UhAVR1, an HR-Triggering Avirulence Effector of Ustilago hordei, Is Secreted via the ER–Golgi Pathway, Localizes to the Cytosol of Barley Cells during in Planta-Expression, and Contributes to Virulence Early in Infection


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Biological Materials and Growth Conditions
2.2. Generation of Ustilago Hordei Strains
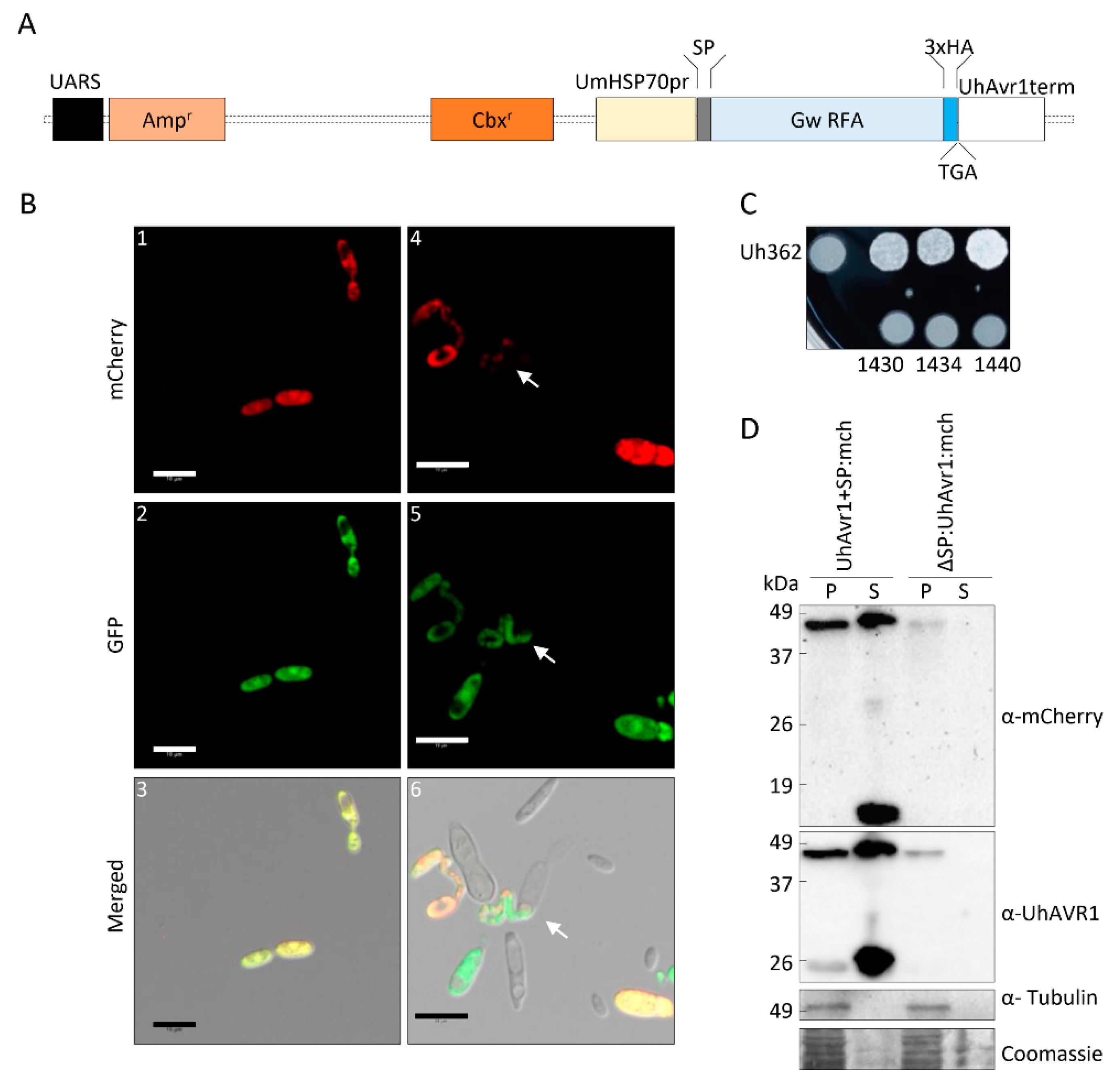
2.3. Generation of the FunGus Secretion System
2.4. Transient Expression Assays
2.5. Overexpression Viral Vector System (VOX)
2.6. N. benthamiana Cell Death Assay
2.7. Pattern-Triggered Immunity (PTI) Suppression Assay
2.8. Barley Protoplast Transfection
2.9. Brefeldin A Inhibitor Assay
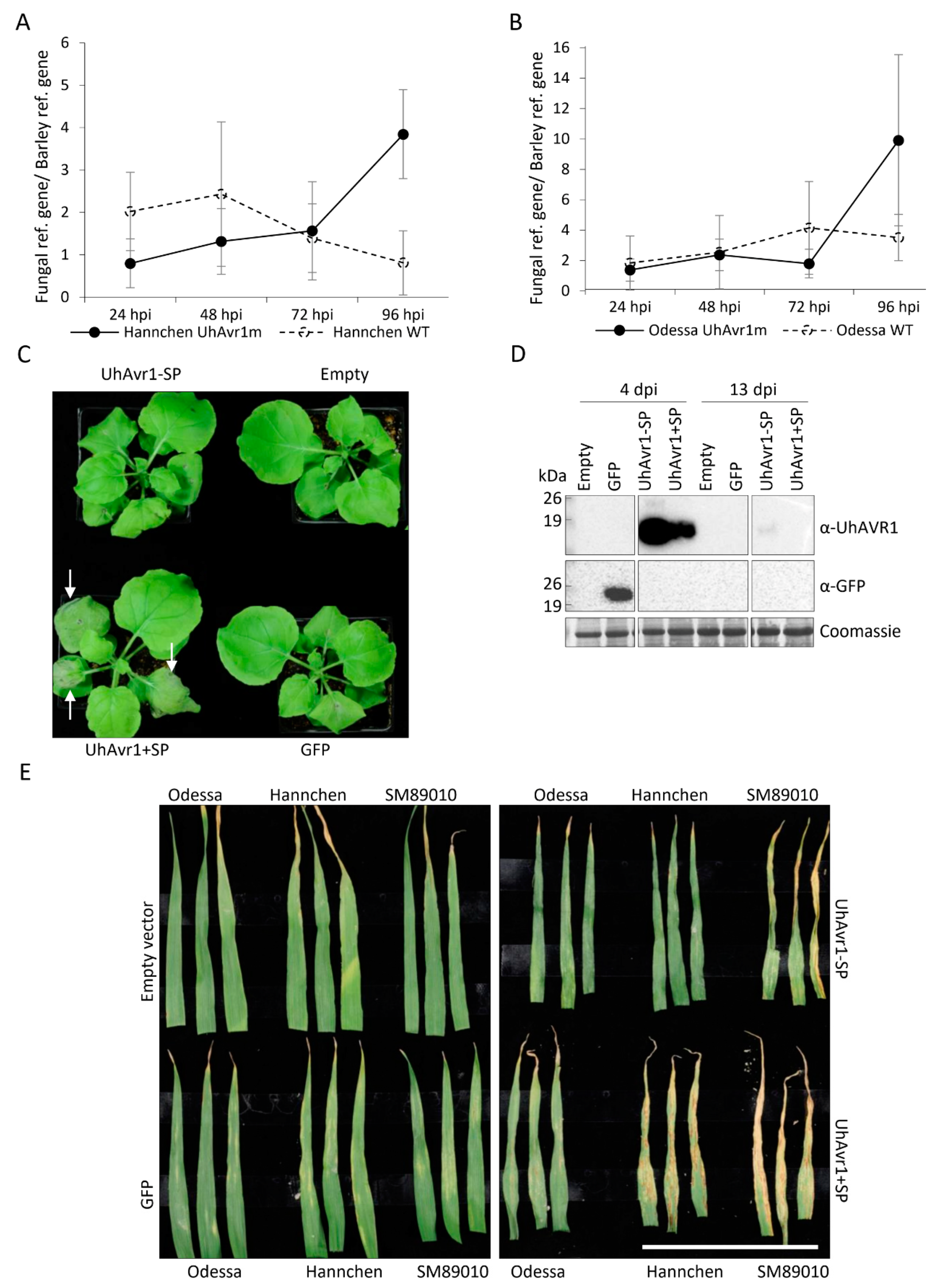
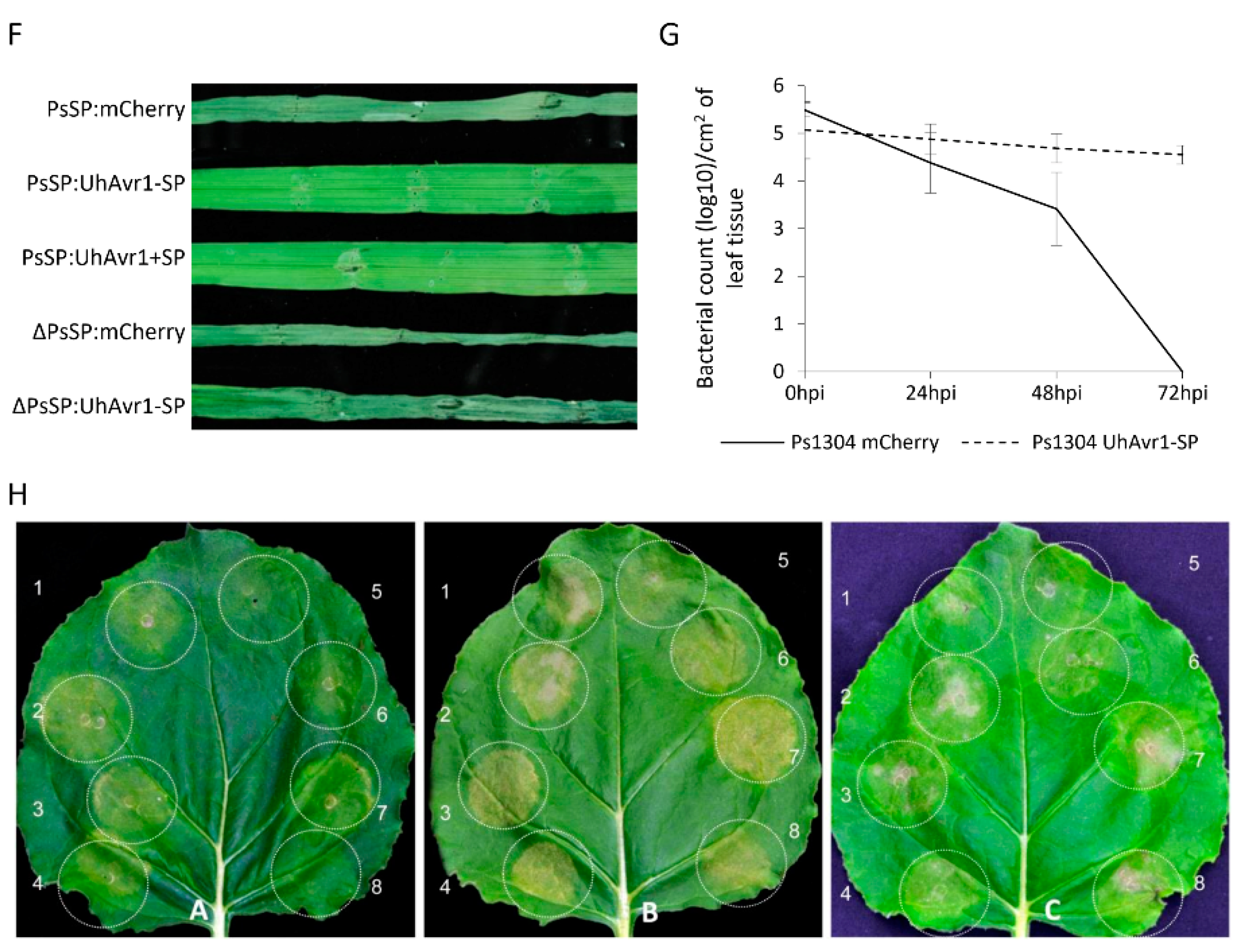
2.10. Pathogenicity Assays
2.11. RNA Extraction and Quantification
2.12. Extraction and Detection of Proteins
2.13. Staining and Microscopy
3. Results
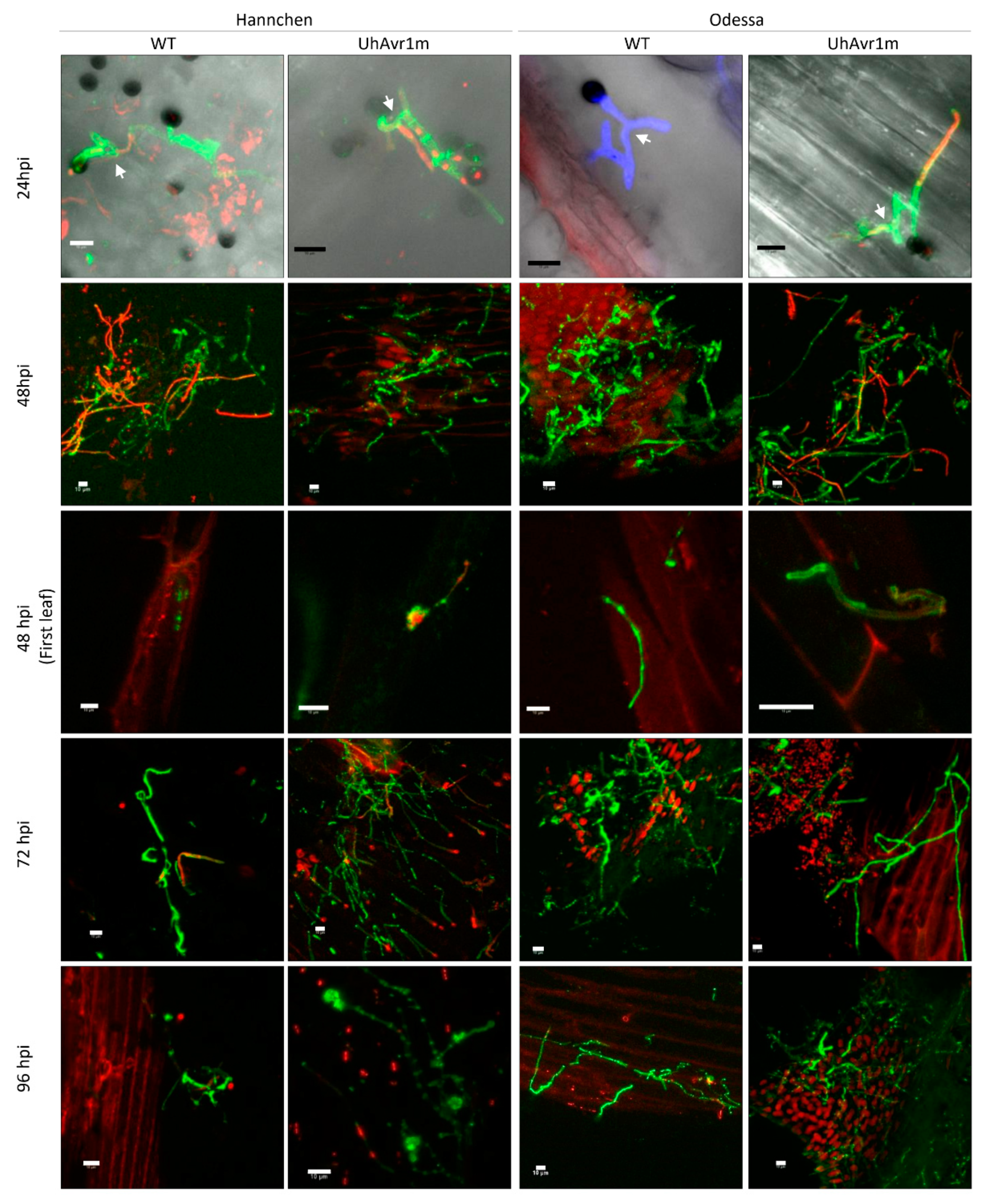
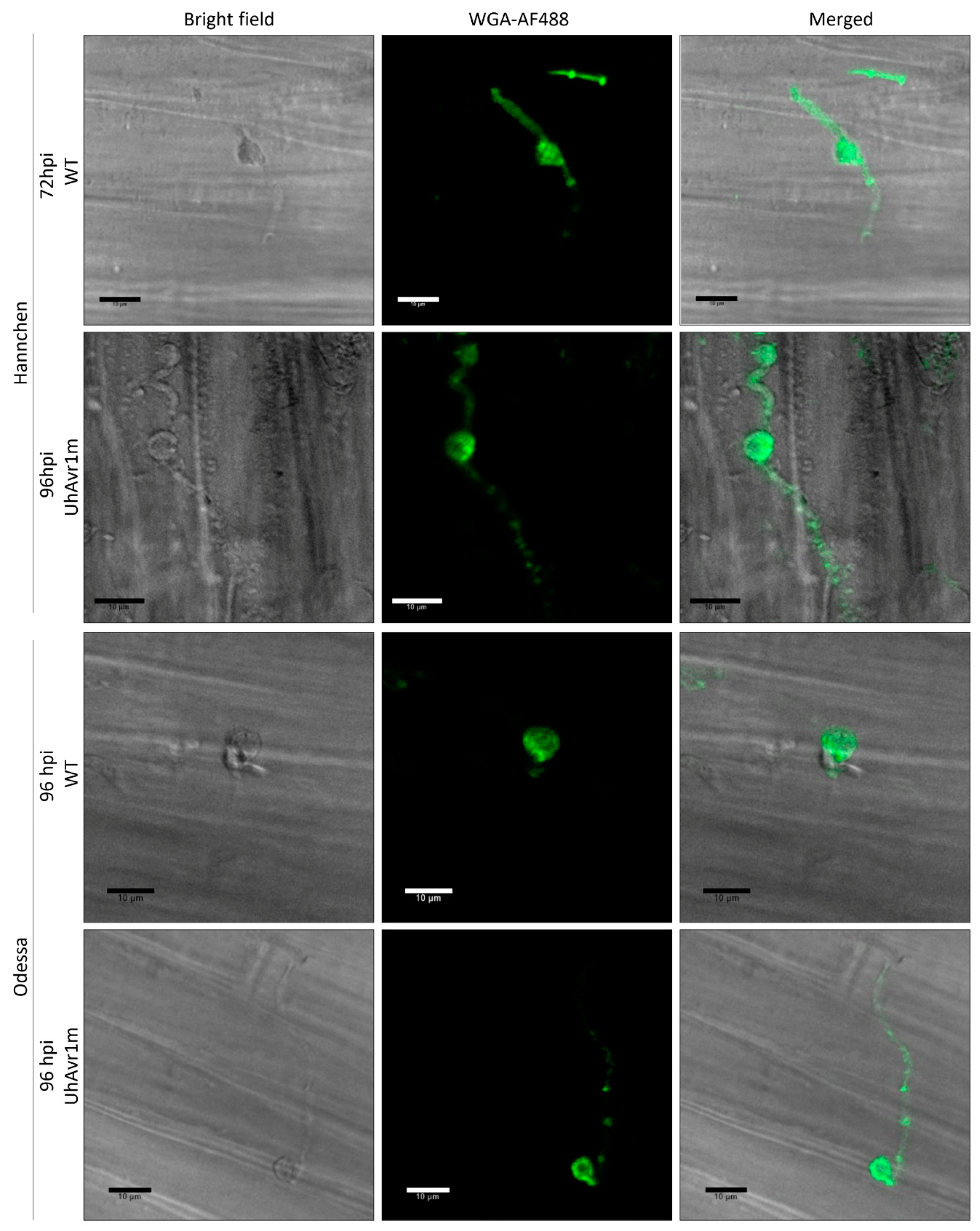
3.1. Fungal Growth Is Affected by the Deletion of UhAvr1 at the Early Stage of Barley Infection
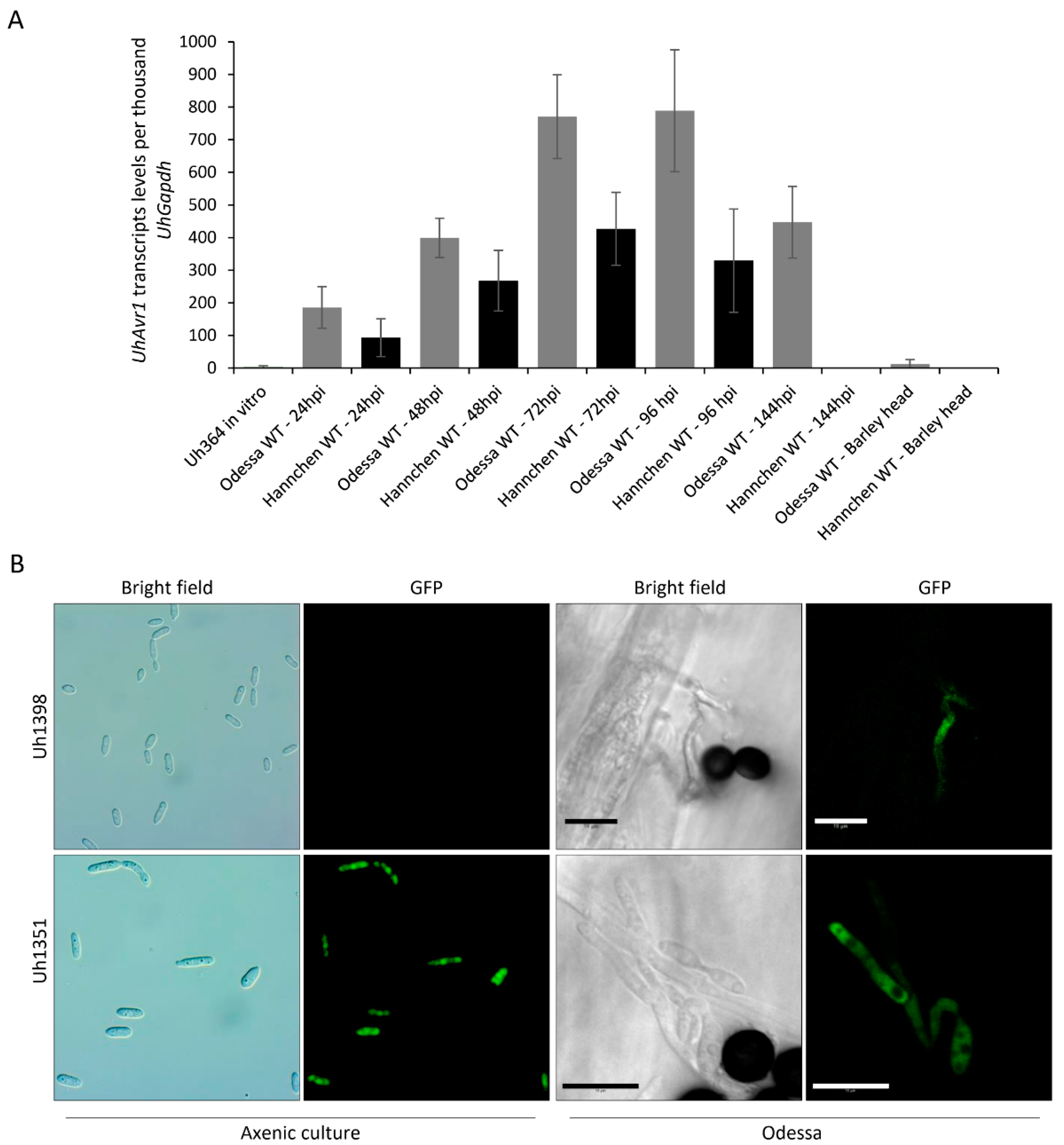
3.2. UhAvr1 Is Expressed In Planta during Early Infection of Barley Seedlings
3.3. UhAVR1 Counteracts Early Host Defence Responses and Triggers an HR in Ruh1-Harboring Barley Plants
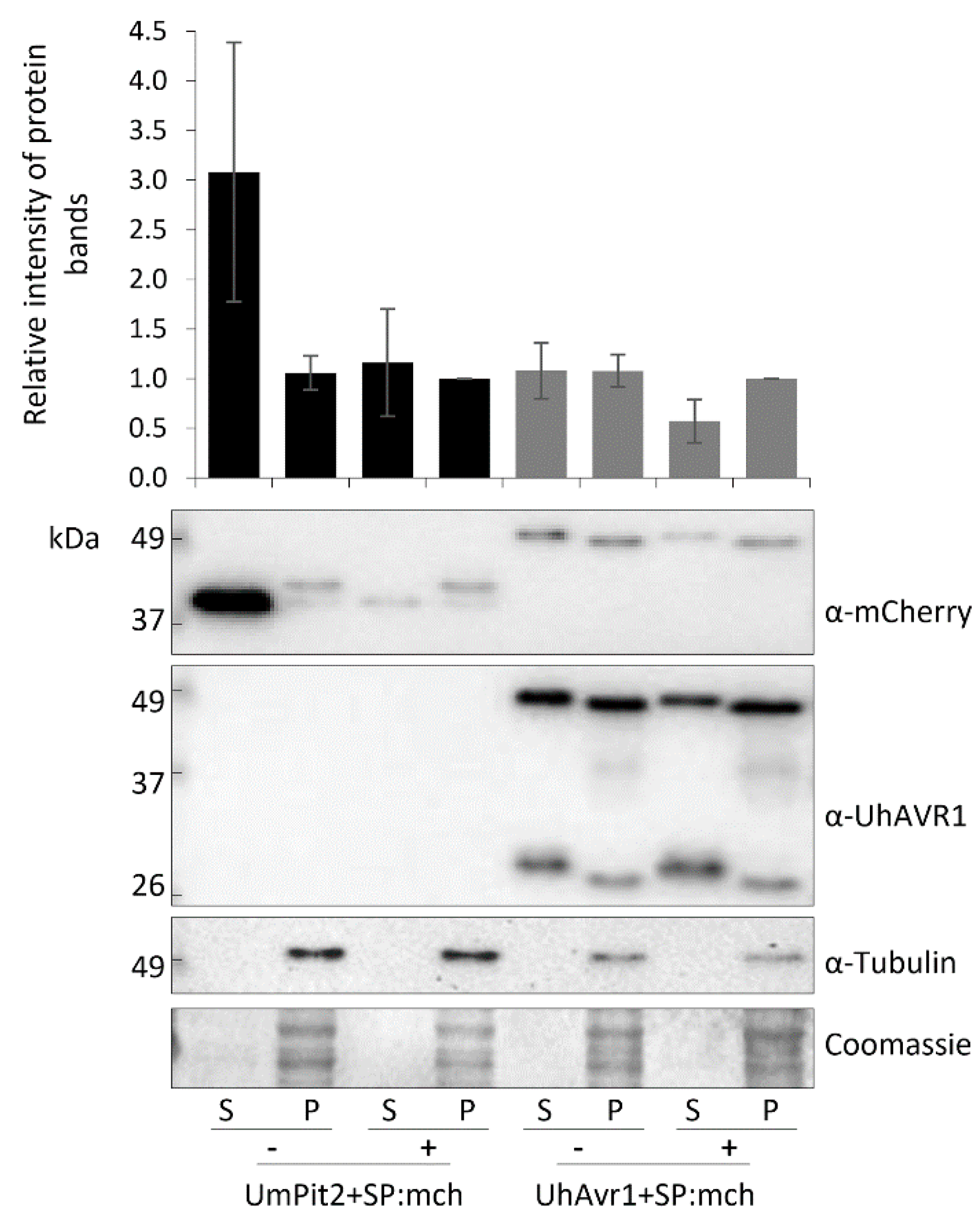
3.4. The Signal Peptide of UhAVR1 Is Essential for Secretion and Is Sensitive to Brefeldin A
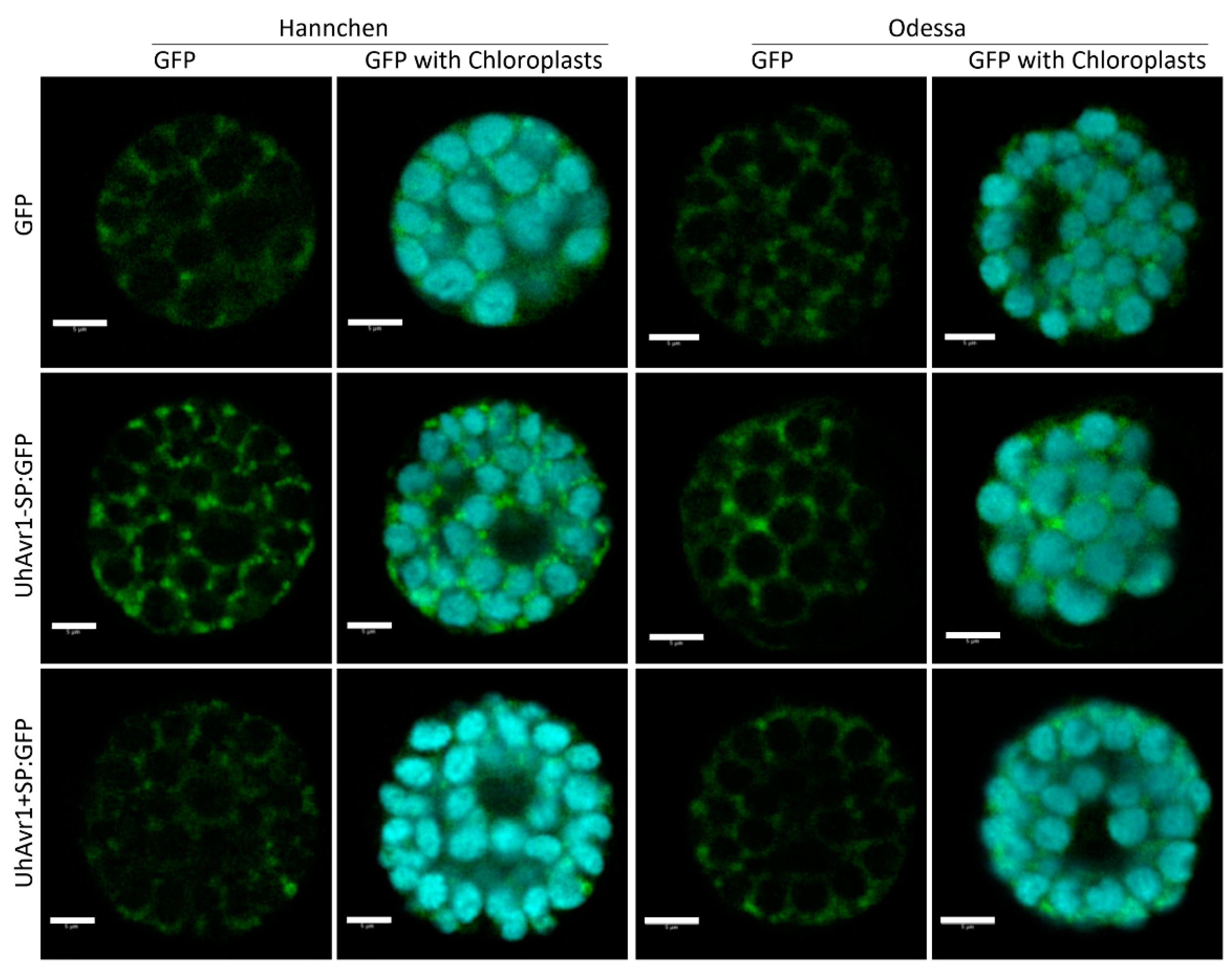
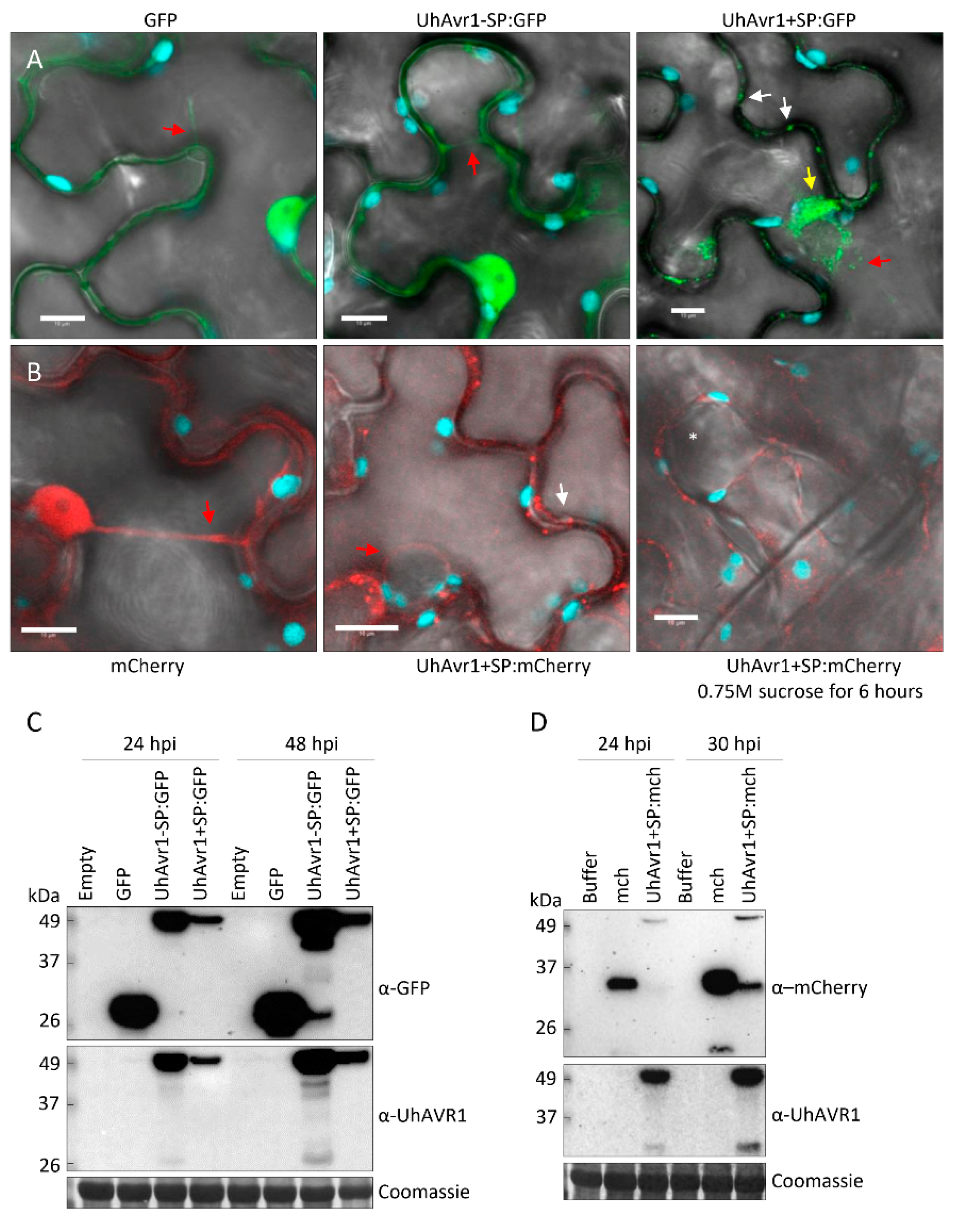
3.5. UhAVR1 Localizes to the Cytosol of Plant Cells when Delivered In Planta
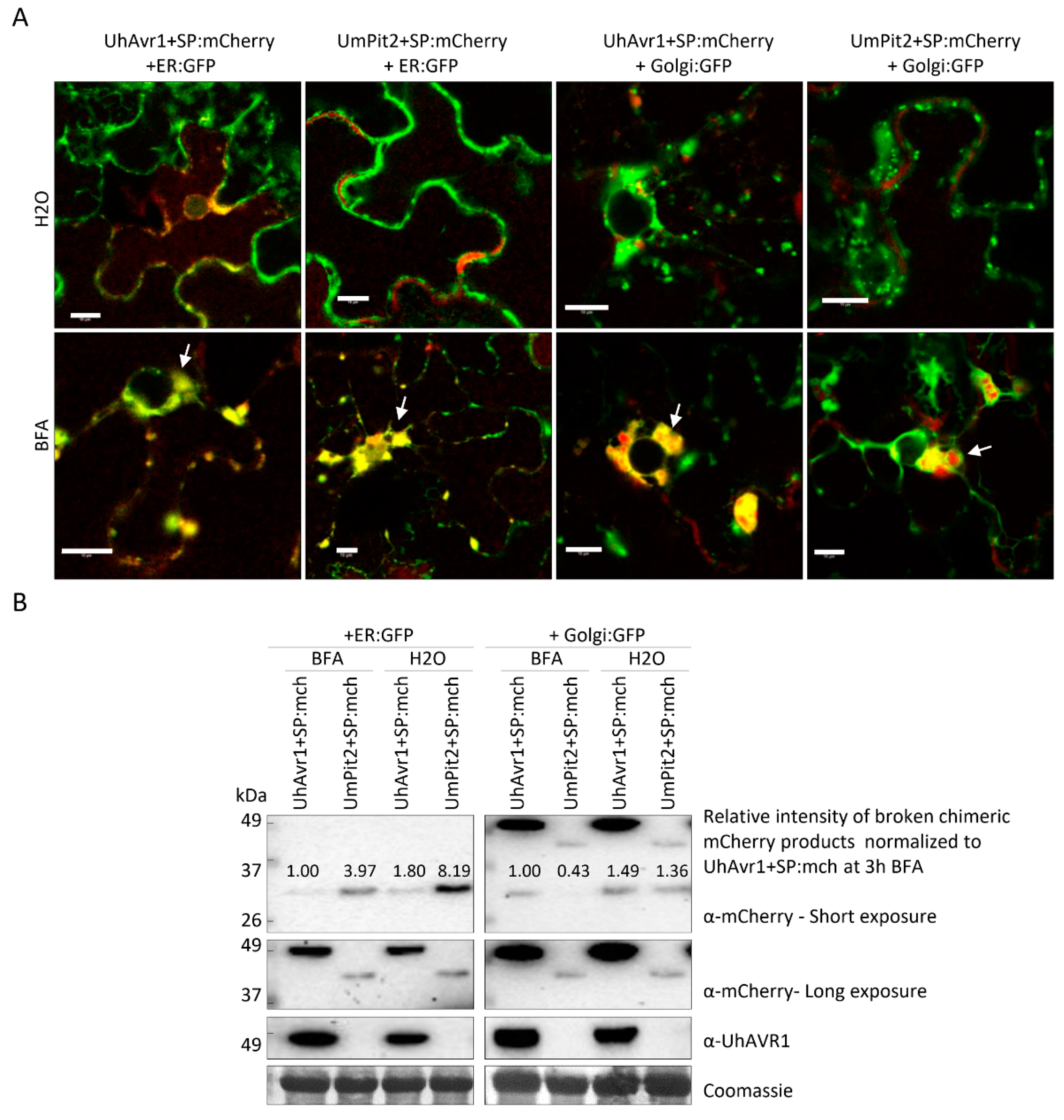
3.6. The UhAVR1 Fungal SP Directs Linked Proteins through the N. benthamiana Secretory Pathway via the ER–Golgi
4. Discussion
4.1. Expression and Function of UhAVR1
4.2. Secretion, Translocation and In Planta Localization of UhAVR1
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Bakkeren, G.; Kronstad, J.W. The pheromone cell signaling components of the Ustilago a mating-type loci determine intercompatibility between species. Genetics 1996, 143, 1601–1613. [Google Scholar] [PubMed]
- Hu, G.G.; Linning, R.; Bakkeren, G. Ultrastructural comparison of a compatible and incompatible interaction triggered by the presence of an avirulence gene during early infection of the smut fungus, Ustilago hordei, in barley. Physiol. Mol. Plant Pathol. 2003, 62, 155–166. [Google Scholar] [CrossRef]
- Hu, G.G.; Linning, R.; Bakkeren, G. Sporidial mating and infection process of the smut fungus, Ustilago hordei, in susceptible barley. Can. J. Bot. 2002, 80, 1103–1114. [Google Scholar] [CrossRef] [Green Version]
- Gaudet, D.A.; Wang, Y.; Penniket, C.; Frick, M.; Lu, Z.-X.; Bakkeren, G.; Laroche, A. Morphological and molecular analyses of host and nonhost interactions involving barley and wheat and the covered smut pathogen Ustilago hordei. Mol. Plant-Microbe Interact. 2010, 23, 1619–1634. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kozar, F. The pathway of infection of Ustilago hordei. Can. J. Genet. Cytol. 1969, 11, 977–986. [Google Scholar] [CrossRef]
- Thomas, P.L.; Metcalfe, D.R. Ustilago hordei, covered smut of barley and Ustilago nigra, false loose smut of barley. Adv. Plant Pathol. 1988, 6, 415–425. [Google Scholar]
- Linning, R.; Lin, D.; Lee, N.; Abdennadher, M.; Gaudet, D.; Thomas, P.; Mills, D.; Kronstad, J.W.; Bakkeren, G. Marker-based cloning of the region containing the UhAvr1 avirulence gene from the basidiomycete barley pathogen Ustilago hordei. Genet. 2004, 166, 99–111. [Google Scholar] [CrossRef] [Green Version]
- Laurie, J.D.; Ali, S.; Linning, R.; Mannhaupt, G.; Wong, P.; Güldener, U.; Münsterkötter, M.; Moore, R.; Kahmann, R.; Bakkeren, G.; et al. Genome comparison of barley and maize smut fungi reveals targeted loss of rna silencing components and species-specific presence of transposable elements. Plant Cell 2012, 24, 1733–1745. [Google Scholar] [CrossRef] [Green Version]
- Uhse, S.; Djamei, A. Effectors of plant-colonizing fungi and beyond. PLoS Pathog. 2018, 14, e1006992. [Google Scholar] [CrossRef] [Green Version]
- Toruño, T.Y.; Stergiopoulos, I.; Coaker, G. Plant-pathogen effectors: Cellular probes interfering with plant defenses in spatial and temporal manners. Annu. Rev. Phytopathol. 2016, 54, 419–441. [Google Scholar] [CrossRef] [Green Version]
- Lo Presti, L.; Lanver, D.; Schweizer, G.; Tanaka, S.; Liang, L.; Tollot, M.; Zuccaro, A.; Reissmann, S.; Kahmann, R. Fungal effectors and plant susceptibility. Annu. Rev. Plant Biol. 2015, 66, 513–545. [Google Scholar] [CrossRef] [PubMed]
- Selin, C.; de Kievit, T.R.; Belmonte, M.F.; Fernando, W.G.D. Elucidating the role of effectors in plant-fungal interactions: Progress and challenges. Front. Microbiol. 2016, 7, 600. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sidhu, G.; Person, C. Genetic control of virulence in Ustilago hordei. I. Segregations for higher levels of virulence. Can. J. Genet. Cytol. 1971, 13, 173–178. [Google Scholar] [CrossRef]
- Ebba, T.; Person, C. Genetic control of virulence in Ustilago hordei. IV. Duplicate genes for virulence and genetic and environmental modification of a gene-for-gene relationship. Can. J. Genet. Cytol. 1975, 17, 631–636. [Google Scholar] [CrossRef]
- Thomas, P.L. Interaction of virulence genes in Ustilago hordei. Can. J. Genet. Cytol. 1976, 18, 141–149. [Google Scholar] [CrossRef]
- Ali, S.; Laurie, J.D.; Linning, R.; Cervantes-Chávez, J.A.; Gaudet, D.; Bakkeren, G. An immunity-triggering effector from the barley smut fungus Ustilago hordei resides in an Ustilaginaceae-specific cluster bearing signs of transposable element-assisted evolution. PLoS Pathog. 2014, 10, e1004223. [Google Scholar] [CrossRef]
- Eitzen, K.; Sengupta, P.; Kroll, S.; Kemen, E.; Doehlemann, G. An antagonistic driver of the microbial phyllosphere suppresses infection of Arabidopsis thaliana by the oomycete pathogen Albugo laibachii via a secreted hydrolase. bioRxiv 2020. [Google Scholar] [CrossRef] [Green Version]
- Ökmen, B.; Mathow, D.; Hof, A.; Lahrmann, U.; Aßmann, D.; Doehlemann, G. Mining the effector repertoire of the biotrophic fungal pathogen Ustilago hordei during host and non-host infection. Mol. Plant Pathol. 2018, 19, 2603–2622. [Google Scholar] [CrossRef] [Green Version]
- Sidhu, G.; Person, C. Genetic control of virulence in Ustilago hordei. II. Identification of genes for host resistance and demonstration of gene-for-gene relations. Can. J. Genet. Cytol. 1972, 14, 209–213. [Google Scholar] [CrossRef]
- Grewal, T.S.; Rossnagel, B.G.; Bakkeren, G.; Scoles, G.J. Identification of resistance genes to barley covered smut and mapping of the Ruh1 gene using Ustilago hordei strains with defined avirulence genes. Can. J. Plant Pathol. 2008, 30, 277–284. [Google Scholar] [CrossRef]
- Bouton, C.; King, R.C.; Chen, H.; Azhakanandam, K.; Bieri, S.; Hammond-Kosack, K.E.; Kanyuka, K. Foxtail mosaic virus: A viral vector for protein expression in cereals. Plant Physiol. 2018, 177, 1352–1367. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Holliday, R. The genetics of Ustilago maydis. Genet. Res. Camb. 1961, 2, 204–230. [Google Scholar] [CrossRef]
- Takahashi, Y.; Omura, T.; Hibino, H.; Sato, M. Detection and identification of Pseudomonas syringae pv. atropurpurea by PCR amplification of specific fragments from an indigenous plasmid. Plant Dis. 1996, 80, 783–788. [Google Scholar] [CrossRef]
- Hu, G.; Kamp, A.; Linning, R.; Naik, S.; Bakkeren, G. Complementation of Ustilago maydis MAPK mutants by a wheat leaf rust, Puccinia triticina homolog: Potential for functional analyses of rust genes. Mol. Plant-Microbe Interact. 2007, 20, 637–647. [Google Scholar] [CrossRef] [Green Version]
- García-Pedrajas, M.D.; Nadal, M.; Kapa, L.B.; Perlin, M.H.; Andrews, D.L.; Gold, S.E. DelsGate, a robust and rapid gene deletion construction method. Fungal Genet. Biol. 2008, 45, 379–388. [Google Scholar] [CrossRef]
- Nielsen, H. Predicting secretory proteins with SignalP. In Protein Function Prediction: Methods and Protocols; Kihara, D., Ed.; Springer: New York, NY, USA, 2017; pp. 59–73. [Google Scholar]
- Karimi, M.; Inzé, D.; Depicker, A. GATEWAY™ vectors for Agrobacterium-mediated plant transformation. Trends Plant Sci. 2002, 7, 193–195. [Google Scholar] [CrossRef]
- Redkar, A.; Doehlemann, G. Ustilago maydis virulence assays in maize. Bio-Protocol 2016, 6, e1760. [Google Scholar] [CrossRef]
- van Engelen, F.A.; Molthoff, J.W.; Conner, A.J.; Nap, J.-P.; Pereira, A.; Stiekema, W.J. pBINPLUS: An improved plant transformation vector based on pBIN19. Transgenic Res. 1995, 4, 288–290. [Google Scholar] [CrossRef]
- Beauchemin, C.; Laliberté, J.-F. The poly(A) binding protein is internalized in virus-induced vesicles or redistributed to the nucleolus during turnip mosaic virus infection. J. Virol. 2007, 81, 10905–10913. [Google Scholar] [CrossRef] [Green Version]
- Nelson, B.K.; Cai, X.; Nebenführ, A. A multicolored set of in vivo organelle markers for co-localization studies in Arabidopsis and other plants. Plant J. 2007, 51, 1126–1136. [Google Scholar] [CrossRef]
- Ali, S.; Magne, M.; Chen, S.; Côté, O.; Stare, B.G.; Obradovic, N.; Jamshaid, L.; Wang, X.; Bélair, G.; Moffett, P. Analysis of putative apoplastic effectors from the nematode, Globodera rostochiensis, and identification of an expansin-like protein that can induce and suppress host defenses. PLoS ONE 2015, 10, e0115042. [Google Scholar] [CrossRef] [PubMed]
- Wang, K.-D.; Empleo, R.; Nguyen, T.T.V.; Moffett, P.; Sacco, M.A. Elicitation of hypersensitive responses in Nicotiana glutinosa by the suppressor of RNA silencing protein P0 from poleroviruses. Mol. Plant Pathol. 2015, 16, 435–448. [Google Scholar] [CrossRef] [PubMed]
- Kamoun, S.; van West, P.; Vleeshouwers, V.G.; de Groot, K.E.; Govers, F. Resistance of Nicotiana benthamiana to Phytophthora infestans is mediated by the recognition of the elicitor protein INF1. Plant Cell 1998, 10, 1413–1426. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bendahmane, A.; Farnham, G.; Moffett, P.; Baulcombe, D.C. Constitutive gain-of-function mutants in a nucleotide binding site-leucine rich repeat protein encoded at the Rx locus of potato. Plant J. 2002, 32, 195–204. [Google Scholar] [CrossRef] [Green Version]
- Rairdan, G.J.; Moffett, P. Distinct domains in the ARC region of the potato resistance protein Rx mediate LRR binding and inhibition of activation. Plant Cell 2006, 18, 2082–2093. [Google Scholar] [CrossRef] [Green Version]
- Tai, T.H.; Dahlbeck, D.; Clark, E.T.; Gajiwala, P.; Pasion, R.; Whalen, M.C.; Stall, R.E.; Staskawicz, B.J. Expression of the Bs2 pepper gene confers resistance to bacterial spot disease in tomato. Proc. Natl. Acad. Sci. USA 1999, 96, 14153–14158. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rentel, M.C.; Leonelli, L.; Dahlbeck, D.; Zhao, B.; Staskawicz, B.J. Recognition of the Hyaloperonospora parasitica effector ATR13 triggers resistance against oomycete, bacterial, and viral pathogens. Proc. Natl. Acad. Sci. USA 2008, 105, 1091–1096. [Google Scholar] [CrossRef] [Green Version]
- Choi, K.H.; Kumar, A.; Schweizer, H.P. A 10-min method for preparation of highly electrocompetent Pseudomonas aeruginosa cells: Application for DNA fragment transfer between chromosomes and plasmid transformation. J. Microbiol. Methods 2006, 64, 391–397. [Google Scholar] [CrossRef]
- Katagiri, F.; Thilmony, R.; He, S.Y. The Arabidopsis thaliana-Pseudomonas syringae interaction. Am. Soc. Plant Biol. 2002, 1, e0039. [Google Scholar] [CrossRef] [Green Version]
- Nebenführ, A.; Ritzenthaler, C.; Robinson, D.G. Brefeldin A: Deciphering an enigmatic inhibitor of secretion. Plant Physiol. 2002, 130, 1102–1108. [Google Scholar] [CrossRef] [Green Version]
- Liu, D.; Shi, L.; Han, C.; Yu, J.; Li, D.; Zhang, Y. Validation of reference genes for gene expression studies in virus-infected Nicotiana benthamiana using quantitative real-time PCR. PLoS ONE 2012, 7, e46451. [Google Scholar] [CrossRef] [PubMed]
- Hua, W.; Zhu, J.; Shang, Y.; Wang, J.; Jia, Q.; Yang, J. Identification of suitable reference genes for barley gene expression under abiotic stresses and hormonal treatments. Plant Mol. Biol. Rep. 2015, 33, 1002–1012. [Google Scholar] [CrossRef]
- Muller, O.; Schreier, P.H.; Uhrig, J.F. Identification and characterization of secreted and pathogenesis-related proteins in Ustilago maydis. Mol. Gen. Genom. 2008, 279, 27–39. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Redkar, A.; Jaeger, E.; Doehlemann, G. Visualization of growth and morphology of fungal hyphae in planta using WGA-AF488 and Propidium Iodide co-staining. Bio-Protocol 2018, 8, e2942. [Google Scholar] [CrossRef]
- Panwar, V.; McCallum, B.; Bakkeren, G. Host-induced gene silencing of wheat leaf rust fungus Puccinia triticina pathogenicity genes mediated by the Barley stripe mosaic virus. Plant Mol. Biol. 2013, 81, 595–608. [Google Scholar] [CrossRef]
- Deising, H.B.; Werner, S.; Wernitz, M. The role of fungal appressoria in plant infection. Microbes Infect. 2000, 2, 1631–1641. [Google Scholar] [CrossRef]
- Ryder, L.S.; Talbot, N.J. Regulation of appressorium development in pathogenic fungi. Curr. Opin. Plant Biol. 2015, 26, 8–13. [Google Scholar] [CrossRef] [Green Version]
- Snetselaar, K.M.; Mims, C.W. Sporidial fusion and infection of maize seedlings by the smut fungus Ustilago maydis. Mycologia 1992, 84, 193–203. [Google Scholar] [CrossRef]
- Snetselaar, K.M.; Mims, C.W. Infection of maize stigmas by Ustilago maydis: Light and electron microscopy. Phytopathology 1993, 83, 843–850. [Google Scholar] [CrossRef]
- Kamper, J.; Kahmann, R.; Bolker, M.; Ma, L.J.; Brefort, T.; Saville, B.J.; Banuett, F.; Kronstad, J.W.; Gold, S.E.; Muller, O.; et al. Insights from the genome of the biotrophic fungal plant pathogen Ustilago maydis. Nature 2006, 444, 97–101. [Google Scholar] [CrossRef]
- Kanzaki, H.; Saitoh, H.; Takahashi, Y.; Berberich, T.; Ito, A.; Kamoun, S.; Terauchi, R. NbLRK1, a lectin-like receptor kinase protein of Nicotiana benthamiana, interacts with Phytophthora infestans INF1 elicitin and mediates INF1-induced cell death. Planta 2008, 228, 977–987. [Google Scholar] [CrossRef]
- Slootweg, E.; Roosien, J.; Spiridon, L.N.; Petrescu, A.-J.; Tameling, W.; Joosten, M.; Pomp, R.; van Schaik, C.; Dees, R.; Borst, J.W.; et al. Nucleocytoplasmic distribution is required for activation of resistance by the potato NB-LRR receptor Rx1 and is balanced by its functional domains. Plant Cell 2010, 22, 4195–4215. [Google Scholar] [CrossRef] [Green Version]
- Mudgett, M.B.; Chesnokova, O.; Dahlbeck, D.; Clark, E.T.; Rossier, O.; Bonas, U.; Staskawicz, B.J. Molecular signals required for type III secretion and translocation of the Xanthomonas campestris AvrBs2 protein to pepper plants. Proc. Natl. Acad. Sci. USA 2000, 97, 13324–13329. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, S.; Boevink, P.C.; Welsh, L.; Zhang, R.; Whisson, S.C.; Birch, P.R.J. Delivery of cytoplasmic and apoplastic effectors from Phytophthora infestans haustoria by distinct secretion pathways. New Phytol. 2017, 216, 205–215. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Doehlemann, G.; Reissmann, S.; Aßmann, D.; Fleckenstein, M.; Kahmann, R. Two linked genes encoding a secreted effector and a membrane protein are essential for Ustilago maydis-induced tumour formation. Mol. Microbiol. 2011, 81, 751–766. [Google Scholar] [CrossRef] [PubMed]
- Mueller, A.N.; Ziemann, S.; Treitschke, S.; Assmann, D.; Doehlemann, G. Compatibility in the Ustilago maydis-maize interaction requires inhibition of host cysteine proteases by the fungal effector Pit2. PLoS Pathog. 2013, 9, e1003177. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tanaka, S.; Djamei, A.; Presti, L.L.; Schipper, K.; Winterberg, S.; Amati, S.; Becker, D.; Büchner, H.; Kumlehn, J.; Reissmann, S.; et al. Experimental approaches to investigate effector translocation into host cells in the Ustilago maydis/maize pathosystem. Eur. J. Cell Biol. 2015, 94, 349–358. [Google Scholar] [CrossRef]
- Boevink, P.; Martin, B.; Oparka, K.; Santa Cruz, S.; Hawes, C. Transport of virally expressed green fluorescent protein through the secretory pathway in tobacco leaves is inhibited by cold shock and brefeldin A. Planta 1999, 208, 392–400. [Google Scholar] [CrossRef]
- Zuo, W.; Okmen, B.; Depotter, J.R.L.; Ebert, M.K.; Redkar, A.; Villamil, J.M.; Doehlemann, G. Molecular interactions between smut fungi and their host plants. Annu. Rev. Phytopathol. 2019, 57, 411–430. [Google Scholar] [CrossRef]
- Bhattacharjee, S.; Halane, M.K.; Kim, S.H.; Gassmann, W. Pathogen effectors target Arabidopsis EDS1 and alter its interactions with immune regulators. Science 2011, 334, 1405–1408. [Google Scholar] [CrossRef]
- Brefort, T.; Tanaka, S.; Neidig, N.; Doehlemann, G.; Vincon, V.; Kahmann, R. Characterization of the largest effector gene cluster of Ustilago maydis. PLoS Pathog. 2014, 10, e1003866. [Google Scholar] [CrossRef] [Green Version]
- Abramovitch, R.B.; Kim, Y.J.; Chen, S.; Dickman, M.B.; Martin, G.B. Pseudomonas type III effector AvrPtoB induces plant disease susceptibility by inhibition of host programmed cell death. EMBO J. 2003, 22, 60–69. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bos, J.I.B.; Kanneganti, T.-D.; Young, C.; Cakir, C.; Huitema, E.; Win, J.; Armstrong, M.R.; Birch, P.R.J.; Kamoun, S. The C-terminal half of Phytophthora infestans RXLR effector AVR3a is sufficient to trigger R3a-mediated hypersensitivity and suppress INF1-induced cell death in Nicotiana benthamiana. Plant J. 2006, 48, 165–176. [Google Scholar] [CrossRef] [PubMed]
- Yoshino, K.; Irieda, H.; Sugimoto, F.; Yoshioka, H.; Okuno, T.; Takano, Y. Cell death of Nicotiana benthamiana is induced by secreted protein NIS1 of Colletotrichum orbiculare and is suppressed by a homologue of CgDN3. Mol. Plant-Microbe Int. 2012, 25, 625–636. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ramachandran, S.R.; Yin, C.; Kud, J.; Tanaka, K.; Mahoney, A.K.; Xiao, F.; Hulbert, S.H. Effectors from wheat rust fungi suppress multiple plant defense responses. Phytopathology 2016, 107, 75–83. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dou, D.; Kale, S.D.; Wang, X.; Chen, Y.; Wang, Q.; Wang, X.; Jiang, R.H.; Arredondo, F.D.; Anderson, R.G.; Thakur, P.B.; et al. Conserved C-terminal motifs required for avirulence and suppression of cell death by Phytophthora sojae effector Avr1b. Plant Cell 2008, 20, 1118–1133. [Google Scholar] [CrossRef] [Green Version]
- Wharton, P.S.; Julian, A.M.; O’Connell, R.J. Ultrastructure of the infection of Sorghum bicolor by Colletotrichum sublineolum. Phytopathology 2001, 91, 149–158. [Google Scholar] [CrossRef] [Green Version]
- Carisse, K.X.; Burnett, P.A.; Tewari, J.P.; Chen, M.H.; Turkington, T.K.; Helm, J.H. Histopathological study of barley cultivars resistant and susceptible to Rhynchosporium secalis. Phytopathology 2000, 90, 94–102. [Google Scholar] [CrossRef]
- Ma, Q.; Shang, H.S. Ultrastructure of stripe rust (Puccinia striiformis f. sp. tritici) interacting with slow-rusting, highly resistant, and susceptible wheat cultivars. J. Plant Pathol. 2009, 91, 597–606. [Google Scholar]
- Kang, Z.; Huang, L.; Buchenauer, H. Ultrastructural changes and localization of lignin and callose in compatible and incompatible interactions between wheat and Puccinia striiformis. J. Plant Dis. Prot. 2002, 109, 25–37. [Google Scholar]
- Iwamoto, M.; Takeuchi, Y.; Takada, Y.; Yamaoka, N. Coleoptile surface cuticle of barley is involved in survival and penetration of Blumeria graminis. Physiol. Mol. Plant Pathol. 2002, 60, 31–38. [Google Scholar] [CrossRef]
- Dalio, R.J.D.; Herlihy, J.; Oliveira, T.S.; McDowell, J.M.; Machado, M. Effector biology in focus: A primer for computational prediction and functional characterization. Mol. Plant-Microbe Interact. 2018, 31, 22–33. [Google Scholar] [CrossRef] [PubMed]
- Viotti, C. ER to Golgi-dependent protein secretion: The conventional pathway. In Unconventional Protein Secretion: Methods and Protocols; Pompa, A., De Marchis, F., Eds.; Springer: New York, NY, USA, 2016; pp. 3–29. [Google Scholar]
- Liu, T.; Song, T.; Zhang, X.; Yuan, H.; Su, L.; Li, W.; Xu, J.; Liu, S.; Chen, L.; Chen, T.; et al. Unconventionally secreted effectors of two filamentous pathogens target plant salicylate biosynthesis. Nat. Commun. 2014, 5, 4686. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Giraldo, M.C.; Dagdas, Y.F.; Gupta, Y.K.; Mentlak, T.A.; Yi, M.; Martinez-Rocha, A.L.; Saitoh, H.; Terauchi, R.; Talbot, N.J.; Valent, B. Two distinct secretion systems facilitate tissue invasion by the rice blast fungus Magnaporthe oryzae. Nat. Commun. 2013, 4, 1996. [Google Scholar] [CrossRef] [Green Version]
- Welsh, L.; Thorpe, P.; Whisson, S.C.; Boevink, P.C.; Birch, P.R.J. The Phytophthora infestans haustorium is a site for secretion of diverse classes of infection-associated proteins. mBio 2018, 9, e01216–e01218. [Google Scholar]
- Dutra, D.; Agrawal, N.; Ghareeb, H.; Schirawski, J. Screening of secreted proteins of Sporisorium reilianum f. sp. zeae for cell death suppression in Nicotiana benthamiana. Front. Plant Sci. 2020, 11, 95. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Christie, M.; Chang, C.-W.; Róna, G.; Smith, K.M.; Stewart, A.G.; Takeda, A.A.S.; Fontes, M.R.M.; Stewart, M.; Vértessy, B.G.; Forwood, J.K.; et al. Structural biology and regulation of protein import into the nucleus. J. Mol. Biol. 2016, 428, 2060–2090. [Google Scholar] [CrossRef] [PubMed]
- Brandizzi, F.; Hanton, S.; DaSilva, L.L.P.; Boevink, P.; Evans, D.; Oparka, K.; Denecke, J.; Hawes, C. ER quality control can lead to retrograde transport from the ER lumen to the cytosol and the nucleoplasm in plants. Plant J. 2003, 34, 269–281. [Google Scholar] [CrossRef] [Green Version]
- Tintor, N.; Paauw, M.; Rep, M.; Takken, F.L.W. The root-invading pathogen Fusarium oxysporum targets pattern-triggered immunity using both cytoplasmic and apoplastic effectors. New Phytol. 2020, 227, 1479–1492. [Google Scholar] [CrossRef] [Green Version]
- Manning, V.A.; Ciuffetti, L.M. Localization of Ptr ToxA produced by Pyrenophora tritici-repentis reveals protein import into wheat mesophyll cells. Plant Cell 2005, 17, 3203–3212. [Google Scholar] [CrossRef] [Green Version]
- Rafiqi, M.; Gan, P.H.P.; Ravensdale, M.; Lawrence, G.J.; Ellis, J.G.; Jones, D.A.; Hardham, A.R.; Dodds, P.N. Internalization of flax rust avirulence proteins into flax and tobacco cells can occur in the absence of the pathogen. Plant Cell 2010, 22, 2017–2032. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Catanzariti, A.M.; Dodds, P.N.; Lawrence, G.J.; Ayliffe, M.A.; Ellis, J.G. Haustorially-expressed secreted proteins from flax rust are highly enriched for avirulence elicitors. Plant Cell 2006, 18, 243–256. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gu, B.; Kale, S.D.; Wang, Q.; Wang, D.; Pan, Q.; Cao, H.; Meng, Y.; Kang, Z.; Tyler, B.M.; Shan, W. Rust secreted protein Ps87 is conserved in diverse fungal pathogens and contains a RXLR-like motif sufficient for translocation into plant cells. PLoS ONE 2011, 6, e27217. [Google Scholar] [CrossRef] [PubMed]
- Di, X.; Gomila, J.; Ma, L.; van den Burg, H.A.; Takken, F.L.W. Uptake of the Fusarium effector Avr2 by tomato is not a cell autonomous event. Front. Plant Sci. 2016, 7, 1915. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ve, T.; Williams, S.J.; Catanzariti, A.M.; Rafiqi, M.; Rahman, M.; Ellis, J.G.; Hardham, A.R.; Jones, D.A.; Anderson, P.A.; Dodds, P.N.; et al. Structures of the flax-rust effector AvrM reveal insights into the molecular basis of plant cell entry and effector-triggered immunity. Proc. Natl. Acad. Sci. USA 2013, 110, 17594–17599. [Google Scholar] [CrossRef] [Green Version]










© 2020 by Her Majesty the Queen in Right of Canada, as represented by the Minister of Agriculture and Agri-Food Canada. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Montenegro Alonso, A.P.; Ali, S.; Song, X.; Linning, R.; Bakkeren, G. UhAVR1, an HR-Triggering Avirulence Effector of Ustilago hordei, Is Secreted via the ER–Golgi Pathway, Localizes to the Cytosol of Barley Cells during in Planta-Expression, and Contributes to Virulence Early in Infection. J. Fungi 2020, 6, 178. https://0-doi-org.brum.beds.ac.uk/10.3390/jof6030178
Montenegro Alonso AP, Ali S, Song X, Linning R, Bakkeren G. UhAVR1, an HR-Triggering Avirulence Effector of Ustilago hordei, Is Secreted via the ER–Golgi Pathway, Localizes to the Cytosol of Barley Cells during in Planta-Expression, and Contributes to Virulence Early in Infection. Journal of Fungi. 2020; 6(3):178. https://0-doi-org.brum.beds.ac.uk/10.3390/jof6030178
Chicago/Turabian StyleMontenegro Alonso, Ana Priscilla, Shawkat Ali, Xiao Song, Rob Linning, and Guus Bakkeren. 2020. "UhAVR1, an HR-Triggering Avirulence Effector of Ustilago hordei, Is Secreted via the ER–Golgi Pathway, Localizes to the Cytosol of Barley Cells during in Planta-Expression, and Contributes to Virulence Early in Infection" Journal of Fungi 6, no. 3: 178. https://0-doi-org.brum.beds.ac.uk/10.3390/jof6030178