
Multigene Phylogeny and Morphology Reveal Unexpectedly High Number of New Species of Cantharellus Subgenus Parvocantharellus (Hydnaceae, Cantharellales) in China

Abstract
:1. Introduction
2. Materials and Methods
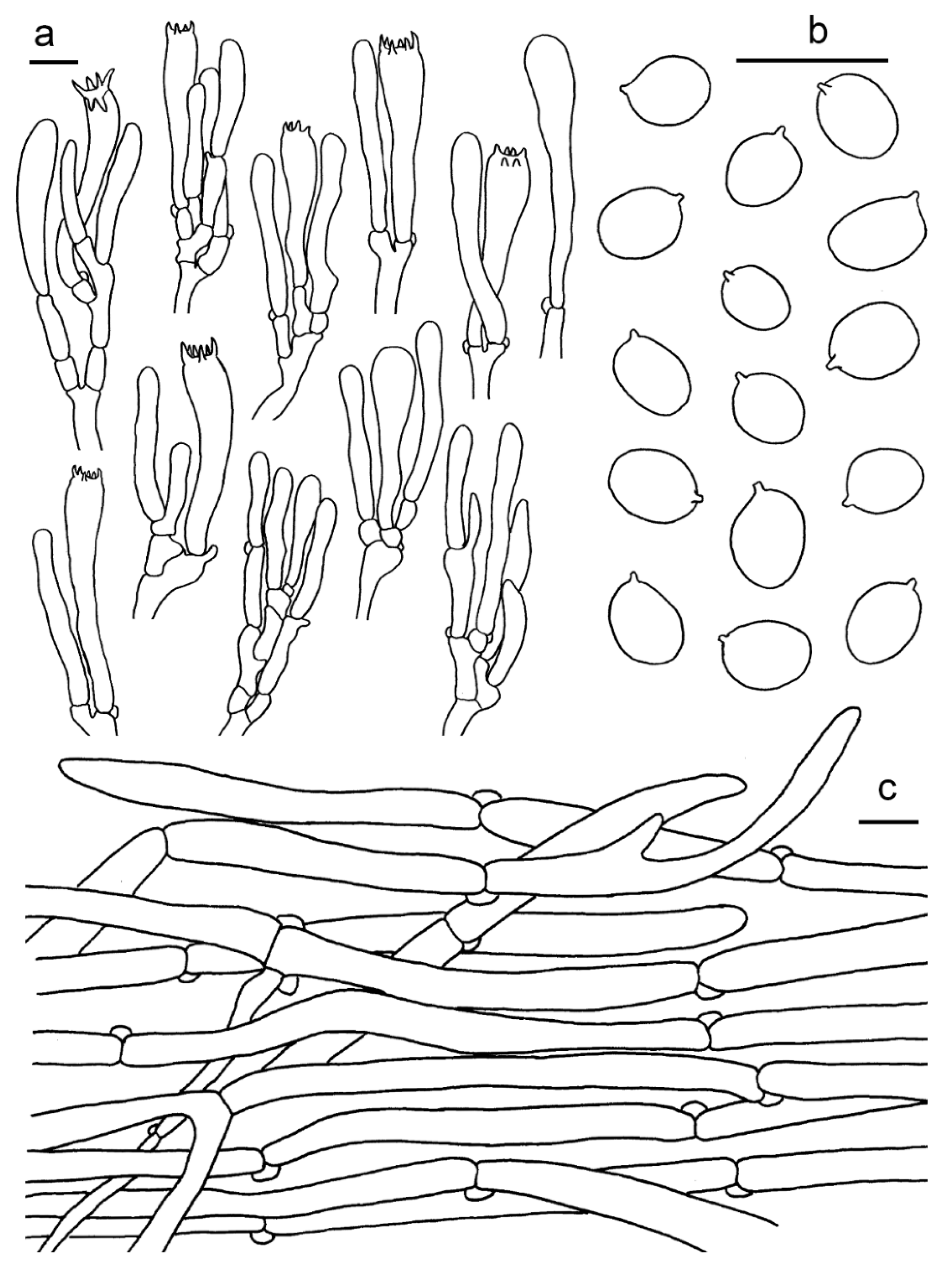
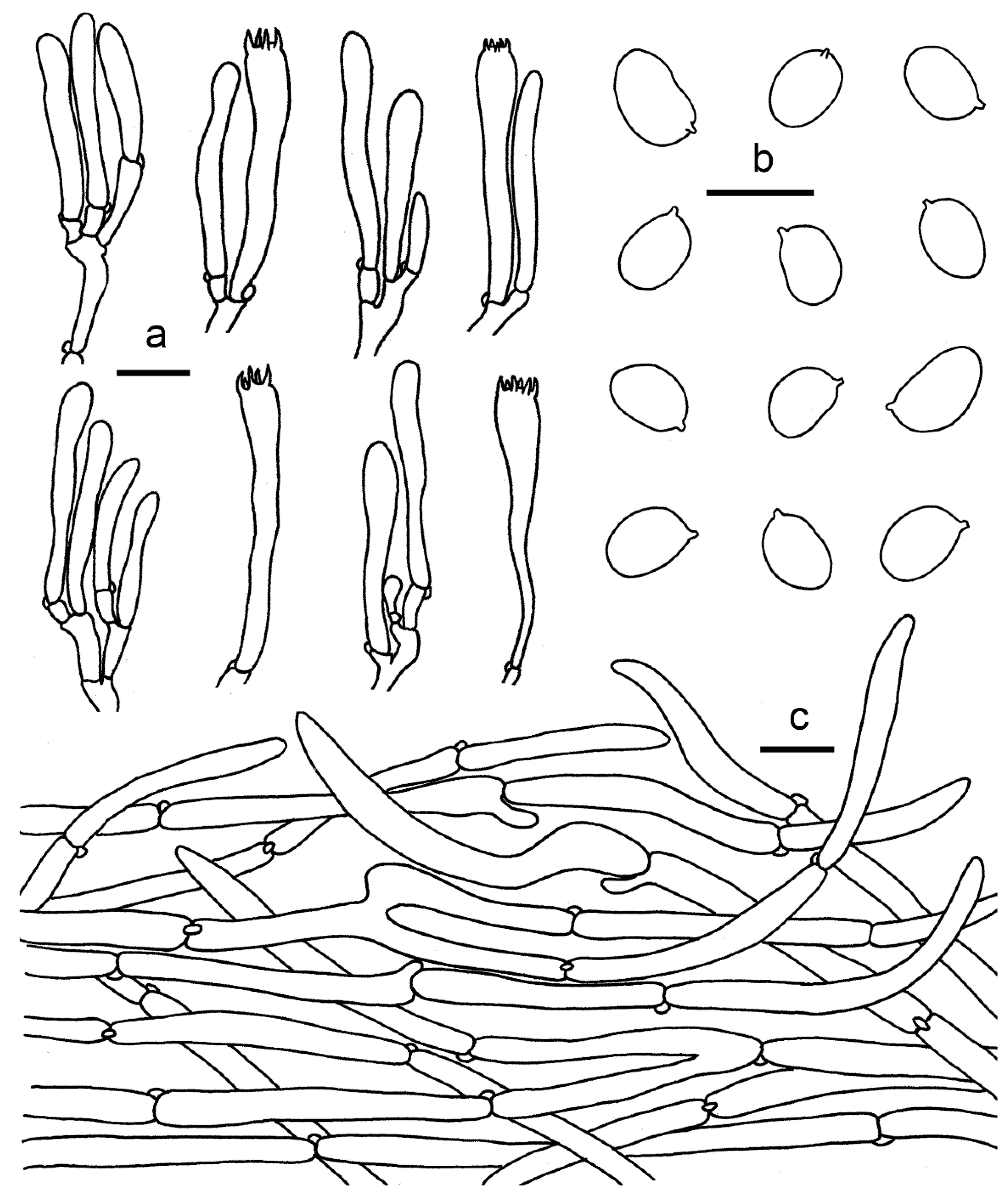
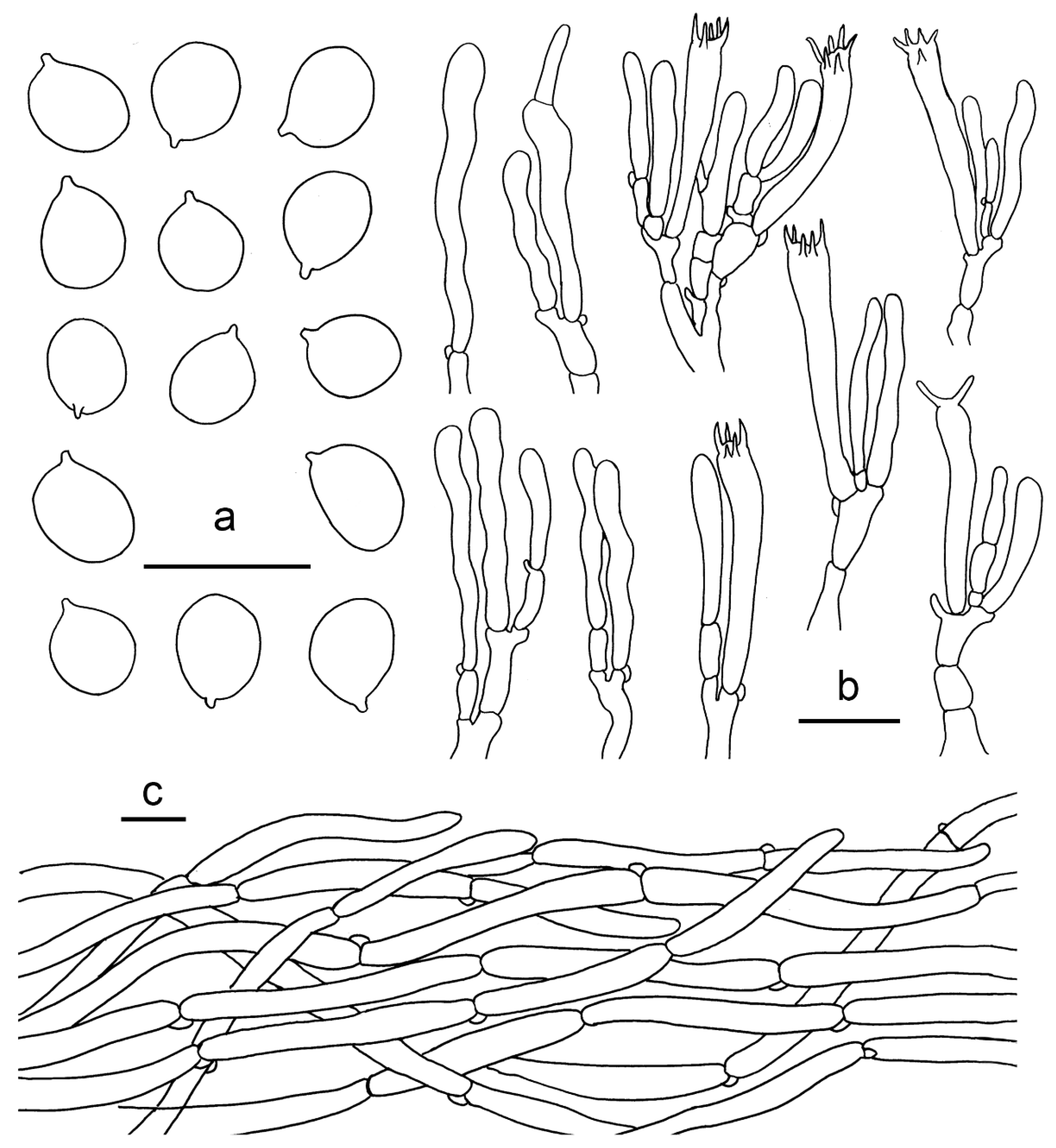
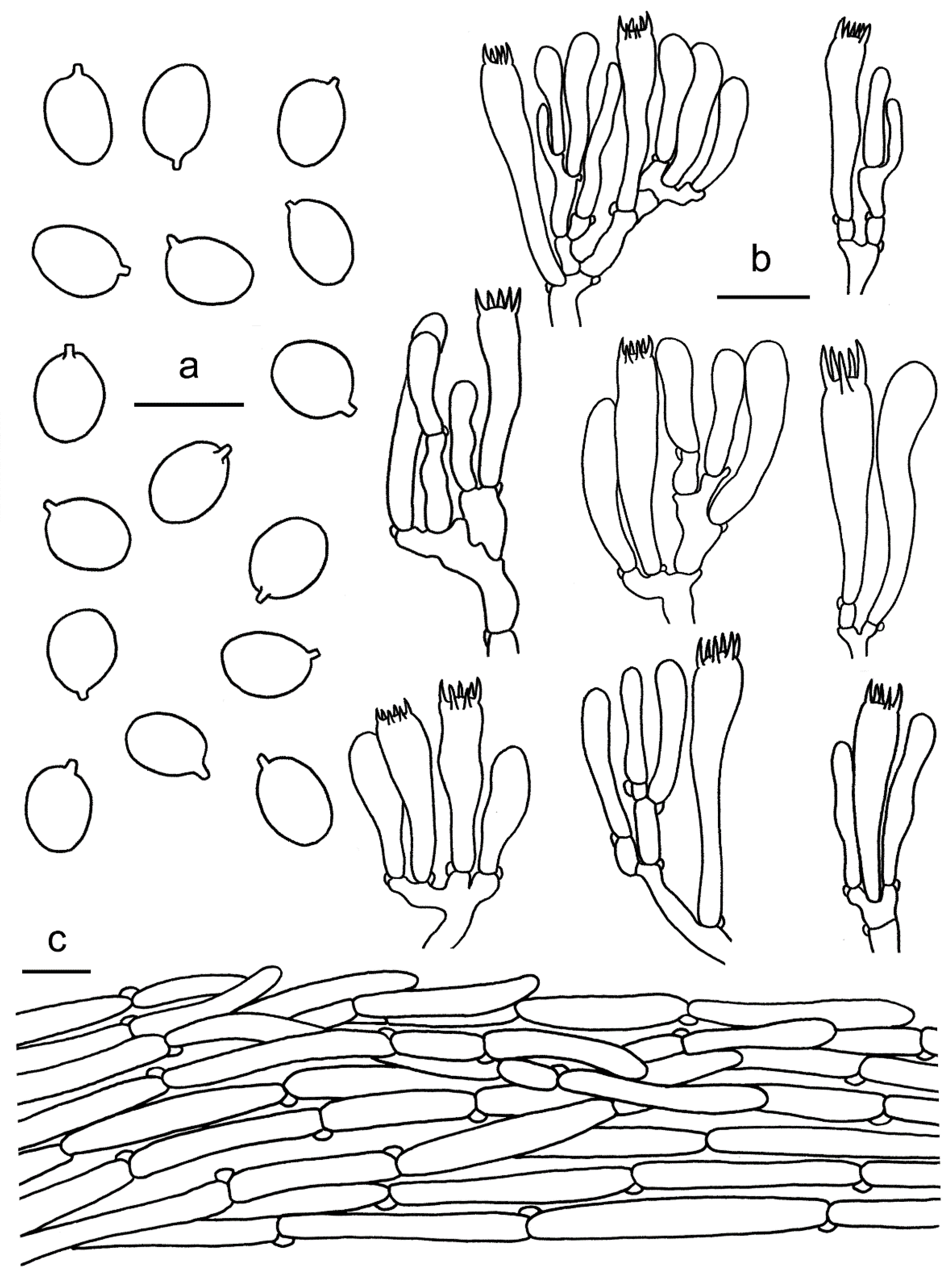
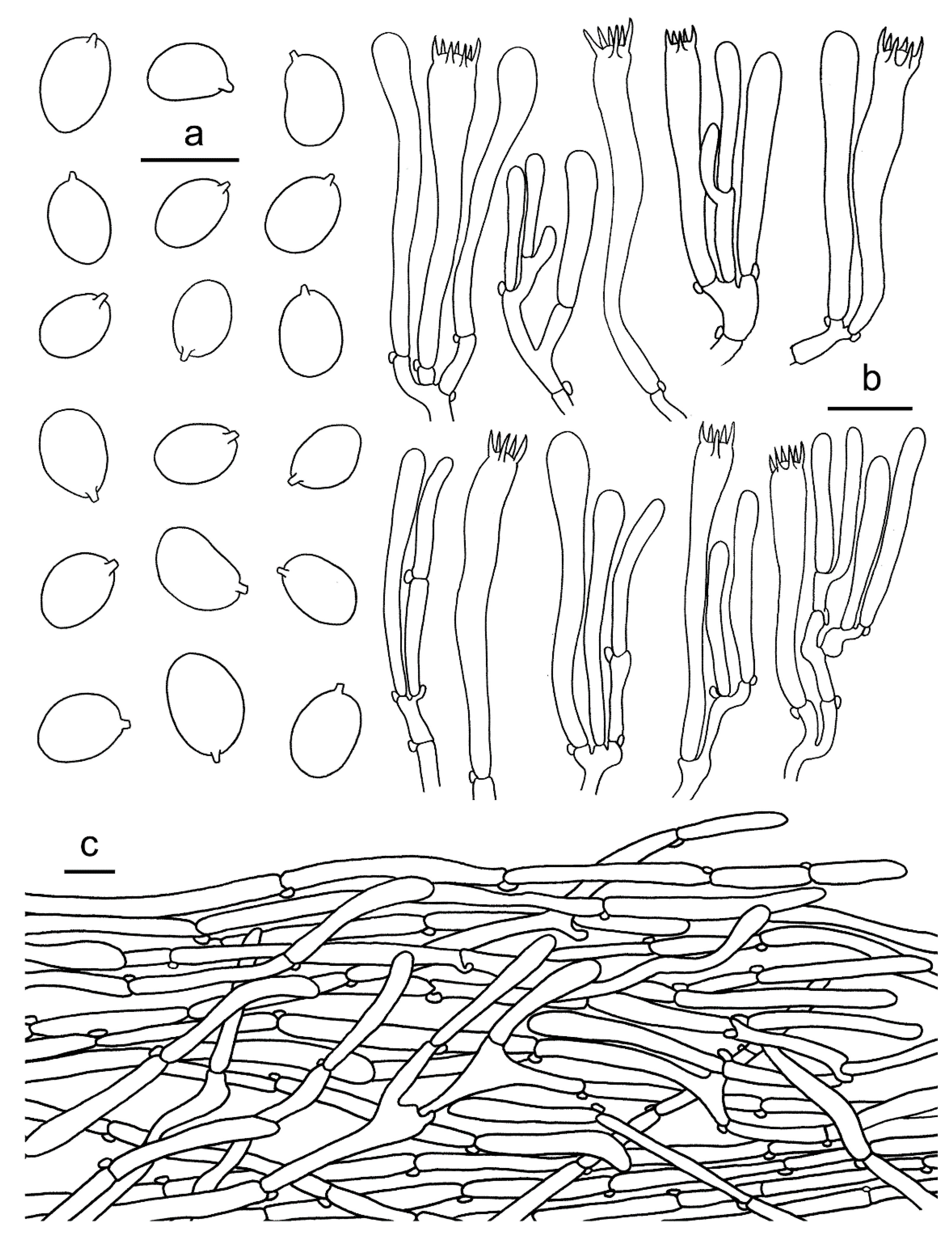
2.1. Morphological Studies
2.2. DNA Extraction, PCR Amplification and Sequencing
2.3. Phylogenetic Analyses
3. Results
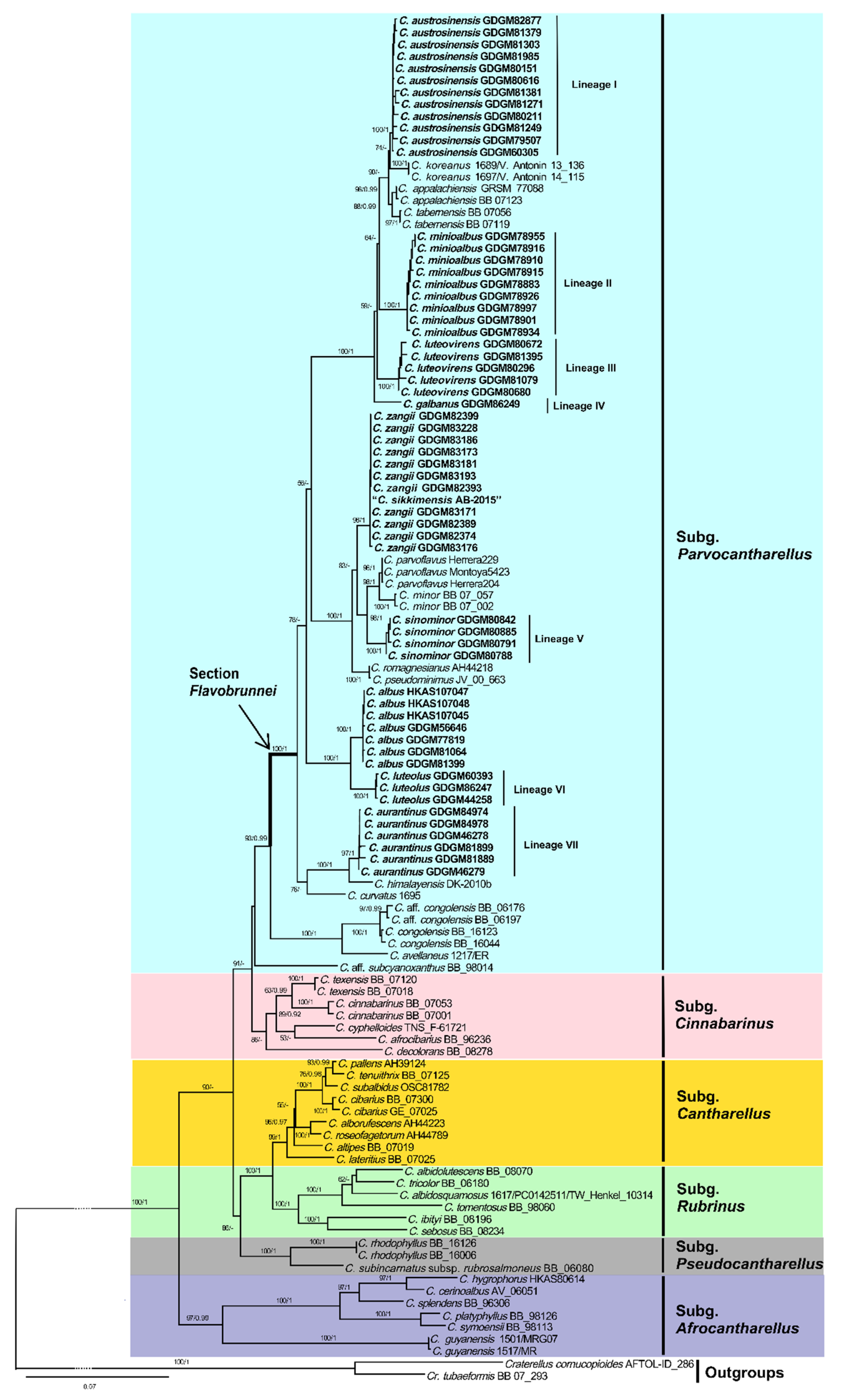
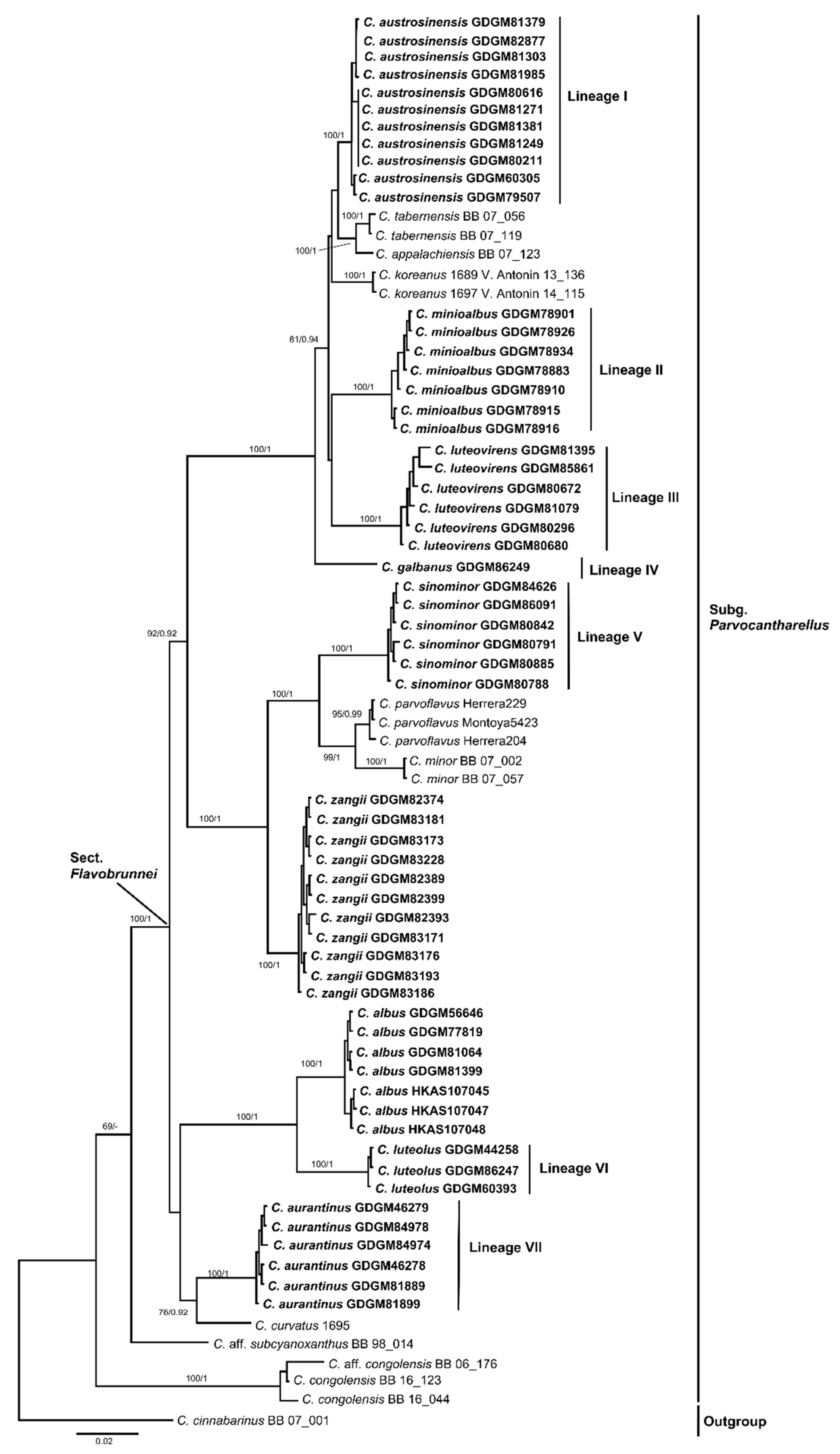
3.1. Molecular Phylogeny
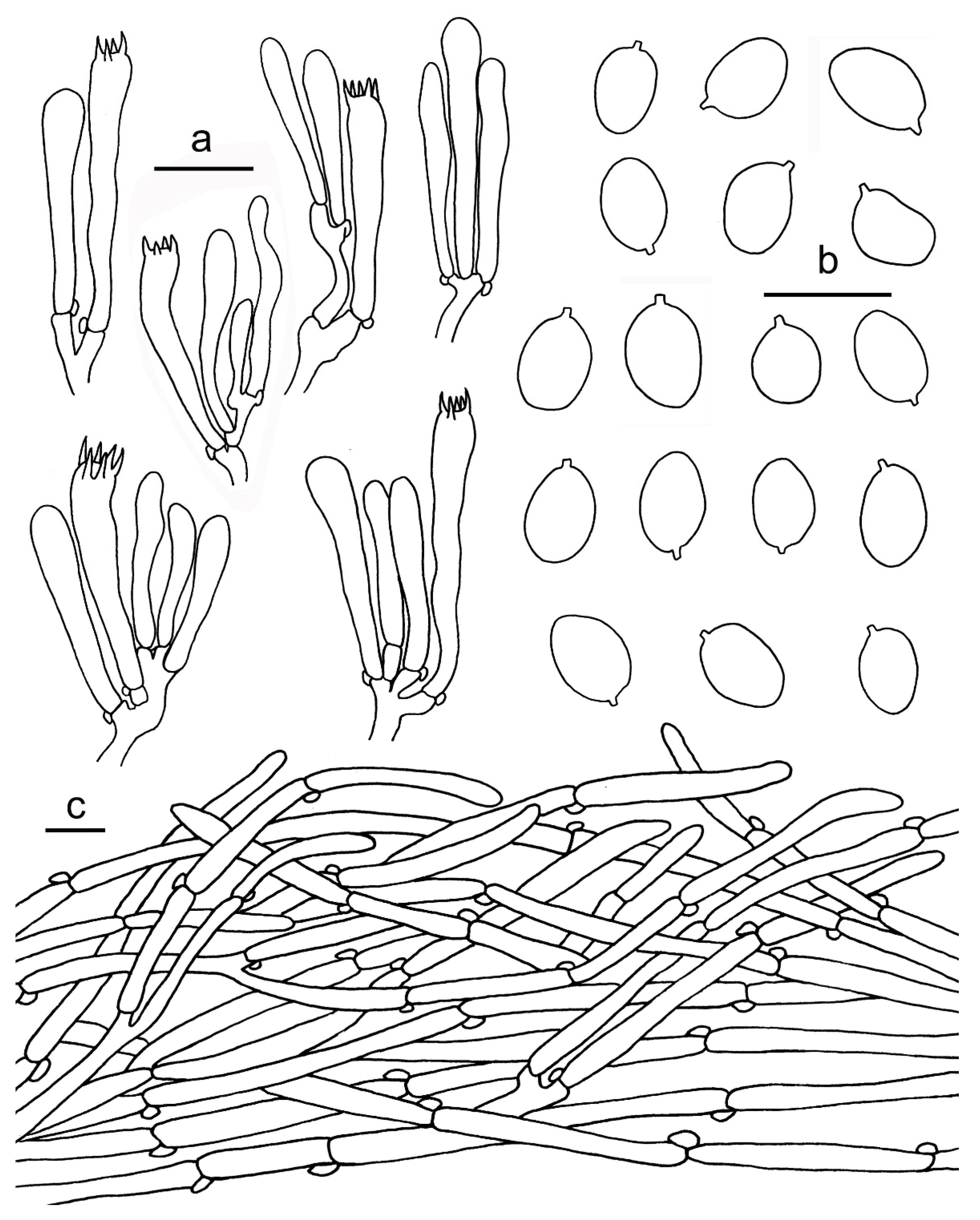
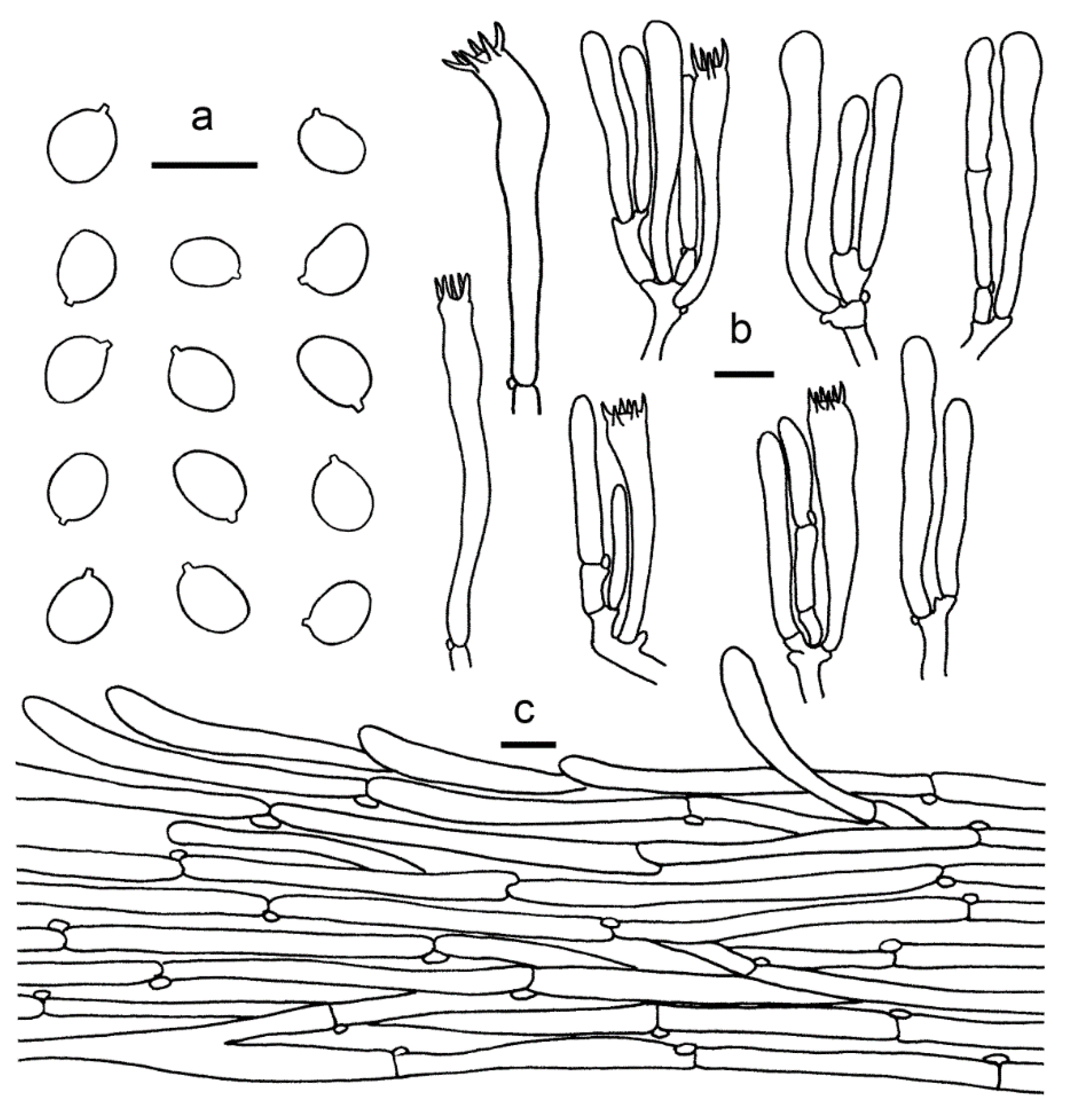
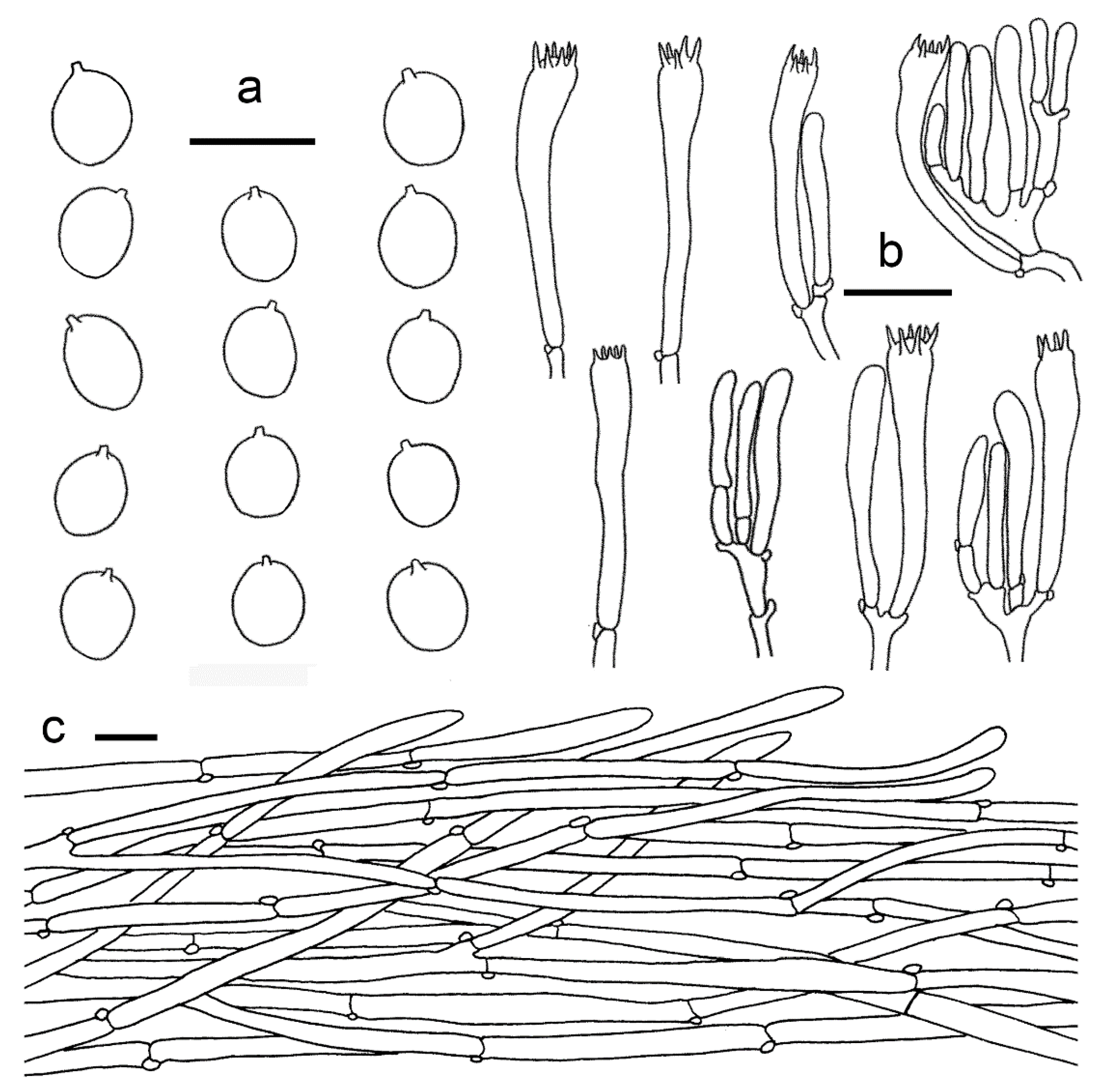
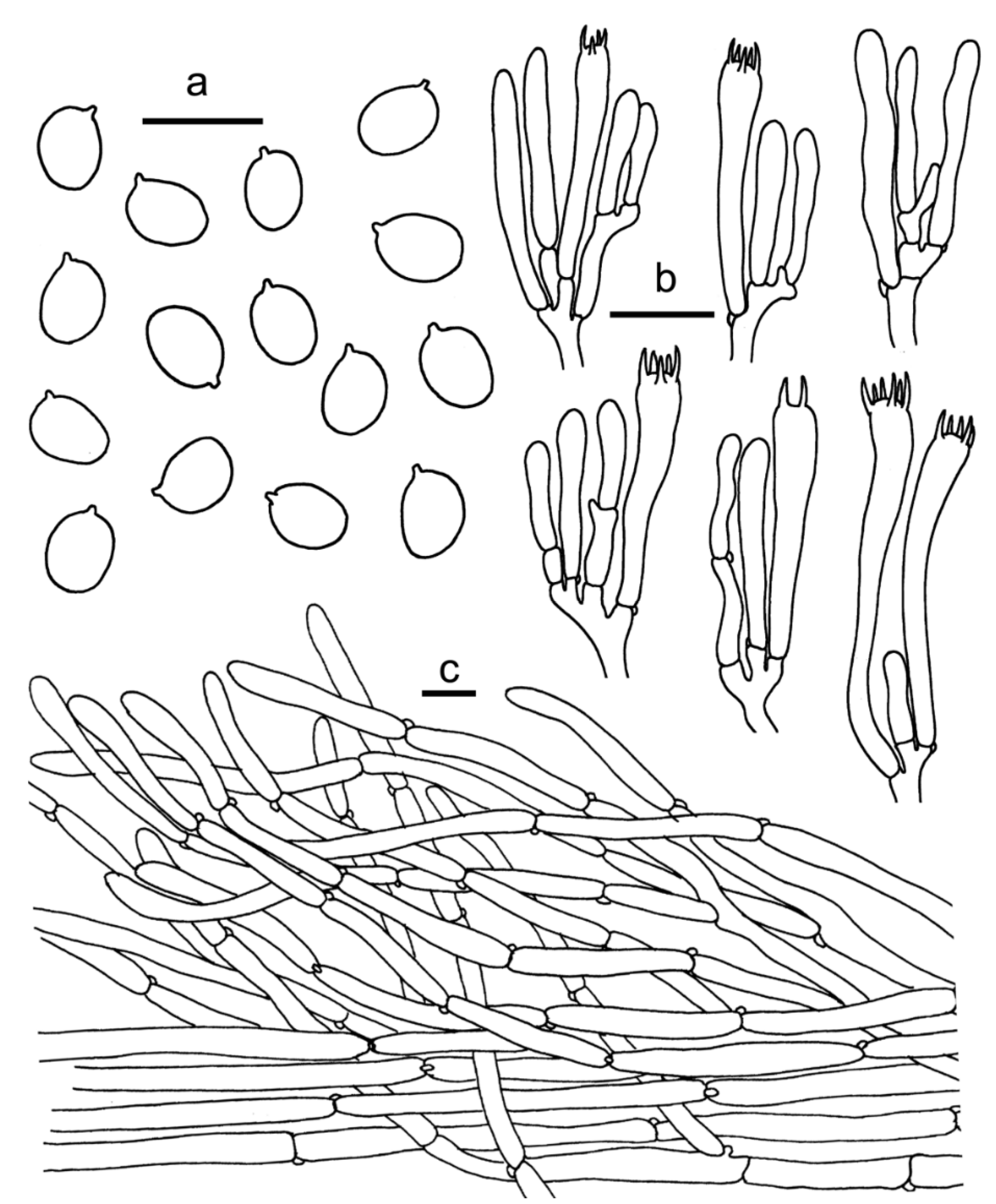
3.2. Taxonomy











3.3. Key to Species of Subgenus Parvocantharellus in China
| 1 Basidiomata white to yellowish-white............................................................................ | 2 |
| 1’ Basidiomata not white, more obviously coloured with yellow to orange tinge...... | 3 |
| 2 Pileus 20–50 mm wide, changing to yellowish when it is bruised; basidiospores 5.5–7.5 × (4) 4.5–6 μm................................................................................................................. | C. albus |
| 2’ Pileus 3–10 mm wide, unchanging in colour when it is bruised; basidiospores 4.5–7 × 4–5.5(–6.2) μm..................................................................................................... | C. minioalbus |
| 3 Pileus relatively larger, usually > 50 mm wide................................... | C. appalachiensis |
| 3’ Pileus smaller, usually < 50 mm wide............................................................................. | 4 |
| 4 Growing under coniferous trees....................................................................................... | 5 |
| 4’ Growing under broadleaf trees....................................................................................... | 8 |
| 5 Basidiospores 6–9 μm long, average length <8 μm...................................................... | 6 |
| 5’ Basidiospores 8–12 μm long, average length >8 μm.................................................... | 7 |
| 6 Pileus pastel yellow, light yellow to greyish-yellow, with a brownish-orange or reddish-brown center, glabrous or tomentosus at centre; basidiospores 6–8 × 4.8–6 μm; Lm × Wm = 7.05(±0.51) × 5.192(±0.34) μm................................................................... | C. austrosinensis |
| 6’ Pileus subtomentosus, greyish-yellow to greyish-orange, usually fading to light yellow to light orange toward the margin; basidiospores (6–)6.5–8.5(–9) × (4.5–)5–6 μm, Lm × Wm = 7.55(±0.61) × 5.56(±0.34) μm................................................................... | C. sinominor |
| 7 Pileus glabrous, bright yellow orange to orange, usually fading to pale orange-buff or pale orange; basidiospores 8–11 × 5–7 μm................................................................. | C. minor |
| 7’ Pileus light brown, brown to brownish-orange, glabrous; basidiospores 8–11(–12) × 5.5–7(–8) μm; distributed in subalpine region, associated with Abies sp.............. | C. zangii |
| 8 Pileus very small, usually < 15 mm wide, greenish-yellow to yellow, well-developed gill-like ridges forked at the margin, basidiospores 6–7.5 × 4.8–5.5 μm.................................................................................................................................. | C.galbanus |
| 8’ Pileus relatively large, usually > 15 mm wide........................................................... | 9 |
| 9 Basidiospores 6–7.5 μm long, average length < 7 μm, pileus yellow to yellowish-orange, hymenophore poorly-developed; basidiospores 6–7(–7.5) × (4.5)4.8–5.5(6) μm, average = 6.79× 5.07 μm............................................................................................... | C. luteovirens |
| 9’ Basidiospores 6–9 μm long, average length > 7 μm.................................................. | 10 |
| 10 Pileus 20–32 mm wide, yellow to orange, hymenophore well-developed, basidiospores 7–8 × 5.2–6.5 μm, average = 7.44 × 6.04 μm...................................................... | C. luteolus |
| 10’ Pileus 15–40 mm wide, light orange, hymenophore well-developed, basidiospores (6.5)7–9 × (4.5)5–6 μm, average = 7.95 × 5.51 μm....................................... | C. aurantinus |
4. Discussion
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Buyck, B.; Kauff, F.; Eyssartier, G.; Couloux, A.; Hofstetter, V. A multilocus phylogeny for worldwide Cantharellus (Cantharellales, Agaricomycetidae). Fungal Divers. 2014, 64, 101–121. [Google Scholar] [CrossRef]
- Jian, S.P.; Dai, R.; Gao, J.U.N.; Feng, B. Cantharellus albus, a striking new species from Southwest China. Phytotaxa 2020, 470, 133–144. [Google Scholar] [CrossRef]
- Corner, E.J.H. A Monograph of Cantharelloid Fungi; Oxford University Press: Oxford, UK, 1966. [Google Scholar]
- Wilson, A.W.; Aime, M.C.; Dierks, J.; Mueller, G.M.; Henkel, T.W. Cantharellaceae of Guyana I: New species, combinations and distribution records of Craterellus and a synopsis of known taxa. Mycologia 2012, 104, 1466–1477. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Buyck, B.; Kauff, F.; Cruaud, C.; Hofstetter, V. Molecular evidence for novel Cantharellus (Cantharellales, Basidiomycota) from tropical African miombo woodland and a key to all tropical African chanterelles. Fungal Divers. 2013, 58, 281–298. [Google Scholar] [CrossRef]
- Buyck, B.; De Crop, E.; Verbeken, A.; Hofstetter, V. Untangling the Central African Cantharellus sect. Tenues: Cantharellus minutissimus sp. nov. and epitypification of Cantharellus alboroseus. Cryptogam. Mycol. 2016, 37, 329–343. [Google Scholar] [CrossRef]
- Buyck, B.; Henkel, T.W.; Dentinger, B.T.M.; Séné, O.; Hofstetter, V. Multigene sequencing provides a suitable epitype, barcode sequences and a precise systematic position for the enigmatic, African Cantharellus miniatescens. Cryptogam. Mycol. 2016, 37, 269–282. [Google Scholar] [CrossRef]
- Buyck, B.; Henkel, T.; Hofstetter, V. The Cantharellus isabellinus species complex (Cantharellales, Hydnaceae) in tropical Africa. Mycosphere 2018, 9, 1209–1221. [Google Scholar] [CrossRef]
- Olariaga, I.; Moreno, G.; Manjón, J.L.; Salcedo, I.; Hofstetter, V.; Rodríguez, D.; Buyck, B. Cantharellus (Cantharellales, Basidiomycota) revisited in Europe through a multigene phylogeny. Fungal Divers. 2017, 83, 263–292. [Google Scholar] [CrossRef]
- Buyck, B.; Hofstetter, V. Cantharellus subg. Pseudocantharellus (Hydnaceae, Cantharellales) revisited: One epityfication, one new synonym and one new species. Mycosphere 2018, 9, 141–148. [Google Scholar] [CrossRef]
- Buyck, B.; Hofstetter, V. The contribution of tef-1 sequences to species delimitation in the Cantharellus cibarius complex in the southeastern USA. Fungal Divers. 2011, 49, 35–46. [Google Scholar] [CrossRef]
- Buyck, B.; Duhem, B.; Das, K.; Jayawardena, R.S.; Niveiro, N.; Pereira, O.L.; Prasher, I.B.; Adhikari, S.; Albertó, E.O.; Bulgakov, T.S.; et al. Fungal Biodiversity Profiles 21–30. Cryptogam. Mycol. 2017, 38, 101–146. [Google Scholar] [CrossRef]
- Foltz, M.J.; Perez, K.E.; Volk, T.J. Molecular phylogeny and morphology reveal three new species of Cantharellus within 20 m of one another in western Wisconsin, USA. Mycologia 2013, 105, 447–461. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Leacock, P.R.; Riddell, J.; Wilson, A.W.; Zhang, R.; Ning, C.; Mueller, G.M. Cantharellus chicagoensis sp. nov. is supported by molecular and morphological analysis as a new yellow chanterelle in midwestern United States. Mycologia 2016, 108, 765–772. [Google Scholar] [CrossRef]
- Eyssartier, G.; Buyck, B. Note nomenclaturale et systématique sur le genre Cantharellus. Doc. Mycol. 2001, 31, 55–56. [Google Scholar]
- Antonín, V.; Hofstetter, V.; Ryoo, R.; Ka, K.H.; Buyck, B. New Cantharellus species from the Republic of Korea. Mycol. Prog. 2017, 16, 75–759. [Google Scholar] [CrossRef]
- Buyck, B. Special issue: Cantharellus. Cryptogam. Mycol. 2016, 37, 255–258. [Google Scholar] [CrossRef]
- Shao, S.C.; Liu, P.G.; Wei, T.Z.; Herrera, M. New insights into the taxonomy of the genus Cantharellus in China: Epityfication of C. yunnanensis W.F. Chiu and the first record of C. cibarius Fr. Cryptogam. Mycol. 2021, 42, 25–37. [Google Scholar] [CrossRef]
- Chiu, W.F. Ten new species of Agaricales from Yunnan, China. Acta Microbiol. Sin. 1973, 13, 129–135. [Google Scholar]
- Zang, M. Some new species of Basidiomycetes from the Xizang autonomous region of China. Acta Microbiol. Sin. 1980, 20, 29–34. [Google Scholar]
- Tian, X.F.; Buyck, B.; Shao, S.C.; Liu, P.G.; Fang, Y. Cantharellus zangii, a new subalpine basidiomycete from southwestern China. Mycotaxon 2012, 120, 99–103. [Google Scholar] [CrossRef]
- Shao, S.C.; Tian, X.F.; Liu, P.G. Cantharellus in southwestern China: A new species and a new record. Mycotaxon 2011, 116, 437–446. [Google Scholar] [CrossRef]
- Shao, S.C.; Buyck, B.; Hofstetter, V.; Tian, X.F.; Geng, Y.H.; Yu, F.Q.; Liu, P.G. Cantharellus hygrophorus, a new species in subgenus Afrocantharellus from tropical southwestern China. Cryptogam. Mycol. 2014, 35, 283–291. [Google Scholar] [CrossRef] [Green Version]
- Shao, S.C.; Buyck, B.; Tian, X.F.; Liu, P.G.; Geng, Y.H. Cantharellus phloginus, a new pink-colored species from southwestern China. Mycoscience 2016, 57, 144–149. [Google Scholar] [CrossRef]
- Shao, S.C.; Liu, P.G.; Tian, X.F.; Buyck, B.; Geng, Y.H. A new species of Cantharellus (Cantharellales, Basidiomycota, Fungi) from subalpine forest in Yunnan, China. Phytotaxa 2016, 252, 273–279. [Google Scholar] [CrossRef] [Green Version]
- An, D.Y.; Liang, Z.Q.; Jiang, S.; Su, M.S.; Zeng, N.K. Cantharellus hainanensis, a new species with a smooth hymenophore from tropical China. Mycoscience 2017, 58, 438–444. [Google Scholar] [CrossRef]
- Zhang, Y.Z.; Liang, Z.Q.; Xie, H.J.; Wu, L.L.; Xue, R.; Zeng, N.K. Cantharellus macrocarpus (Cantharellaceae, Cantharellales), a new species from tropical China. Phytotaxa 2021, 484, 170–180. [Google Scholar] [CrossRef]
- Kornerup, A.; Wanscher, J.H. Taschenlexikon der Farben, 3rd ed.; Muster-Schmidt Verlag: Göttingen, Germany, 1981. [Google Scholar]
- Vilgalys, R.; Hester, M. Rapid genetic identification and mapping of enzymatically amplified ribosomal DNA from several Cryptococcus species. J. Bacteriol. 1990, 172, 4238–4246. [Google Scholar] [CrossRef] [Green Version]
- Morehouse, E.A.; James, T.Y.; Ganley, A.R.D.; Vilgalys, R.; Berger, L.; Murphy, P.J.; Longcore, J.E. Multilocus sequence typing suggests the chytrid pathogen of amphibians is a recently emerged clone. Mol. Ecol. 2003, 12, 395–403. [Google Scholar] [CrossRef]
- Katoh, K.; Rozewicki, J.; Yamada, K.D. MAFFT online service: Multiple sequence alignment, interactive sequence choice and visualization. Brief Bioinform. 2019, 20, 1160–1166. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hall, T.A. BioEdit: A user-friendly biological sequence alignment editor and analysis program for Windows 95/98/NT. Nucleic Acids Symp. Ser. 1999, 41, 95–98. [Google Scholar] [CrossRef]
- Buyck, B.; Hofstetter, V.; Ryoo, R.; Ka, K.-H.; Antonín, V. New Cantharellus species from South Korea. MycoKeys 2020, 76, 31–47. [Google Scholar] [CrossRef] [PubMed]
- Buyck, B.; Kauff, F.; Randrianjohany, E.; Hofstetter, V. Sequence data reveal a high diversity of Cantharellus associated with endemic vegetation in Madagascar. Fungal Divers. 2015, 70, 189–208. [Google Scholar] [CrossRef]
- Moncalvo, J.M.; Nilsson, R.H.; Koster, B.; Dunham, S.M.; Bernauer, T.; Matheny, P.B.; McLenon, T.; Margaritescu, S.; Weiß, M.; Garnica, S.; et al. The cantharelloid clade: Dealing with incongruent gene trees and phylogenetic reconstruction methods. Mycologia 2006, 98, 937–948. [Google Scholar] [CrossRef]
- Suhara, H.; Kurogi, S. Cantharellus cyphelloides (Cantharellales), a new and unusual species from a Japanese evergreen broad-leaved forest. Mycol. Prog. 2015, 14, 55. [Google Scholar] [CrossRef]
- Montoya, L.; Herrera, M.; Bandala, V.M.; Ramos, A. Two new species and a new record of yellow Cantharellus from tropical Quercus forests in eastern Mexico with the proposal of a new name for the replacement of Craterellus confluens. MycoKeys 2021, 80, 91–114. [Google Scholar] [CrossRef] [PubMed]
- Deepika, K.; Upadhyay, R.C.; Reddy, M.S. New records of Cantharellus species from the northwestern Himalayas of India. Mycology 2013, 4, 205–220. [Google Scholar] [CrossRef]
- Das, K.; Hofstetter, V.; Chakraborty, D.; Baghela, A.; Singh, S.K.; Buyck, B. Cantharellus sikkimensis sp. nov. (Cantharellales, Agaricomycetes) from the Indian Himalayas. Phytotaxa 2015, 222, 267–275. [Google Scholar] [CrossRef]
- Buyck, B.; Cruaud, C.; Couloux, A.; Hofstetter, V. Cantharellus texensis sp. nov. from Texas, a Southern lookalike of C. cinnabarinus revealed by tef-1 sequence data. Mycologia 2011, 103, 1037–1046. [Google Scholar] [CrossRef]
- Matheny, P.B.; Wang, Z.; Binder, M.; Curtis, J.M.; Lim, Y.W.; Nilsson, R.H.; Hughes, K.W.; Hofstetter, V.; Ammirati, J.F.; Schoch, C.L.; et al. Contributions of rpb2 and tef1 to the phylogeny of mushrooms and allies (Basidiomycota, Fungi). Mol. Phylogenetics Evol. 2007, 43, 430–451. [Google Scholar] [CrossRef]
- Zhang, M.; Li, T.H.; Wang, C.Q.; Zeng, N.K.; Deng, W.Q. Phylogenetic overview of Aureoboletus (Boletaceae, Boletales), with descriptions of six new species from China. Mycokeys 2019, 61, 111–145. [Google Scholar] [CrossRef]
- Stamatakis, A. RAxML-VI-HPC: Maximum likelihood-based phylogenetic analyses with thousands of taxa and mixed models. Bioinformatics 2006, 22, 2688–2690. [Google Scholar] [CrossRef] [PubMed]
- Ronquist, F.; Huelsenbeck, J.P. MrBayes 3: Bayesian phylogenetic inference under mixed models. Bioinformatics 2003, 19, 1572–1574. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nylander, J. MrModeltest 2.3. Computer Program and Documentation Distributed by the Author; Evolutionary Biology Centre, Uppsala University: Uppsala, Sweden, 2004. [Google Scholar]
- Kesel, A.D.; Amalfi, M.; Kasongo Wa Ngoy, B.; Yorou, N.S.; Raspé, O.; Degreef, J.; Buyck, B. New and interesting Cantharellus from tropical Africa. Cryptogam. Mycol. 2016, 37, 283–327. [Google Scholar] [CrossRef]
- Shao, S.C.; Tian, X.F.; Liu, P.G. Two species with intercontinental disjunct distribution of the genus Cantharellus. J. Yunnan Agric. Univ. 2012, 27, 150–155. [Google Scholar]
- Ryvarden, L.; Petersen, R. Notes on cantharelloid fungi IV. Two new species of Cantharellus. Sven. Bot. Tidskr. 1971, 65, 399–405. [Google Scholar]
- Feibelman, T.P.; Bennett, J.W.; Cibula, W.G. Cantharellus tabernensis: A new species from the Southeastern United States. Mycologia 1996, 88, 295–301. [Google Scholar] [CrossRef]
- Buyck, B.; Lewis, D.P.; Eyssartier, G.; Hofstetter, V. Cantharellus quercophilus sp. nov. and its comparison to other small, yellow or brown American chanterelles. Cryptogam. Mycol. 2010, 31, 17–33. [Google Scholar]
- Peck, C.H. Report of the Botanist. Annual Rep. N. Y. State Mus. 1872, 23, 27–136. [Google Scholar]
- Bigelow, H.E. The cantharelloid fungi of New England and adjacent areas. Mycologia 1978, 70, 707–756. [Google Scholar] [CrossRef]
- Kuo, M. Cantharellus minor. Retrieved from the MushroomExpert.Com Web Site 2015. Available online: http://www.mushroomexpert.com/cantharellus_minor.html (accessed on 1 March 2015).
- Li, Y.; Li, T.H.; Yang, Z.L.; Bau, T.; Dai, Y.C. Atlas of Chinese Macrofungal Resources; Central China Farmers Publishing House: Zhengzhou, China, 2015.
- Zhao, G.P.; Hu, J.J.; Li, Y.; Zhang, B. A review of the studies of the genus Cantharellus. Microbiol. China 2020, 48, 1260–1272. [Google Scholar] [CrossRef]
- Yang, Z.L.; Wu, G.; Li, Y.C.; Wang, X.H.; Cai, Q. Common Edible and Poisonous Mushrooms of Southwestern China; Science Press: Beijing, China, 2021. [Google Scholar]
- Tian, X.F.; Liu, P.G.; Shao, S.C. Research status and prospect of the genus Cantharellus Adans. ex Fr. (Cantharellaceae, Basidiomycota). Microbiology 2009, 36, 1577–1586. [Google Scholar] [CrossRef]




{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Taxa | Voucher | Locality | GenBank Accession No. | Reference | ||
|---|---|---|---|---|---|---|
| LSU | tef1 | rpb2 | ||||
| Cantharellus afrocibarius | BB 96.236 | Zambia | KF294669 | JX192994 | KF294747 | [5] |
| C. afrocibarius | BB 96.235 | Zambia | KF294668 | JX192993 | KF294746 | [5] |
| C. albidolutescens | BB 08.070 | Madagascar | KF294646 | JX192982 | KF294723 | [5] |
| C. albidosquamosus | PC0142511 | Cameroon | MT002285 | MT002270 | MT004806 | [33] |
| C. alborufescens | AH44223 | Spain | KR677531 | KX828816 | KX828735 | [34] |
| C. albus | HKAS107047 | China | MT782542 | MT776017 | MT776014 | [2] |
| C. albus | HKAS107048 | China | MT782541 | MT776016 | MT776013 | [2] |
| C. albus | HKAS107045 | China | MT782540 | MT776015 | MT776012 | [2] |
| C. albus | GDGM56646 | China | MZ605071 | MZ613974 | MZ614019 | Present study |
| C. albus | GDGM81399 | China | MZ605074 | MZ613977 | MZ614022 | Present study |
| C. albus | GDGM81064 | China | MZ605073 | MZ613976 | MZ614021 | Present study |
| C. albus | GDGM77819 | China | MZ605072 | MZ613975 | MZ614020 | Present study |
| C. altipes | BB 07.019 | USA | KF294627 | GQ914939 | KF294702 | [1] |
| C. appalachiensis | GRSM77088 | USA | DQ898690 | – | DQ898748 | [35] |
| C. appalachiensis | BB 07.123 | USA | KF294635 | GQ914979 | KF294711 | [1] |
| C. aurantinus | GDGM46278 | China | MZ766517 | MZ766560 | Present study | |
| C. aurantinus | GDGM46279 | China | MZ766518 | MZ766561 | MZ766571 | Present study |
| C. aurantinus | GDGM84974 | China | MZ766521 | MZ766564 | MZ766572 | Present study |
| C. aurantinus | GDGM84978 | China | MZ766522 | MZ766565 | Present study | |
| C. aurantinus | GDGM81889 | China | MZ766519 | MZ766562 | MZ766574 | Present study |
| C. aurantinus | GDGM81899 | China | MZ766520 | MZ766563 | MZ766573 | Present study |
| C. austrosinensis | GDGM60305 | China | MZ605077 | MZ613980 | MZ614023 | Present study |
| C. austrosinensis | GDGM79507 | China | MZ605078 | MZ613981 | MZ614024 | Present study |
| C. austrosinensis | GDGM81303 | China | MZ605084 | MZ613986 | MZ614029 | Present study |
| C. austrosinensis | GDGM81249 | China | MZ605082 | MZ613983 | MZ614027 | Present study |
| C. austrosinensis | GDGM80616 | China | MZ605081 | MZ613982 | MZ614026 | Present study |
| C. austrosinensis | GDGM80211 | China | MZ605080 | MZ613984 | MZ614025 | Present study |
| C. austrosinensis | GDGM81381 | China | MZ605086 | MZ613988 | MZ614031 | Present study |
| C. austrosinensis | GDGM81379 | China | MZ605085 | MZ613987 | MZ614030 | Present study |
| C. austrosinensis | GDGM81271 | China | MZ605083 | MZ613985 | MZ614028 | Present study |
| C. austrosinensis | GDGM82877 | China | MZ605088 | MZ613990 | MZ614033 | Present study |
| C. austrosinensis | GDGM80151 | China | MZ605079 | – | – | Present study |
| C. austrosinensis | GDGM81985 | China | MZ605087 | MZ613989 | MZ614032 | Present study |
| C. avellaneus | 1217/ER | Madagascar | KX857093 | – | KX856997 | [17] |
| C. cerinoalbus | AV 06.051 | Malaysia | KF294663 | – | KF294741 | [1] |
| C. cibarius | GE 07.025 | France | KF294658 | GQ914949 | KF294736 | [1] |
| C. cibarius | BB 07.300 | Slovakia | KF294641 | GQ914950 | KF294718 | [1] |
| C. cinnabarinus | BB 07.053 | USA | KF294630 | GQ914984 | KF294705 | [1] |
| C. cinnabarinus | BB 07.001 | USA | KF294624 | GQ914985 | KF294698 | [1] |
| C. congolensis | 1645/BB16.044 | Saharan Africa | KX857102 | KX857075 | KX857006 | [17] |
| C. congolensis | 1676/BB16.123 | Saharan Africa | KX857106 | KX857078 | KX857010 | [17] |
| C. aff. congolensis | BB 06.176 | Madagascar | KF294606 | – | KF294680 | [1] |
| C. aff. congolensis | BB 06.197 | Madagascar | KF294608 | – | KF294683 | [1] |
| C. curvatus | BRNM:825749 | South Korea | MW124390 | – | [33] | |
| C. cyphelloides | TNS F-61721 | Japan | NG059027 | – | – | [36] |
| C. decolorans | BB 08.278 | Madagascar | KF294654 | GQ914968 | KF294731 | [1] |
| C. galbanus | GDGM86249 | China | ZM766516 | MZ766568 | MZ766577 | Present study |
| C. guyanensis | 1517/MR | Guyane | KX857095 | KX857061 | KX856999 | [17] |
| C. guyanensis | 1501/MRG07 | Guyane | KX857094 | KX857060 | KX856998 | [17] |
| C. parvoflavus | Montoya 5423 | Mexico | MT371337 | MT449706 | – | [37] |
| C. parvoflavus | Herrera 204 | Mexico | MT371338 | MT449707 | – | [37] |
| C. parvoflavus | Herrera 229 | Mexico | MT371339 | MT449708 | – | [37] |
| C. himalayensis | DK-2010b | India | HM750917 | – | – | [38] |
| C. hygrophoroides | HKAS80614 | China | KJ004002 | KJ004003 | – | [23] |
| C. ibityi | BB 08.196 | Madagascar | KF294650 | GQ914980 | KF294727 | [1] |
| C. koreanus | 1697/V. Antonin 14.115 | Republic of Korea | – | KY271940 | – | [17] |
| C. koreanus | 1689/V. Antonin 13.136 | Republic of Korea | – | KY271941 | – | [26] |
| C. lateritius | BB 07.025 | USA | KF294628 | GQ914957 | KF294703 | [1] |
| C. luteolus | GDGM44258 | China | ZM766514 | MZ766566 | MZ766570 | Present study |
| C. luteolus | GDGM60393 | China | ZM766515 | MZ766566 | MZ766575 | Present study |
| C. luteolus | GDGM86247 | China | MZ766513 | MZ766567 | MZ766576 | Present study |
| C. luteovirens | GDGM45899 | China | MZ605095 | – | – | Present study |
| C. luteovirens | GDGM80296 | China | MZ605089 | MZ613991 | MZ614034 | Present study |
| C. luteovirens | GDGM81395 | China | MZ605093 | MZ613995 | MZ614037 | Present study |
| C. luteovirens | GDGM81079 | China | MZ605092 | MZ613994 | MZ614036 | Present study |
| C. luteovirens | GDGM80672 | China | MZ605090 | MZ613992 | MZ614035 | Present study |
| C. luteovirens | GDGM80680 | China | MZ605091 | MZ613993 | – | Present study |
| C. minioalbus | GDGM78910 | China | MZ605098 | MZ613999 | MZ614043 | Present study |
| C. minioalbus | GDGM78934 | China | MZ605102 | MZ614003 | MZ614047 | Present study |
| C. minioalbus | GDGM78883 | China | MZ605096 | MZ613997 | MZ614041 | Present study |
| C. minioalbus | GDGM78901 | China | MZ605097 | MZ613998 | MZ614042 | Present study |
| C. minioalbus | GDGM78916 | China | MZ605100 | MZ614001 | MZ614045 | Present study |
| C. minioalbus | GDGM78915 | China | MZ605099 | MZ614000 | MZ614044 | Present study |
| C. minioalbus | GDGM78926 | China | MZ605101 | MZ614002 | MZ614046 | Present study |
| C. minioalbus | GDGM78955 | China | MZ605103 | – | – | Present study |
| C. minioalbus | GDGM78997 | China | MZ605104 | – | – | Present study |
| C. minor | BB 07.057 | USA | KF294632 | JX192979 | KF294707 | [1] |
| C. minor | BB 07.002 | USA | KF294625 | JX192978 | KF294699 | [1] |
| C. pallens | AH39124 | Morocco | KX828804 | KX828834 | KX828755 | [9] |
| C. platyphyllus | BB 98.126 | Tanzania | KF294620 | JX192975 | KF294694 | [1] |
| C. pseudominimus | JV 00.663 | Portugal | KF294657 | JX192991 | KF294735 | [1,5] |
| C. rhodophyllus | BB 16.126 | Congo | MK422958 | MG450695 | – | [10] |
| C. rhodophyllus | BB 16.006 | Congo | MK422957 | MG450696 | – | [10] |
| C. romagnesianus | AH44218 | Spain | KX828807 | KX828836 | KX828757 | [9] |
| C. roseofagetorum | AH44789 | Georgia | KX828812 | KX828839 | KX828760 | [9] |
| C. sebosus | BB 08.234 | Madagascar | KF294652 | JX192986 | KF294729 | [1] |
| C. sikkimensis | AB-2015 | India | KP938966 | – | – | [39] |
| C. sinominor | GDGM80791 | China | MZ605106 | MZ614005 | MZ614049 | Present study |
| C. sinominor | GDGM80788 | China | MZ605105 | MZ614004 | MZ614048 | Present study |
| C. sinominor | GDGM80842 | China | MZ605107 | MZ614006 | MZ614050 | Present study |
| C. sinominor | GDGM80885 | China | MZ605108 | MZ614007 | MZ614051 | Present study |
| C. splendens | BB 96.306 | Zambia | KF294670 | – | KF294748 | [1] |
| C. subalbidus | OSC81782 | USA | KX828814 | KX828841 | KX828762 | [9] |
| C. aff. subcyanoxanthus | BB 98.014 | Tanzania | KF294615 | JX192973 | KF294689 | [1] |
| C. subincarnatus subsp. rubrosalmoneus | BB 06.080 | Madagascar | KF294602 | JX192963 | KF294676 | [1] |
| C. symoensii | BB 98.113 | Tanzania | KF294619 | JX192974 | KF294693 | [1] |
| C. tabernensis | BB 07.119 | USA | KF294634 | GQ914976 | KF294709 | [1] |
| C. tabernensis | BB 07.056 | USA | KF294631 | GQ914974 | KF294706 | [1] |
| C. tenuithrix | BB 07.125 | USA | JN940600 | GQ914947 | KF294712 | [1,40] |
| C. texensis | 341/O7.120 | USA | JN940601 | GQ914987 | KF294710 | [1,40] |
| C. texensis | BB 07.018 | USA | KF294626 | GQ914988 | KF294701 | [1] |
| C. tomentosus | BB 98.060 | Tanzania | KF294672 | JX192995 | KF294750 | [1] |
| C. tricolor | BB 06.180 | Madagascar | JN940604 | JX192969 | KF294682 | [1,7] |
| C. zangii | GDGM83171 | China | MZ605113 | MZ614012 | MZ614056 | Present study |
| C. zangii | GDGM83173 | China | MZ605114 | MZ614013 | MZ614057 | Present study |
| C. zangii | GDGM83186 | China | MZ605117 | MZ614016 | MZ614060 | Present study |
| C. zangii | GDGM82399 | China | MZ605112 | MZ614011 | MZ614055 | Present study |
| C. zangii | GDGM82389 | China | MZ605110 | MZ614009 | MZ614053 | Present study |
| C. zangii | GDGM83176 | China | MZ605115 | MZ614014 | MZ614058 | Present study |
| C. zangii | GDGM83181 | China | MZ605116 | MZ614015 | MZ614059 | Present study |
| C. zangii | GDGM82393 | China | MZ605111 | MZ614010 | MZ614054 | Present study |
| C. zangii | GDGM82374 | China | MZ605109 | MZ614008 | MZ614052 | Present study |
| C. zangii | GDGM83193 | China | MZ605118 | MZ614017 | MZ614061 | Present study |
| C. zangii | GDGM83228 | China | MZ605119 | MZ614018 | MZ614062 | Present study |
| Craterellus cornucopioides | AFTOL-ID 286 | USA | AY700188 | – | DQ366287 | [41] |
| Cr. tubaeformis | BB 07.293 | Slovakia | KF294640 | GQ914989 | KF294717 | [1,11] |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zhang, M.; Wang, C.-Q.; Buyck, B.; Deng, W.-Q.; Li, T.-H. Multigene Phylogeny and Morphology Reveal Unexpectedly High Number of New Species of Cantharellus Subgenus Parvocantharellus (Hydnaceae, Cantharellales) in China. J. Fungi 2021, 7, 919. https://0-doi-org.brum.beds.ac.uk/10.3390/jof7110919
Zhang M, Wang C-Q, Buyck B, Deng W-Q, Li T-H. Multigene Phylogeny and Morphology Reveal Unexpectedly High Number of New Species of Cantharellus Subgenus Parvocantharellus (Hydnaceae, Cantharellales) in China. Journal of Fungi. 2021; 7(11):919. https://0-doi-org.brum.beds.ac.uk/10.3390/jof7110919
Chicago/Turabian StyleZhang, Ming, Chao-Qun Wang, Bart Buyck, Wang-Qiu Deng, and Tai-Hui Li. 2021. "Multigene Phylogeny and Morphology Reveal Unexpectedly High Number of New Species of Cantharellus Subgenus Parvocantharellus (Hydnaceae, Cantharellales) in China" Journal of Fungi 7, no. 11: 919. https://0-doi-org.brum.beds.ac.uk/10.3390/jof7110919