
Potent and Selective Inhibitors of Human Monoamine Oxidase A from an Endogenous Lichen Fungus Diaporthe mahothocarpus

 , ,
, ,  and
and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Evaluation of Enzyme Inhibitory Activities of ELF Extracts
2.2. MAO Activity Assay
2.3. ChE Activity Assay
2.4. BACE1 Activity Assay
2.5. Culture and Extraction of ELF29
2.6. Isolation of Compounds from ELF29 Crude Extract
2.7. Structure Analysis of the Compounds through NMR and LC/MS
2.8. Analysis of Inhibitory Activities and Kinetics of the Compounds
2.9. Analysis of Inhibitor Reversibility
2.10. Docking Simulations of the Compounds with hMAO-A and hMAO-B
2.11. Analysis of Pharmacokinetic and Physicochemical Parameters of the Compounds Using in Silico Method
3. Results
3.1. Inhibitory Activities of ELF Extracts against hMAO-A, hMAO-B, AChE, and BChE
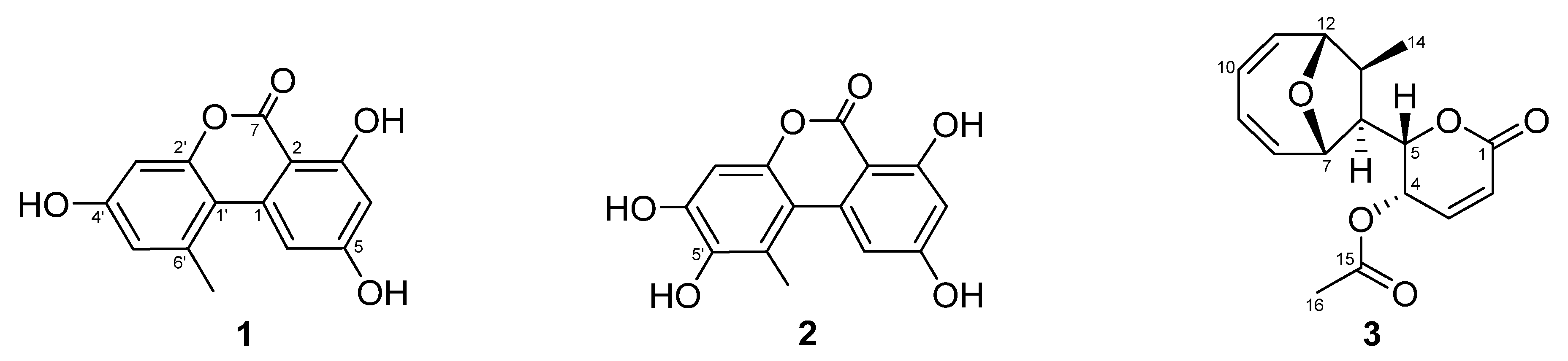
3.2. Isolation and Identification of the Compounds from the ELF29 Extract
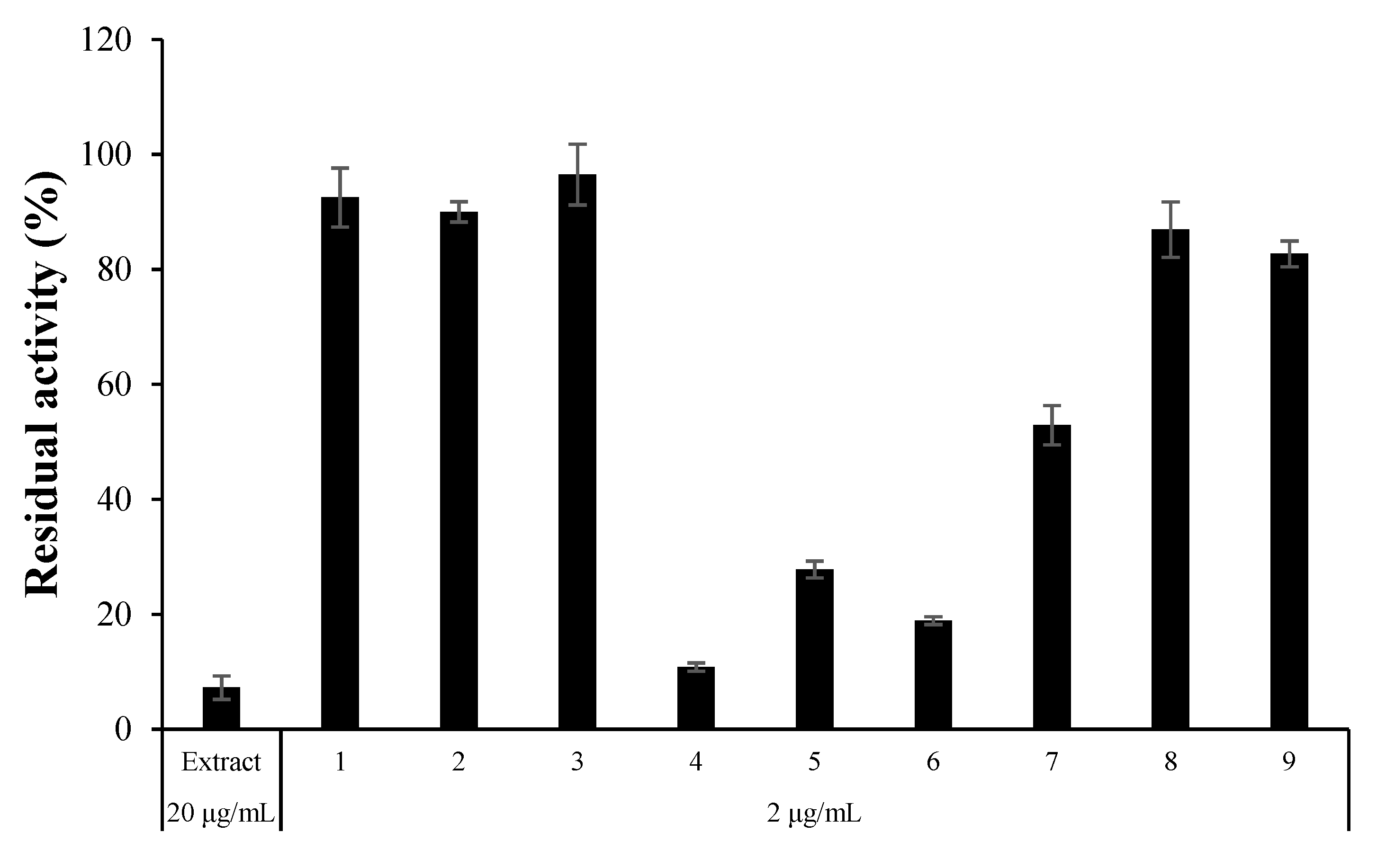
3.3. Inhibitory Activities of the Isolated Compounds against MAOs, ChEs, and BACE1
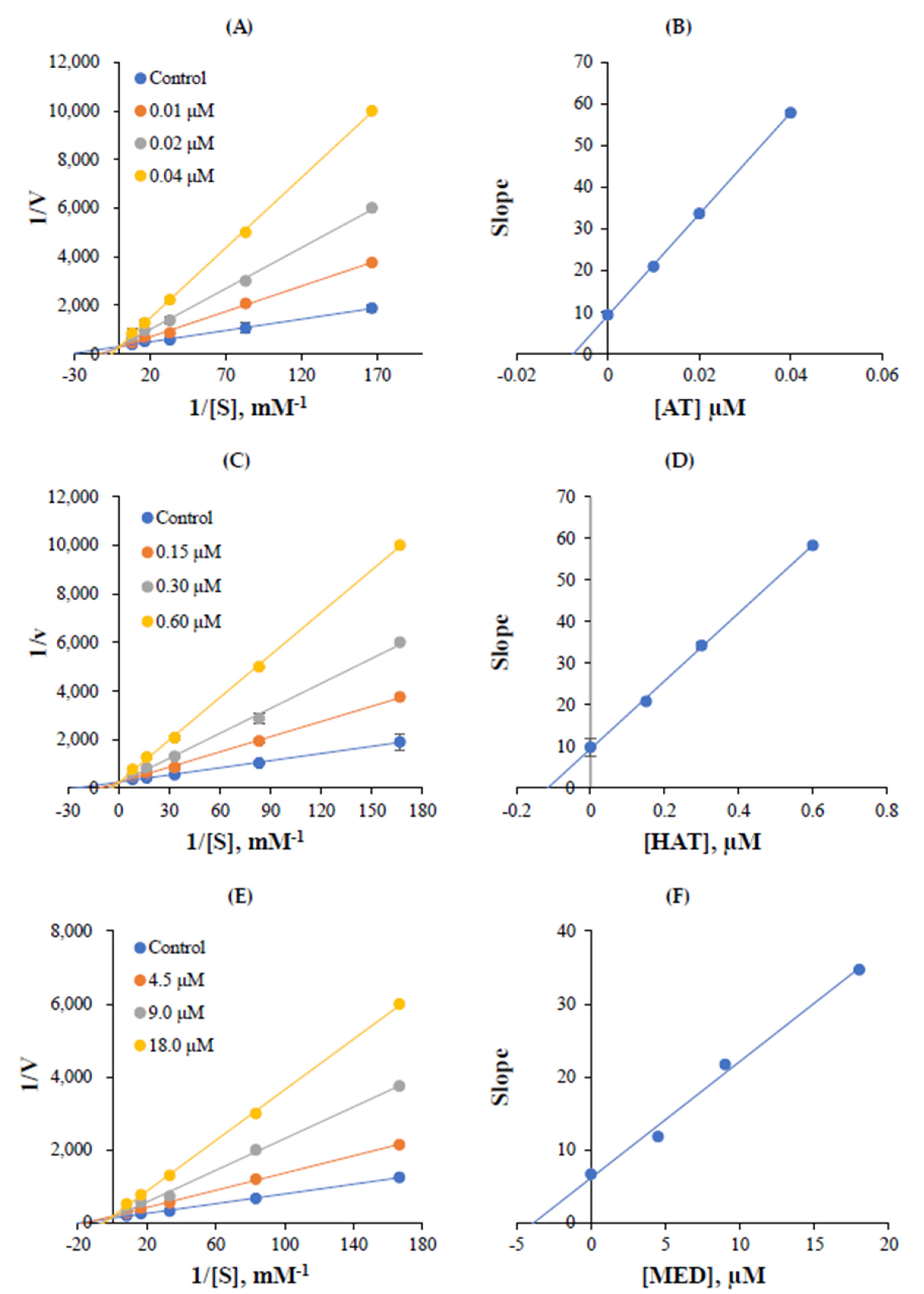
3.4. Kinetic Studies
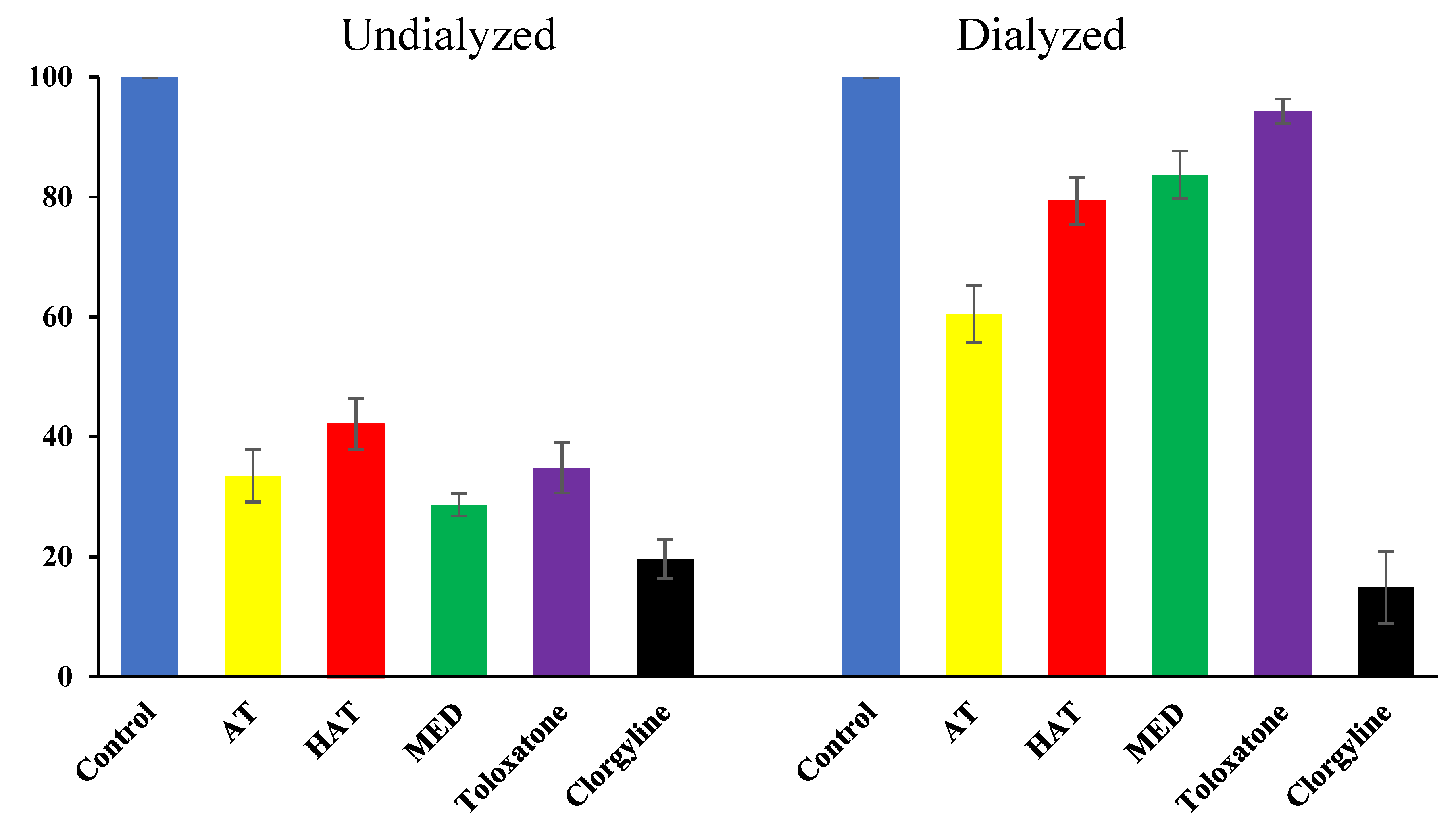
3.5. Reversibility Analysis of hMAO-A Inhibition by the Compounds
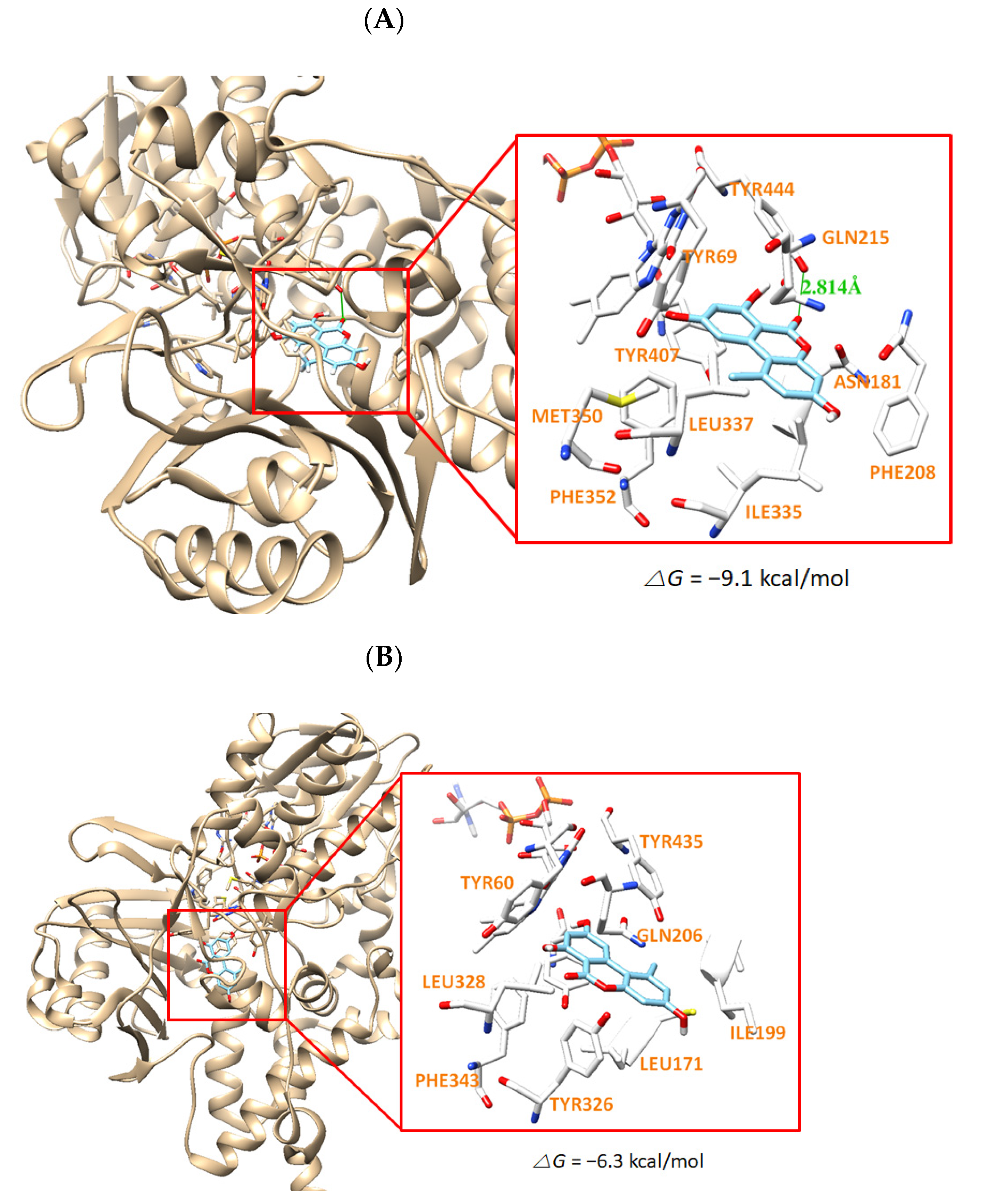
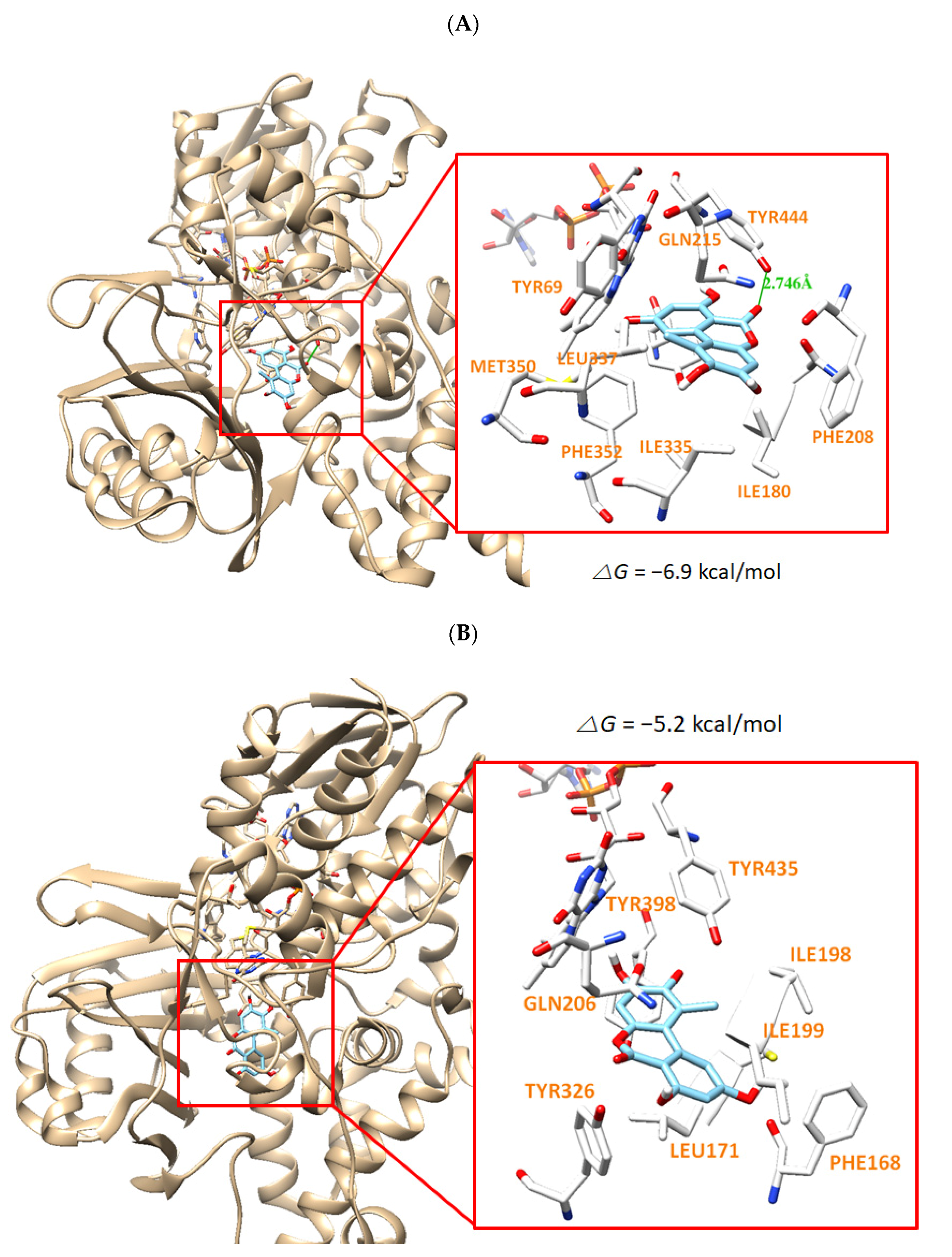
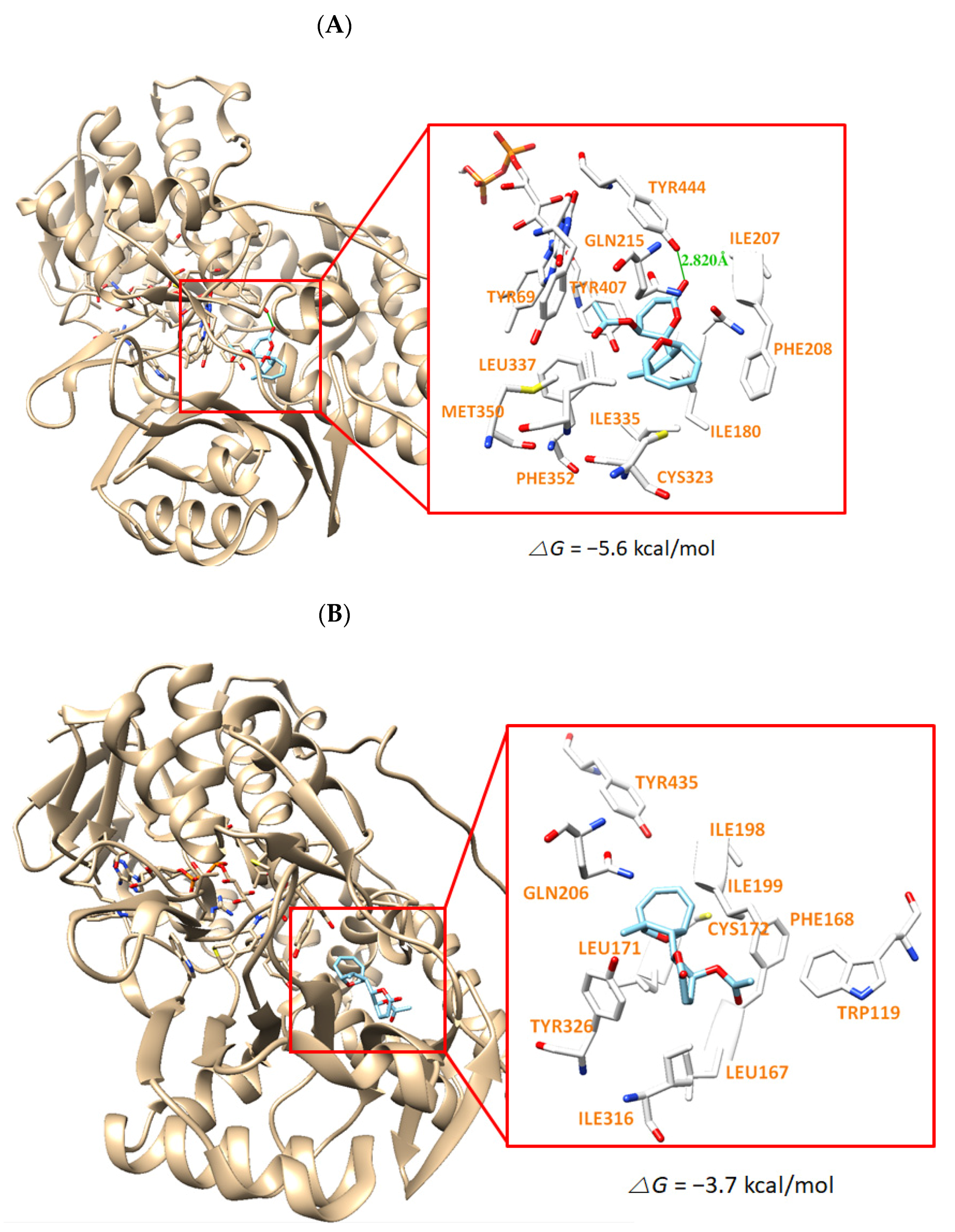
3.6. Molecular Docking Simulation
3.7. In Silico Pharmacokinetic Analysis of the Compounds
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Nash, T.H. Lichen Biology, 2nd ed.; Cambridge University Press: Cambridge, UK, 1996. [Google Scholar]
- Zambare, V.P.; Christopher, L.P. Biopharmaceutical potential of lichens. Pharm. Biol. 2012, 50, 778–798. [Google Scholar] [CrossRef]
- Ingólfsdóttir, K. Usnic acid. Phytochemistry 2002, 61, 729–736. [Google Scholar] [CrossRef]
- Kumar, K.C.; Müller, K. Lichen metabolites. 1. Inhibitory action against leukotriene B4 biosynthesis by a non-redox mechanism. J. Nat. Prod. 1999, 62, 817–820. [Google Scholar] [CrossRef]
- Bérdy, J. Bioactive microbial metabolites. J. Antibiot. 2005, 58, 1–26. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jeong, G.S.; Kang, M.G.; Han, S.A.; Noh, J.I.; Park, J.E.; Nam, S.J.; Park, D.; Yee, S.T.; Kim, H. Selective inhibition of human monoamine oxidase B by 5-hydroxy-2-methyl-chroman-4-one isolated from an endogenous lichen fungus Daldinia Fissa. J. Fungi. 2021, 7, 84. [Google Scholar] [CrossRef] [PubMed]
- de Zwart, P.L.; Jeronimus, B.F.; de Jonge, P. Empirical evidence for definitions of episode, remission, recovery, relapse and recurrence in depression: A systematic review. Epidemiol. Psychiatr. Sci. 2019, 28, 544–562. [Google Scholar] [CrossRef] [PubMed]
- Richard, B.S. Radical ideas about monoamine oxidase. Acc. Chem. Res. 1995, 28, 335–342. [Google Scholar]
- Berchtold, N.C.; Cotman, C.W. Evolution in the conceptualization of dementia and Alzheimer’s disease: Greco-Roman period to the 1960s. Neurobiol. Aging 1998, 19, 173–189. [Google Scholar] [CrossRef]
- Burns, A.; Iliffe, S. Alzheimer’s disease. BMJ 2009, 338, b158. [Google Scholar] [CrossRef] [Green Version]
- Ramsay, R.R. Monoamine oxidases: The biochemistry of the proteins as targets in medicinal chemistry and drug discovery. Curr. Top. Med. Chem. 2012, 12, 2189–2209. [Google Scholar] [CrossRef]
- Youdim, M.B.; Edmondson, D.; Tipton, K.F. The therapeutic potential of monoamine oxidase inhibitors. Nat. Rev. Neurosci. 2006, 7, 295–309. [Google Scholar] [CrossRef]
- Bierer, L.M.; Haroutunian, V.; Gabriel, S.; Knott, P.J.; Carlin, L.S.; Purohit, D.P.; Perl, D.P.; Schmeidler, J.; Kanof, P.; Davis, K.L. Neurochemical correlates of dementia severity in Alzheimer’s disease: Relative importance of the cholinergic deficits. J. Neurochem. 1995, 64, 749–760. [Google Scholar] [CrossRef]
- Kumar, A.; Pintus, F.; Di Petrillo, A.; Medda, R.; Caria, P.; Matos, M.J.; Viña, D.; Pieroni, E.; Delogu, F.; Era, B.; et al. Novel 2-pheynlbenzofuran derivatives as selective butyrylcholinesterase inhibitors for Alzheimer’s disease. Sci. Rep. 2018, 8, 4424. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Colović, M.B.; Krstić, D.Z.; Lazarević-Pašti, T.D.; Bondžić, A.M.; Vasić, V.M. Acetylcholinesterase inhibitors: Pharmacology and toxicology. Curr. Neuropharmacol. 2013, 11, 315–335. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Giacobini, E. Cholinesterase inhibitors: New roles and therapeutic alternatives. Pharmacol. Res. 2004, 50, 433–440. [Google Scholar] [CrossRef] [PubMed]
- Ghosh, A.K.; Osswald, H.L. BACE1 (β-secretase) inhibitors for the treatment of Alzheimer’s disease. Chem. Soc. Rev. 2014, 43, 6765–6813. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vejdovszky, K.; Hahn, K.; Braun, D.; Warth, B.; Marko, D. Synergistic estrogenic effects of Fsarium and Alternaria mycotoxins in vitro. Arch. Toxicol. 2017, 91, 1447–1460. [Google Scholar] [CrossRef] [Green Version]
- Schmutz, C.; Cenk, E.; Marko, D. The Alternaria mycotoxin alternariol triggers the immune response of IL-1β-stimulated, differentiated Caco-2 Cells. Mol. Nutr. Food Res. 2019, 63, e1900341. [Google Scholar] [CrossRef] [Green Version]
- Solhaug, A.; Eriksen, G.S.; Holme, J.A. Mechanisms of action and toxicity of the mycotoxin alternariol: A Review. Basic Clin. Pharmacol. Toxicol. 2016, 119, 533–539. [Google Scholar] [CrossRef]
- Osman, M.Y.H.; Osman, H.M.Y. Dual effect of Alternariol on Acetylcholinestrase and Monoamine oxidase Extracted from Different Parts of Rat Brain. FASEB J. 2009, 23, 676.2. [Google Scholar] [CrossRef]
- Tao, M.H.; Chen, Y.C.; Wei, X.Y.; Tan, J.W.; Zhang, W.M. Chemical constituents of the endophytic fungus phomopsis sp. A240 isolated from Taxus chinensis var. mairei. Helv. Chim. Acta. 2014, 97, 426–430. [Google Scholar] [CrossRef]
- Tao, M.; Chen, Y.; Zhang, W. Application of 5′-hydroxy Alternariol in Preparation of Antioxidants or Cosmetics. China Patent CN102488683 A, 13 June 2012. [Google Scholar]
- Lin, X.; Huang, Y.; Fang, M.; Wang, J.; Zheng, Z.; Su, W. Cytotoxic and antimicrobial metabolites from marine lignicolous fungi, Diaporthe sp. FEMS Microbiol. Lett. 2005, 251, 53–58. [Google Scholar] [CrossRef] [Green Version]
- Zhu, J.; Chen, Q.; Xia, X.; Mo, P.; Shen, Y.; Yu, C. Mycoepoxydiene suppresses RANKL-induced osteoclast differentiation and reduces ovariectomy-induced bone loss in mice. Appl. Microbiol. Biotechnol. 2013, 97, 767–774. [Google Scholar] [CrossRef]
- Wang, J.; Zhao, B.; Zhang, W.; Wu, X.; Wang, R.; Huang, Y.; Chen, D.; Park, K.; Weimer, B.C.; Shen, Y. Mycoepoxydiene, a fungal polyketide, induces cell cycle arrest at the G2/M phase and apoptosis in HeLa cells. Bioorg. Med. Chem. Lett. 2010, 20, 7054–7058. [Google Scholar] [CrossRef] [PubMed]
- Wang, J.; Zhao, B.; Yi, Y.; Zhang, W.; Wu, X.; Zhang, L.; Shen, Y. Mycoepoxydiene, a fungal polyketide inhibits MCF-7 cells through simultaneously targeting p53 and NF-κB pathways. Biochem. Pharmacol. 2012, 84, 891–899. [Google Scholar] [CrossRef]
- Jin, K.; Li, L.; Sun, X.; Xu, Q.; Song, S.; Shen, Y.; Deng, X. Mycoepoxydiene suppresses HeLa cell growth by inhibiting glycolysis and the pentose phosphate pathway. Appl. Microbiol. Biotechnol. 2017, 101, 4201–4213. [Google Scholar] [CrossRef] [PubMed]
- Chen, Q.; Chen, T.; Li, W.; Zhang, W.; Zhu, J.; Li, Y.; Huang, Y.; Shen, Y.; Yu, C. Mycoepoxydiene inhibits lipopolysaccharide-induced inflammatory responses through the suppression of TRAF6 polyubiquitination. PloS ONE 2012, 7, e44890. [Google Scholar]
- Lee, H.W.; Ryu, H.W.; Kang, M.G.; Park, D.; Oh, S.R.; Kim, H. Potent selective monoamine oxidase B inhibition by maackiain, a pterocarpan from the roots of Sophora flavescens. Bioorg. Med. Chem. Lett. 2016, 26, 4714–4719. [Google Scholar] [CrossRef]
- Baek, S.C.; Choi, B.; Nam, S.J.; Kim, H. Inhibition of monoamine oxidase A and B by demethoxycurcumin and bisdemethoxycurcumin. J. Appl. Biol. Chem. 2018, 61, 187–190. [Google Scholar] [CrossRef]
- Ellman, G.L.; Courtney, K.D.; Andres, J.V.; Feather-Stone, R.M. A new and rapid colorimetric determination of acetylcholinesterase activity. Biochem. Pharmacol. 1961, 7, 88–95. [Google Scholar] [CrossRef]
- Lee, J.P.; Kang, M.G.; Lee, J.Y.; Oh, J.M.; Baek, S.C.; Leem, H.H.; Park, D.; Cho, M.L.; Kim, H. Potent inhibition of acetylcholinesterase by sargachromanol I from Sargassum siliquastrum and by selected natural compounds. Bioorg. Chem. 2019, 89, 103043. [Google Scholar] [CrossRef] [PubMed]
- Wilson, E.N.; Do Carmo, S.; Iulita, M.F.; Hall, H.; Ducatenzeiler, A.; Marks, A.R.; Allard, S.; Jia, D.T.; Windheim, J.; Cuello, A.C. BACE1 inhibition by microdose lithium formulation NP03 rescues memory loss and early stage amyloid neuropathology. Transl. Psychiatry 2017, 7, e1190. [Google Scholar] [CrossRef] [Green Version]
- Baek, S.C.; Lee, H.W.; Ryu, H.W.; Kang, M.G.; Park, D.; Kim, S.H.; Cho, M.L.; Oh, S.R.; Kim, H. Selective inhibition of monoamine oxidase A by hispidol. Bioorg. Med. Chem. Lett. 2018, 28, 584–588. [Google Scholar] [CrossRef]
- Baek, S.C.; Park, M.H.; Ryu, H.W.; Lee, J.P.; Kang, M.G.; Park, D.; Park, C.M.; Oh, S.R.; Kim, H. Rhamnocitrin isolated from Prunus padus var. seoulensis: A potent and selective reversible inhibitor of human monoamine oxidase A. Bioorg. Chem. 2019, 83, 317–325. [Google Scholar] [CrossRef]
- Trott, O.; Olson, A.J. AutoDock Vina: Improving the speed and accuracy of docking with a new scoring function, efficient optimization, and multithreading. J. Comput. Chem. 2010, 31, 455–461. [Google Scholar] [CrossRef] [Green Version]
- Mills, J.E.; Dean, P.M. Three-dimensional hydrogen-bond geometry and probability information from a crystal survey. J. Comput. Aided. Mol. Des. 1996, 10, 607–622. [Google Scholar] [CrossRef]
- Pettersen, E.F.; Goddard, T.D.; Huang, C.C.; Couch, G.S.; Greenblatt, D.M.; Meng, E.C.; Ferrin, T.E. UCSF Chimera--a visualization system for exploratory research and analysis. J. Comput. Chem. 2004, 25, 1605–1612. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Phillips, J.C.; Hardy, D.J.; Maia, J.; Stone, J.E.; Ribeiro, J.V.; Bernardi, R.C.; Buch, R.; Fiorin, G.; Hénin, J.; Jiang, W.; et al. Scalable molecular dynamics on CPU and GPU architectures with NAMD. J. Chem. Phys. 2020, 153, 044130. [Google Scholar] [CrossRef]
- Humphrey, W.; Dalke, A.; Schulten, K. VMD: Visual molecular dynamics. J. Mol. Graph. 1996, 14, 33–38. [Google Scholar] [CrossRef]
- Vanommeslaeghe, K.; Hatcher, E.; Acharya, C.; Kundu, S.; Zhong, S.; Shim, J.; Darian, E.; Guvench, O.; Lopes, P.; Vorobyov, I.; et al. CHARMM general force field: A force field for drug-like molecules compatible with the CHARMM all-atom additive biological force fields. J. Comput. Chem. 2010, 31, 671–690. [Google Scholar] [CrossRef] [Green Version]
- Daina, A.; Michielin, O.; Zoete, V. SwissADME: A free web tool to evaluate pharmacokinetics, drug-likeness and medicinal chemistry friendliness of small molecules. Sci. Rep. 2017, 7, 42717. [Google Scholar] [CrossRef] [Green Version]
- de Souza, G.D.; Mithöfer, A.; Daolio, C.; Schneider, B.; Rodrigues-Filho, E. Identification of Alternaria alternata mycotoxins by LC-SPE-NMR and their cytotoxic effects to soybean (Glycine max) cell suspension culture. Molecules 2013, 18, 2528–2538. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, W.; Hao, B.; Du, L.; Shen, Y. Cytotoxic Polyketides with an Oxygen-Bridged Cyclooctadiene Core Skeleton from the Mangrove Endophytic Fungus Phomosis sp. A818. Molecules 2017, 22, 1547. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Luo, H.; Li, C.; Kim, J.C.; Liu, Y.; Jung, J.S.; Koh, Y.J.; Hur, J.S. Biruloquinone, an acetylcholinesterase inhibitor produced by lichen-forming fungus Cladonia macilenta. J. Microbiol. Biotechnol. 2013, 23, 161–166. [Google Scholar] [CrossRef] [PubMed]
- de Barros Alves, G.M.; de Sousa Maia, M.B.; de Souza Franco, E.; Galvão, A.M.; da Silva, T.G.; Gomes, R.M.; Martins, M.B.; da Silva Falcão, E.P.; de Castro, C.M.; da Silva, N.H. Expectorant and antioxidant activities of purified fumarprotocetraric acid from Cladonia verticillaris lichen in mice. Pulm. Pharmacol. Ther. 2014, 27, 139–143. [Google Scholar] [CrossRef] [PubMed]
- Coskun, Z.M.; Ersoz, M.; Acikgoz, B.; Karalti, I.; Cobanoglu, G.; Sesal, C. Anti-Proliferative and Apoptotic Effects of Methanolic Extracts from Different Cladonia Species on Human Breast Cancer Cells. Folia Biol. 2015, 61, 97–103. [Google Scholar]
- Williamson, P.M.; Highet, A.S.; Gams, W.; Sivasithamparam, K.; Cowling, W.A. Diaporthe toxica sp. nov., the cause of lupinosis in sheep. Mycol. Res. 1994, 98, 1364–1368. [Google Scholar] [CrossRef]
- Gao, Y.; Su, Y.; Sun, W.; Cai, L. Diaporthe species occurring on Lithocarpus glabra in China, with descriptions of five new species. Fungal Biol. 2015, 119, 295–309. [Google Scholar] [CrossRef]
- Okuyama, E.; Hossain, C.F.; Yamazaki, M. Monoamine oxidase inhibitors from a lichen, Solorina crocea (L.) ACH. Jpn. J. Pharmacogn. 1991, 45, 159–162. [Google Scholar]
- Kinoshita, K.; Saito, D.; Koyama, K.; Takahashi, K.; Sato, Y.; Okuyama, E.; Fujimoto, H.; Yamazaki, M. Monoamine oxidase inhibitory effects of some lichen compounds and their synthetic analogues. J. Hattori. Bot. Lab. 2002, 92, 277–284. [Google Scholar]
- Davis, V.M.; Stack, M.E. Evaluation of alternariol and alternariol methyl ether for mutagenic activity in Salmonella Typhimurium. Appl. Environ. Microbiol. 1994, 60, 3901–3902. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lee, H.W.; Kim, Y.J.; Nam, S.J.; Kim, H. Potent Selective Inhibition of Monoamine Oxidase A by Alternariol Monomethyl Ether Isolated from Alternaria Brassicae. J. Microbiol. Biotechnol. 2017, 27, 316–320. [Google Scholar] [CrossRef] [PubMed]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| ELF No. | Residual Activity at 20 µg/mL (%) | |||
|---|---|---|---|---|
| hMAO-A | hMAO-B | AChE | BChE | |
| 3 | 14.9 ± 1.91 | 25.2 ± 5.49 | 87.9 ± 4.29 | 95.2 ± 1.42 |
| 5 | 48.3 ± 0.48 | 68.9 ± 3.14 | 72.0 ± 9.02 | 14.5 ± 1.81 |
| 12 | 68.9 ± 0.96 | 13.5 ± 0.52 | 90.2 ± 1.24 | 72.8 ± 3.30 |
| 19 | 81.7 ± 4.84 | 10.9 ± 2.06 | 91.9 ± 0.84 | 93.5 ± 2.45 |
| 21 | 4.18 ± 2.69 | −4.67 ± 1.32 | 63.3 ± 1.32 | 64.8 ± 1.20 |
| 23 | 80.1 ± 6.66 | 68.2 ± 6.61 | 86.6 ± 2.80 | 20.9 ± 1.84 |
| 24 | 7.25 ± 1.02 | 2.80 ± 0.42 | 56.6 ± 0.98 | 93.2 ± 6.70 |
| 28 | 35.1 ± 5.64 | 18.6 ± 0.001 | 95.7 ± 4.22 | 71.3 ± 0.24 |
| 29 | 7.25 ± 2.05 | 10.8 ± 1.39 | 94.3 ± 1.42 | 86.3 ± 2.92 |
| 45 | 14.8 ± 0.47 | 41.0 ± 2.26 | 91.1 ± 3.14 | 83.3 ± 0.00 |
| 49 | 15.0 ± 0.40 | 34.6 ± 5.27 | 97.1 ± 6.09 | 26.2 ± 1.59 |
| 50 | 50.0 ± 2.80 | 10.1 ± 2.26 | 94.4 ± 1.17 | 45.2 ± 2.96 |
| 54 | 71.4 ± 3.59 | 43.2 ± 5.41 | 86.8 ± 2.33 | 22.2 ± 0.50 |
| 93 | 9.83 ± 0.48 | −1.70 ± 7.23 | 78.0 ± 4.09 | 56.1 ± 2.20 |
| 114 | 16.7 ± 0.55 | 15.5 ± 1.68 | 75.5 ± 6.43 | 97.2 ± 0.72 |
| ELF No. | Residual Activity at 20 µg/mL (%) | |||
|---|---|---|---|---|
| Original Extract | Cultured Extract | |||
| hMAO-A | hMAO-B | hMAO-A | hMAO-B | |
| 21 | 4.18 ± 2.69 | −4.67 ± 1.32 | >50 | 45.6 ± 8.71 |
| 29 | 7.25 ± 2.05 | 10.8 ± 1.39 | 10.7 ± 4.38 | 53.4 ± 10.1 |
| 93 | 9.83 ± 0.48 | −1.70 ± 7.23 | >50 | >50 |
| No. | Sample Name | Residual Activity at 2 µg/mL (%) | Name | MW |
|---|---|---|---|---|
| hMAO-A | ||||
| 1 | LFF29-C18-40L-F4-1&2-T1 | 88.2 ± 3.07 | ||
| 2 | LFF29-C18-40L-F4-1&2-T2 | 85.5 ± 4.17 | ||
| 3 | LFF29-C18-40L-F4-1&2-T3 | 86.9 ± 0.47 | ||
| 4 | LFF29-C18-40L-F4-3 | 3.39 ± 3.43 | 5′-hydroxy-alternariol | 274.23 |
| 5 | LFF29-C18-40L-F4-3-1 | 44.9 ± 5.90 | ||
| 6 | LFF29-C18-40L-F4-3-2 | 82.0 ± 0.92 | ||
| 7 | LFF29-C18-40L-F4-4-4 | 66.7 ± 1.84 | Mycoepoxydiene | 290.32 |
| 8 | LFF29-C18-40L-F4-4-5 | −5.41 ± 0.98 | Alternariol | 258.05 |
| 9 | LFF29-C18-40L-F4-4-6 | 14.7 ± 1.85 | ||
| 10 | LFF29-C18-40L-F4-4-7 | 46.6 ± 2.12 | ||
| 11 | LFF29-C18-40L-F4-4-8 | 90.5 ± 1.30 | ||
| 12 | LFF29-C18-40L-F4-4-9 | 80.8 ± 0.46 |
| Residual Activity at 10 µM (%) | |||||
|---|---|---|---|---|---|
| hMAO-A | hMAO-B | AChE | BChE | BACE1 | |
| AT a | −3.1 ± 0.0 | 75.6 ± 2.3 | 58.7 ± 2.0 | 93.3 ± 4.1 | 80.9 ± 0.2 |
| HAT a | 17.7 ± 1.1 | 83.3 ± 4.0 | 65.6 ± 3.8 | 96.2 ± 2.3 | 71.3 ± 3.8 |
| MED | 42.7 ± 0.9 | 85.4 ± 6.9 | 65.2 ± 2.7 | 95.5 ± 5.5 | 80.5 ± 2.7 |
| IC50 (µM) | SI b | ||||
| hMAO-A | hMAO-B | AChE | BACE1 | ||
| AT a | 0.020 ± 0.001 | 20.7 ± 2.2 | 10.0 ± 0.9 | 36.7 ± 3.3 | 1035 |
| HAT a | 0.31 ± 0.02 | >40 | 19.9 ± 0.2 | 24.7 ± 1.7 | >129 |
| MED | 8.7 ± 0.3 | >40 | 18.6 ± 1.3 | 35.9 ± 0.3 | >4.59 |
| Toloxatone | 1.1 ± 0.03 | ||||
| Clorgyline | 0.007 ± 0.001 | ||||
| Lazabemide | 0.063 ± 0.015 | ||||
| Pargyline | 0.028 ± 0.004 | ||||
| Donepezil | 0.009 ± 0.002 | ||||
| Tacrine | 0.27 ± 0.02 | ||||
| Quercetin | 20.5 ± 0.6 | ||||
| Compound | GI Absorption | BBB Permeant | P-gp Substrate | CYP1A2 Inhibitor | CYP2C19 Inhibitor | CYP2C9 Inhibitor | CYP2D6 Inhibitor | CYP3A4 Inhibitor | Log Kp (Skin Permeation) |
|---|---|---|---|---|---|---|---|---|---|
| AT | High | No | No | Yes | No | No | Yes | No | −6.18 cm/s |
| HAT | High | No | No | Yes | No | No | No | No | −5.82 cm/s |
| MED | High | Yes | No | No | No | No | No | No | −6.67 cm/s |
| Compound | Mw (g/mol) | cLog P | HBD | HBA | TPSA (Ų) | RB | Lipinski Violations |
|---|---|---|---|---|---|---|---|
| AT | 274.23 | 1.83 | 4 | 6 | 111.13 | 0 | 0 |
| HAT | 258.23 | 2.17 | 3 | 5 | 90.9 | 0 | 0 |
| MED | 290.31 | 1.67 | 0 | 5 | 61.83 | 3 | 0 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Jeong, G.S.; Hillman, P.F.; Kang, M.-G.; Hwang, S.; Park, J.-E.; Nam, S.-J.; Park, D.; Kim, H. Potent and Selective Inhibitors of Human Monoamine Oxidase A from an Endogenous Lichen Fungus Diaporthe mahothocarpus. J. Fungi 2021, 7, 876. https://0-doi-org.brum.beds.ac.uk/10.3390/jof7100876
Jeong GS, Hillman PF, Kang M-G, Hwang S, Park J-E, Nam S-J, Park D, Kim H. Potent and Selective Inhibitors of Human Monoamine Oxidase A from an Endogenous Lichen Fungus Diaporthe mahothocarpus. Journal of Fungi. 2021; 7(10):876. https://0-doi-org.brum.beds.ac.uk/10.3390/jof7100876
Chicago/Turabian StyleJeong, Geum Seok, Prima F. Hillman, Myung-Gyun Kang, Sungbo Hwang, Jong-Eun Park, Sang-Jip Nam, Daeui Park, and Hoon Kim. 2021. "Potent and Selective Inhibitors of Human Monoamine Oxidase A from an Endogenous Lichen Fungus Diaporthe mahothocarpus" Journal of Fungi 7, no. 10: 876. https://0-doi-org.brum.beds.ac.uk/10.3390/jof7100876