, T.; et al. Immune sensing of Candida albicans requires cooperative recognition of mannans and glucans by lectin and Toll-like receptors. J. Clin. Investig. 2006, 116, 1642–1650. [Google Scholar] [CrossRef]
- Netea, M.G.; Warris, A.; Van der Meer, J.W.; Fenton, M.J.; Verver-Janssen, T.J.; Jacobs, L.E.; Andresen, T.; Verweij, P.E.; Kullberg, B.J. Aspergillus fumigatus evades immune recognition during germination through loss of toll-like receptor-4-mediated signal transduction. J. Infectig. Dis. 2003, 188, 320–326. [Google Scholar] [CrossRef] [Green Version]
- van de Veerdonk, F.L.; Kullberg, B.J.; van der Meer, J.W.; Gow, N.A.; Netea, M.G. Host-microbe interactions: Innate pattern recognition of fungal pathogens. Curr. Opin. Microbiol. 2008, 11, 305–312. [Google Scholar] [CrossRef]
- Chai, L.Y.; Vonk, A.G.; Kullberg, B.J.; Verweij, P.E.; Verschueren, I.; van der Meer, J.W.; Joosten, L.A.; Latge, J.P.; Netea, M.G. Aspergillus fumigatus cell wall components differentially modulate host TLR2 and TLR4 responses. Microbes Infect. 2011, 13, 151–159. [Google Scholar] [CrossRef]
- Orsi, C.F.; Borghi, E.; Colombari, B.; Neglia, R.G.; Quaglino, D.; Ardizzoni, A.; Morace, G.; Blasi, E. Impact of Candida albicans hyphal wall protein 1 (HWP1) genotype on biofilm production and fungal susceptibility to microglial cells. Microb. Pathog. 2014, 69–70, 20–27. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Schneider, A.; Blatzer, M.; Posch, W.; Schubert, R.; Lass-Florl, C.; Schmidt, S.; Lehrnbecher, T. Aspergillus fumigatus responds to natural killer (NK) cells with upregulation of stress related genes and inhibits the immunoregulatory function of NK cells. Oncotarget 2016, 7, 71062–71071. [Google Scholar] [CrossRef] [PubMed]
- Mengesha, B.G.; Conti, H.R. The role of IL-17 in protection against mucosal Candida infections. J. Fungi (Basel) 2017, 3, 52. [Google Scholar] [CrossRef]
- Hanisch, U.K.; Kettenmann, H. Microglia: Active sensor and versatile effector cells in the normal and pathologic brain. Nat. Neurosci. 2007, 10, 1387–1394. [Google Scholar] [CrossRef] [PubMed]
- Salazar, F.; Brown, G.D. Antifungal innate immunity: A perspective from the last 10 years. J. Innate Immun. 2018, 10, 373–397. [Google Scholar] [CrossRef]
- Romani, L. Immunity to fungal infections. Nat. Rev. Immunol. 2011, 11, 275–288. [Google Scholar] [CrossRef]
- Li, F.; Jiang, D.; Samuel, M.A. Microglia in the developing retina. Neural Dev. 2019, 14, 12. [Google Scholar] [CrossRef] [Green Version]
- Huang, Y.; Xu, Z.; Xiong, S.; Qin, G.; Sun, F.; Yang, J.; Yuan, T.F.; Zhao, L.; Wang, K.; Liang, Y.X.; et al. Dual extra-retinal origins of microglia in the model of retinal microglia repopulation. Cell Discov. 2018, 4, 9. [Google Scholar] [CrossRef] [Green Version]
- Hammond, T.R.; Dufort, C.; Dissing-Olesen, L.; Giera, S.; Young, A.; Wysoker, A.; Walker, A.J.; Gergits, F.; Segel, M.; Nemesh, J.; et al. Single-cell RNA sequencing of microglia throughout the mouse lifespan and in the injured brain reveals complex cell-state changes. Immunity 2019, 50, 253–271.e2. [Google Scholar] [CrossRef] [Green Version]
- O’Koren, E.G.; Yu, C.; Klingeborn, M.; Wong, A.Y.W.; Prigge, C.L.; Mathew, R.; Kalnitsky, J.; Msallam, R.A.; Silvin, A.; Kay, J.N.; et al. Microglial function is distinct in different anatomical locations during retinal homeostasis and degeneration. Immunity 2019, 50, 723–737.e7. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cannas, S.; Usai, D.; Pinna, A.; Benvenuti, S.; Tardugno, R.; Donadu, M.; Zanetti, S.; Kaliamurthy, J.; Molicotti, P. Essential oils in ocular pathology: An experimental study. J. Infect. Dev. Ctries. 2015, 9, 650–654. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Donadu, M.G.; Usai, D.; Marchetti, M.; Usai, M.; Mazzarello, V.; Molicotti, P.; Montesu, M.A.; Delogu, G.; Zanetti, S. Antifungal activity of oils macerates of North Sardinia plants against Candida species isolated from clinical patients with candidiasis. Nat. Prod. Res. 2019, 1–5. [Google Scholar] [CrossRef] [PubMed]
- Bua, A.; Usai, D.; Donadu, M.G.; Delgado Ospina, J.; Paparella, A.; Chaves-Lopez, C.; Serio, A.; Rossi, C.; Zanetti, S.; Molicotti, P. Antimicrobial activity of Austroeupatorium inulaefolium (H.B.K.) against intracellular and extracellular organisms. Nat. Prod. Res. 2018, 32, 2869–2871. [Google Scholar] [CrossRef]
- Chaves-Lopez, C.; Usai, D.; Donadu, M.G.; Serio, A.; Gonzalez-Mina, R.T.; Simeoni, M.C.; Molicotti, P.; Zanetti, S.; Pinna, A.; Paparella, A. Potential of Borojoa patinoi Cuatrecasas water extract to inhibit nosocomial antibiotic resistant bacteria and cancer cell proliferation in vitro. Food Funct. 2018, 9, 2725–2734. [Google Scholar] [CrossRef]
- Pinna, A.; Donadu, M.G.; Usai, D.; Dore, S.; D’Amico-Ricci, G.; Boscia, F.; Zanetti, S. In vitro antimicrobial activity of a new ophthalmic solution containing povidone-iodine 0.6% (IODIM®). Acta Ophthalmol. 2020, 98, e178–e180. [Google Scholar] [CrossRef]
- Pinna, A.; Donadu, M.G.; Usai, D.; Dore, S.; Boscia, F.; Zanetti, S. In vitro antimicrobial activity of a new ophthalmic solution containing hexamidine diisethionate 0.05% (Keratosept). Cornea 2020. [Google Scholar] [CrossRef]
Figure 1.
CHME-3 cells were challenged with A. flavus and C. albicans at indicated time points. Postinfection human microglial total RNA was subjected to RT-PCR and qRT-PCR using SYBR green assay for Toll-Like Receptors (TLRs) (1-7 and -9) and β-actin gene was used as internal control and uninfected CHME-3 cells were taken as control. The data were shown as the mean ± Standard error (SE) from three sets of independent experiments and * p < 0.05, ** p < 0.005.
Figure 1.
CHME-3 cells were challenged with A. flavus and C. albicans at indicated time points. Postinfection human microglial total RNA was subjected to RT-PCR and qRT-PCR using SYBR green assay for Toll-Like Receptors (TLRs) (1-7 and -9) and β-actin gene was used as internal control and uninfected CHME-3 cells were taken as control. The data were shown as the mean ± Standard error (SE) from three sets of independent experiments and * p < 0.05, ** p < 0.005.

Figure 2.
CHME-3 cells were challenged with A. flavus and C. albicans at indicated time points. Postinfection human microglial total RNA was subjected to RT-PCR and qRT-PCR using SYBR green assay with cytokine-specific primes. Representative RT-PCRs of each cytokines were shown. Human microglia with no fungal infection cDNA was used as control. β-actin was used as an endogenous control. Data are from n = 3 independent experiments done in triplicate. * p < 0.05, ** p< 0.005 compared to control.
Figure 2.
CHME-3 cells were challenged with A. flavus and C. albicans at indicated time points. Postinfection human microglial total RNA was subjected to RT-PCR and qRT-PCR using SYBR green assay with cytokine-specific primes. Representative RT-PCRs of each cytokines were shown. Human microglia with no fungal infection cDNA was used as control. β-actin was used as an endogenous control. Data are from n = 3 independent experiments done in triplicate. * p < 0.05, ** p< 0.005 compared to control.

Figure 3.
CHME-3 cells were challenged with A. flavus and C. albicans at indicated time points. The bar graphs show that there is no significant expression of (a) Tissue Inhibitor of Metalloproteinase-1 (TIMP-1) and (b) Matrix metalloproteinase-2 (MMP-2) in CHME-3 cells infected with A. flavus and C. albicans. (c) Significantly elevated expression of MMP-9 was observed in CHME-3 cells challenged with A. flavus and C. albicans compared with control. Data shown are the mean standard errors of three independent experiments performed in triplicate ** p < 0.005.
Figure 3.
CHME-3 cells were challenged with A. flavus and C. albicans at indicated time points. The bar graphs show that there is no significant expression of (a) Tissue Inhibitor of Metalloproteinase-1 (TIMP-1) and (b) Matrix metalloproteinase-2 (MMP-2) in CHME-3 cells infected with A. flavus and C. albicans. (c) Significantly elevated expression of MMP-9 was observed in CHME-3 cells challenged with A. flavus and C. albicans compared with control. Data shown are the mean standard errors of three independent experiments performed in triplicate ** p < 0.005.

Figure 4.
CHME-3 cells were infected with A. flavus and C. albicans. Production of (a) IL-6, (b) IL-8, (c) IL-10 and (d) MMP-9 were measured in cell-free supernatants after 24 h at 37 °C by ELISA. Values shown are the mean ± SE of three independent experiments performed in triplicate. Significant variations in production of cytokines and MMPs were determined by Student’s t-test. * p < 0.05, ** p < 0.005.
Figure 4.
CHME-3 cells were infected with A. flavus and C. albicans. Production of (a) IL-6, (b) IL-8, (c) IL-10 and (d) MMP-9 were measured in cell-free supernatants after 24 h at 37 °C by ELISA. Values shown are the mean ± SE of three independent experiments performed in triplicate. Significant variations in production of cytokines and MMPs were determined by Student’s t-test. * p < 0.05, ** p < 0.005.

Figure 5.
(a) Representative images of microglial cells infected with A. flavus and C. albicans were stained with anti-IL-17 and anti-IL-1β, labeled with Alexa 594 (red) after 12 h p.i. and uninfected CHME-3 cells were taken as control. Magnification 20×. (b) The bar graph depicting cell fluorescence intensity (normalized to background) in microglial cells challenged with A. flavus and C. albicans measured using NIH Image J software. Relative immunofluorescence staining was quantified from randomly selected fields. Values represent the mean fluorescence intensity ± SE (gray-value of 16-bit images). * p < 0.05, ** p < 0.005.
Figure 5.
(a) Representative images of microglial cells infected with A. flavus and C. albicans were stained with anti-IL-17 and anti-IL-1β, labeled with Alexa 594 (red) after 12 h p.i. and uninfected CHME-3 cells were taken as control. Magnification 20×. (b) The bar graph depicting cell fluorescence intensity (normalized to background) in microglial cells challenged with A. flavus and C. albicans measured using NIH Image J software. Relative immunofluorescence staining was quantified from randomly selected fields. Values represent the mean fluorescence intensity ± SE (gray-value of 16-bit images). * p < 0.05, ** p < 0.005.

Figure 6.
Gene expression heatmap constructed employing heatmapper.ca for 10 differentially expressed genes in human microglial cells challenged with A. flavus (AF) and C. albicans (CA) at 12 and 24 h p.i. The cluster value is coded with a range of colors from blue (for the lowest expression level) to red (for the highest expression level).
Figure 6.
Gene expression heatmap constructed employing heatmapper.ca for 10 differentially expressed genes in human microglial cells challenged with A. flavus (AF) and C. albicans (CA) at 12 and 24 h p.i. The cluster value is coded with a range of colors from blue (for the lowest expression level) to red (for the highest expression level).

Figure 7.
Protein–protein interaction network for 10 target genes with a medium confidence level 0.40. Connecting line color indicates the type of information used to infer the interaction. Purple line indicates experimental evidence; blue line indicates database evidence; green line indicates gene neighborhood; red line indicates gene fusion; dark blue indicates gene co-occurrence; yellow line indicates text mining; black line indicates coexpression of genes; light blue indicates protein homology.
Figure 7.
Protein–protein interaction network for 10 target genes with a medium confidence level 0.40. Connecting line color indicates the type of information used to infer the interaction. Purple line indicates experimental evidence; blue line indicates database evidence; green line indicates gene neighborhood; red line indicates gene fusion; dark blue indicates gene co-occurrence; yellow line indicates text mining; black line indicates coexpression of genes; light blue indicates protein homology.


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Table 1.
Clinical and Demographic details of the patients diagnosed clinically with fungal endophthalmitis.
Table 1.
Clinical and Demographic details of the patients diagnosed clinically with fungal endophthalmitis.
| Demographic Characteristics | A. flavus | C. albicans |
|---|---|---|
| Age | 66 years | 54 years |
| Gender | Female | Male |
| Initial Visual Acuity | HM+ | 20/80p |
| Final Visual Acuity | HM+ | PL + PR Inaccurate |
| Microbiology | Aspergillus flavus | Candida albicans |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Gandhi, J.; Naik, P.; Kaur, I.; Kumar, A.; Joseph, J. Microglial Response to Aspergillus flavus and Candida albicans: Implications in Endophthalmitis. J. Fungi 2020, 6, 162. https://0-doi-org.brum.beds.ac.uk/10.3390/jof6030162
AMA Style
Gandhi J, Naik P, Kaur I, Kumar A, Joseph J. Microglial Response to Aspergillus flavus and Candida albicans: Implications in Endophthalmitis. Journal of Fungi. 2020; 6(3):162. https://0-doi-org.brum.beds.ac.uk/10.3390/jof6030162
Chicago/Turabian StyleGandhi, Jaishree, Poonam Naik, Inderjeet Kaur, Ashok Kumar, and Joveeta Joseph. 2020. "Microglial Response to Aspergillus flavus and Candida albicans: Implications in Endophthalmitis" Journal of Fungi 6, no. 3: 162. https://0-doi-org.brum.beds.ac.uk/10.3390/jof6030162
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.