
Pineapple Mycobiome Related to Fruitlet Core Rot Occurrence and the Influence of Fungal Species Dispersion Patterns

 , ,
, ,  , , and
, , and
Abstract
:1. Introduction
2. Materials and Methods
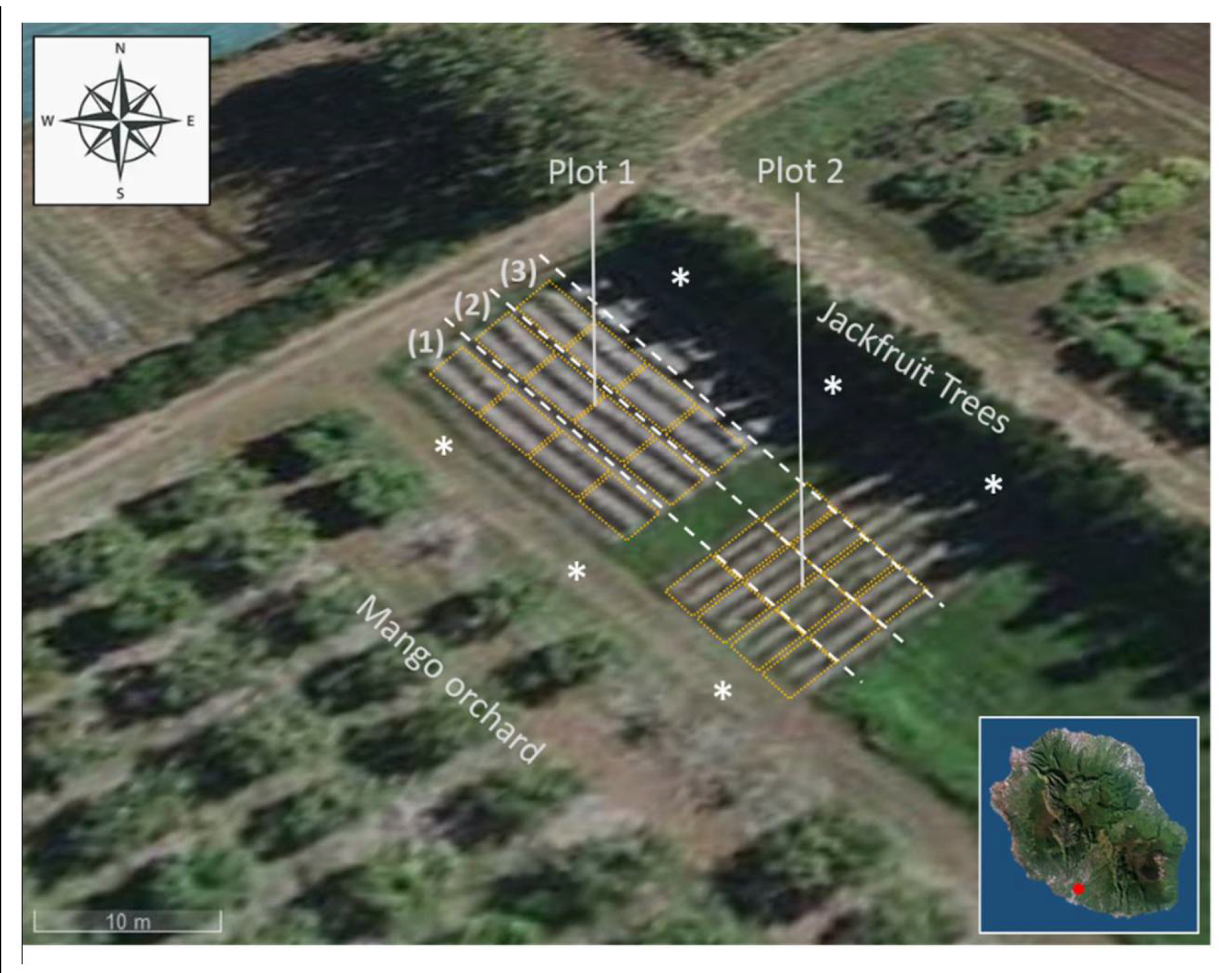
2.1. Plant Material and Agricultural Practices
2.2. FCR Symptom Occurrence and Fruitlets Sampling
2.3. Isolation of Cultivatable Fungal Flora from Fruitlet and Soil Samples
2.3.1. Fruitlet Tissues
2.3.2. Soil
2.4. Molecular Characterization of Cultivatable Fungal Flora from Fruitlets and Soil Samples

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Sequence ID of Primers Pair | Target Locus | Sequence Forward (5′→3′) | Sequence Reverse (5′→3′) | Product Length (bp) | References |
|---|---|---|---|---|---|
| ef1/ef2 | Translation Elongation Factor-1α | ATGGGTAAGGAAGACAAGAC | GGAAGTACCAGTGATCATGTT | 380–680 | [31,32,33] |
| Bt2a/Bt2b | β-tubulin | GGTAACCAAATCGGTGCTGCTTTC | ACCCTCAGTGTAGTGACCCTTGGC | 250–500 | [34] |
| ITS1F/ITS2 | Internal Transcribed Spacer 1 | CTTGGTCATTTAGAGGAAGTAA | GCTGCGTTCTTCATCGATGC | 145–695 | [35,36] |
| ITS1F/ITS4 | Entire Internal Transcribed Spacer | TCCTCCGCTTATTGATATGC | 600–800 | ||
| GC-ITS1F/ITS4 | CGCCCGCCGCGCGCGGCGGGCGGGGCGGGGGCACGGGGGGCTTGGTCATTTAGAGGAAGTAA | TCCTCCGCTTATTGATATGC |
2.5. Koch’s Postulates
2.5.1. Controlled Inoculations and Plant Material
2.5.2. Fruit Sampling
2.5.3. DNA Extraction
2.5.4. PCR-Denaturing Gradient Gel Electrophoresis (DGGE)
2.6. Computational Analysis
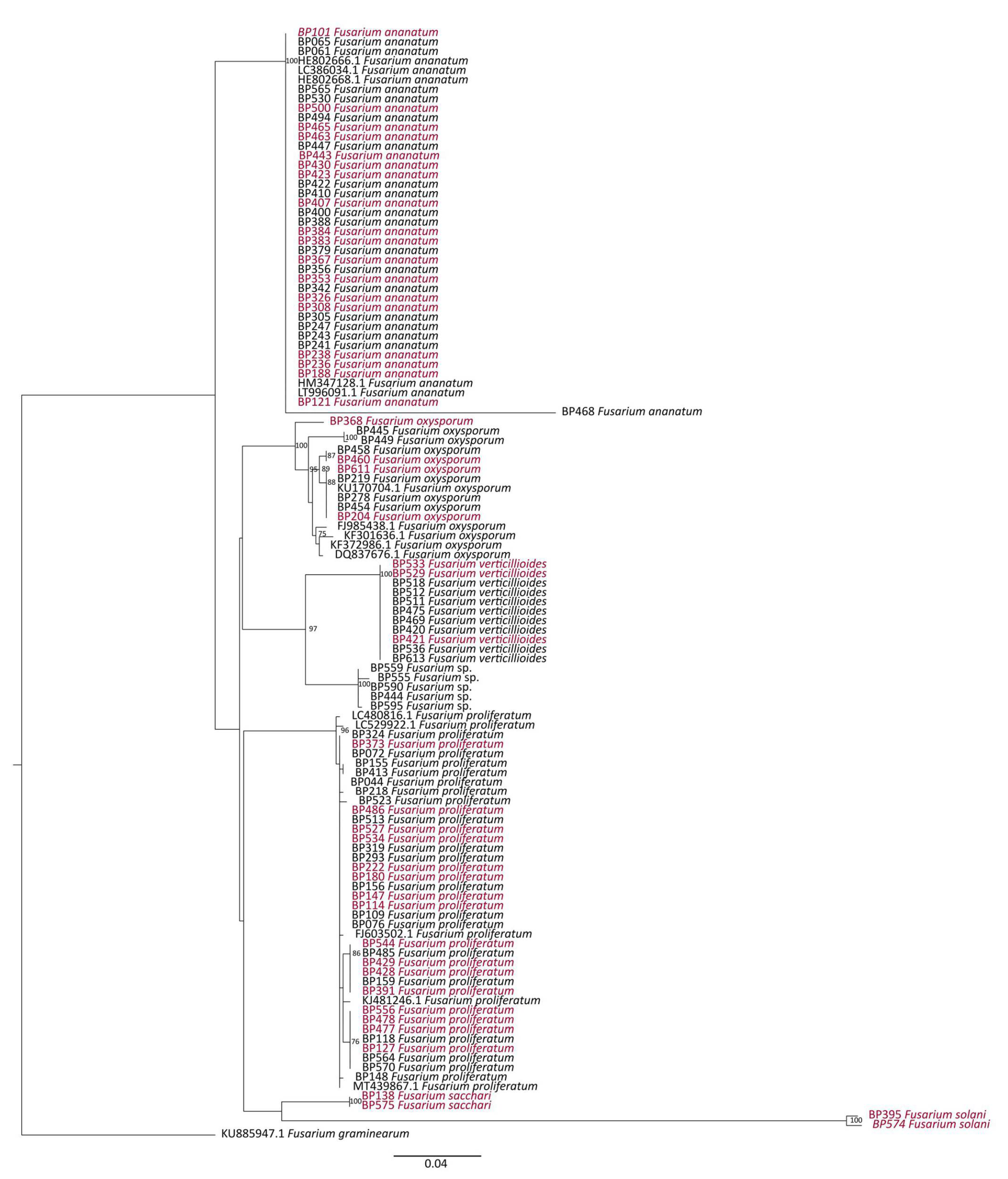
2.6.1. Phylogenetic Analysis
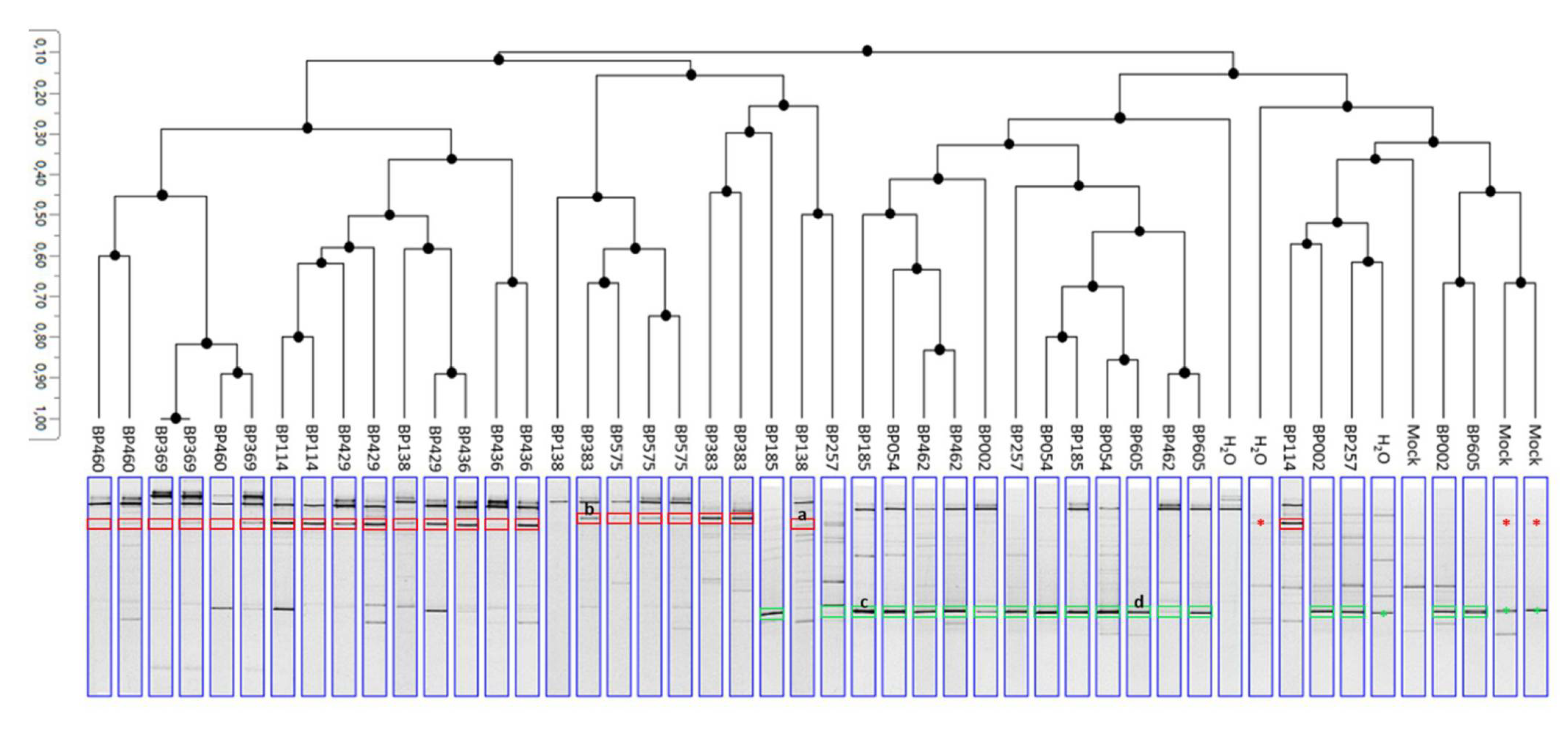
2.6.2. DGGE Band Pattern
2.6.3. Statistical Analysis
3. Results
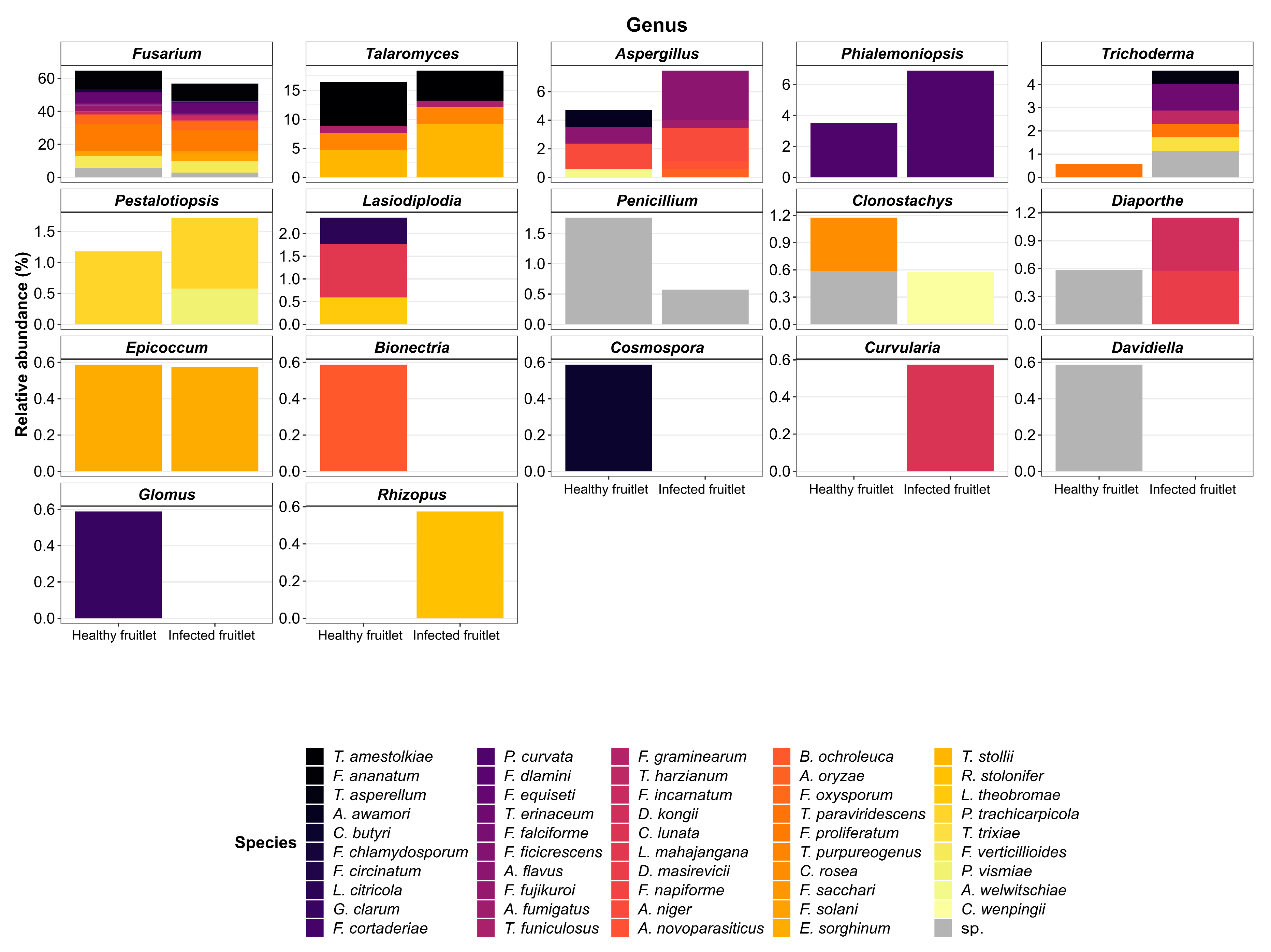
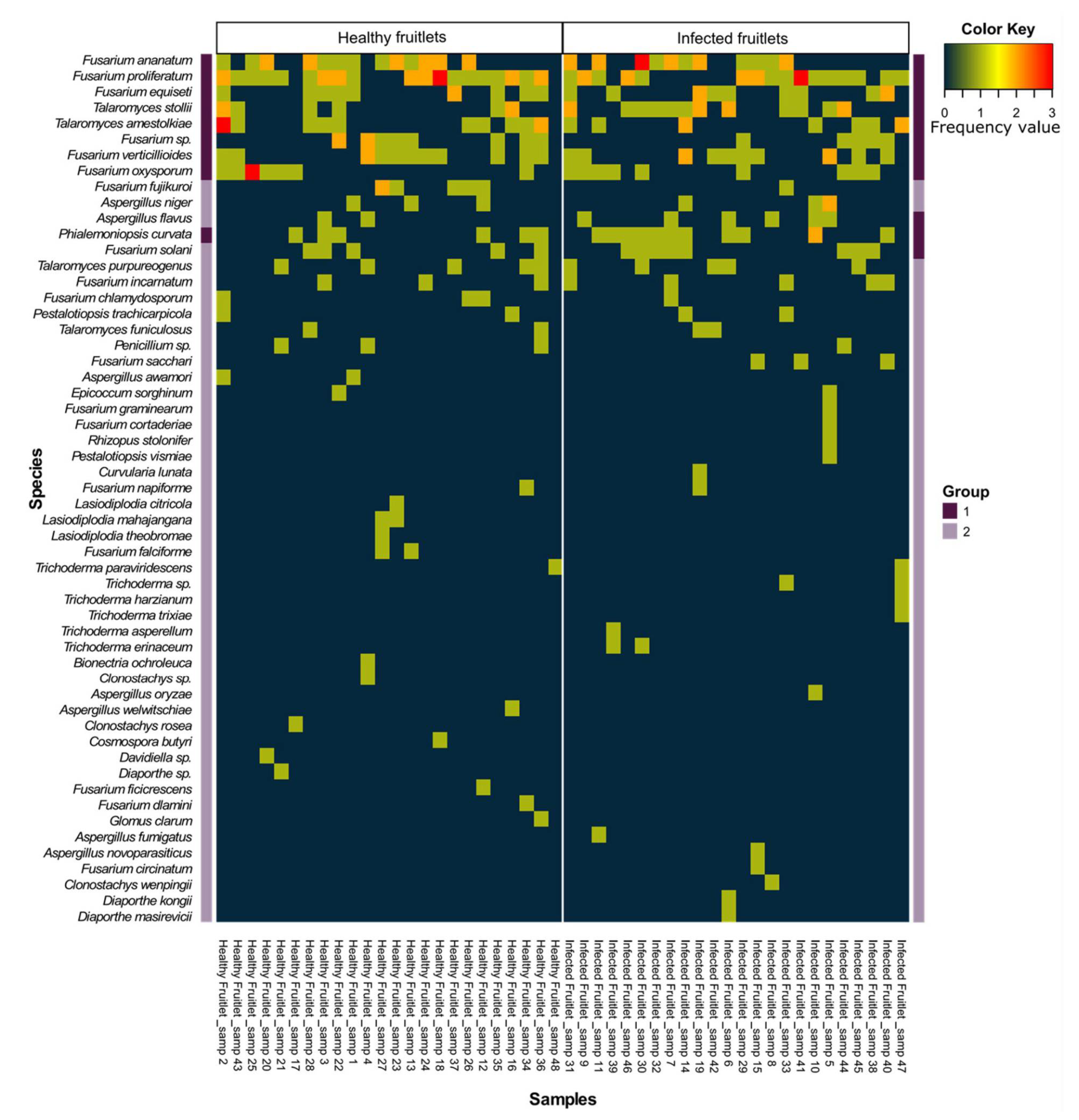
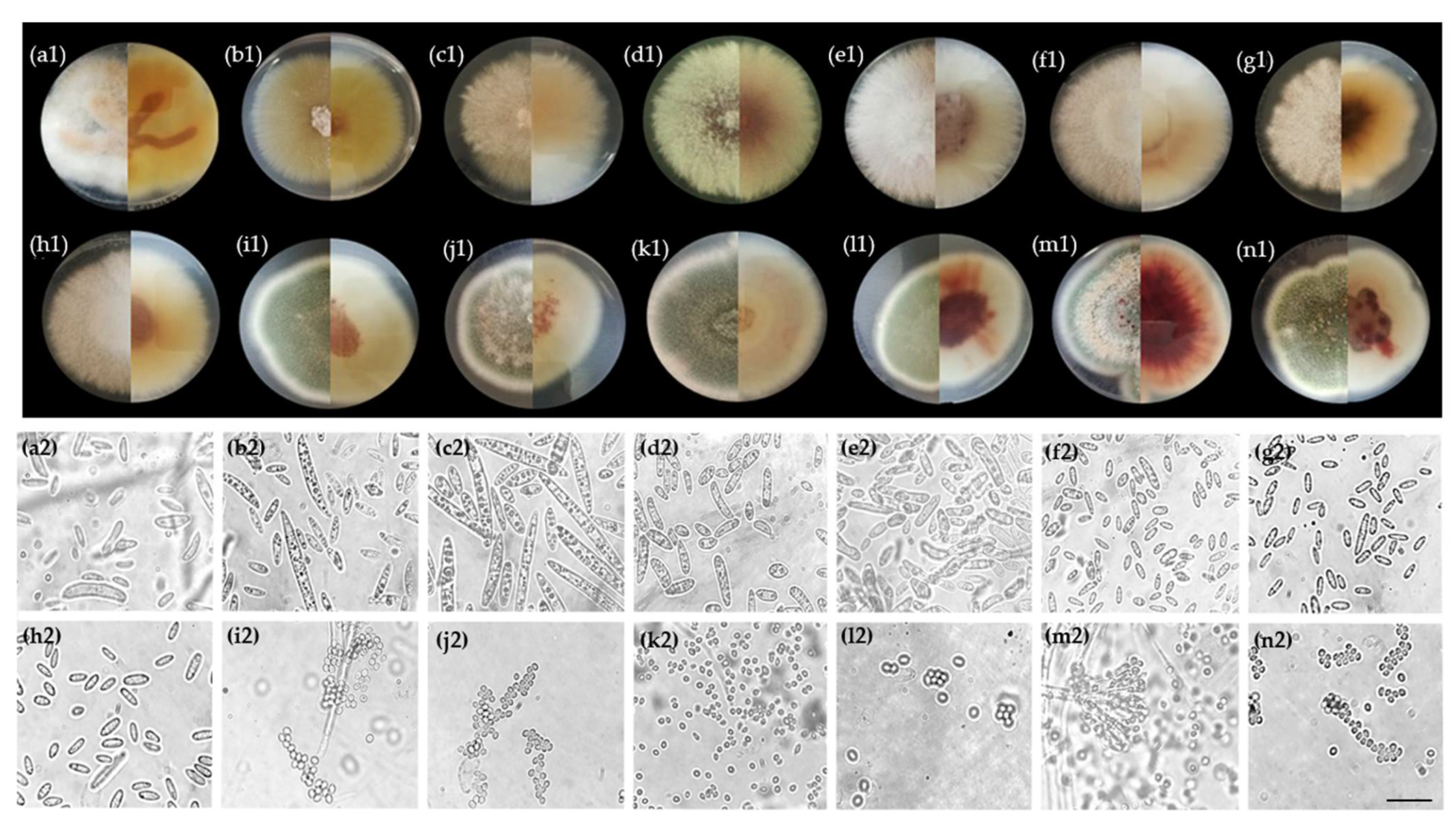
3.1. Fungal Flora of Healthy and Naturally Infected Fruitlets
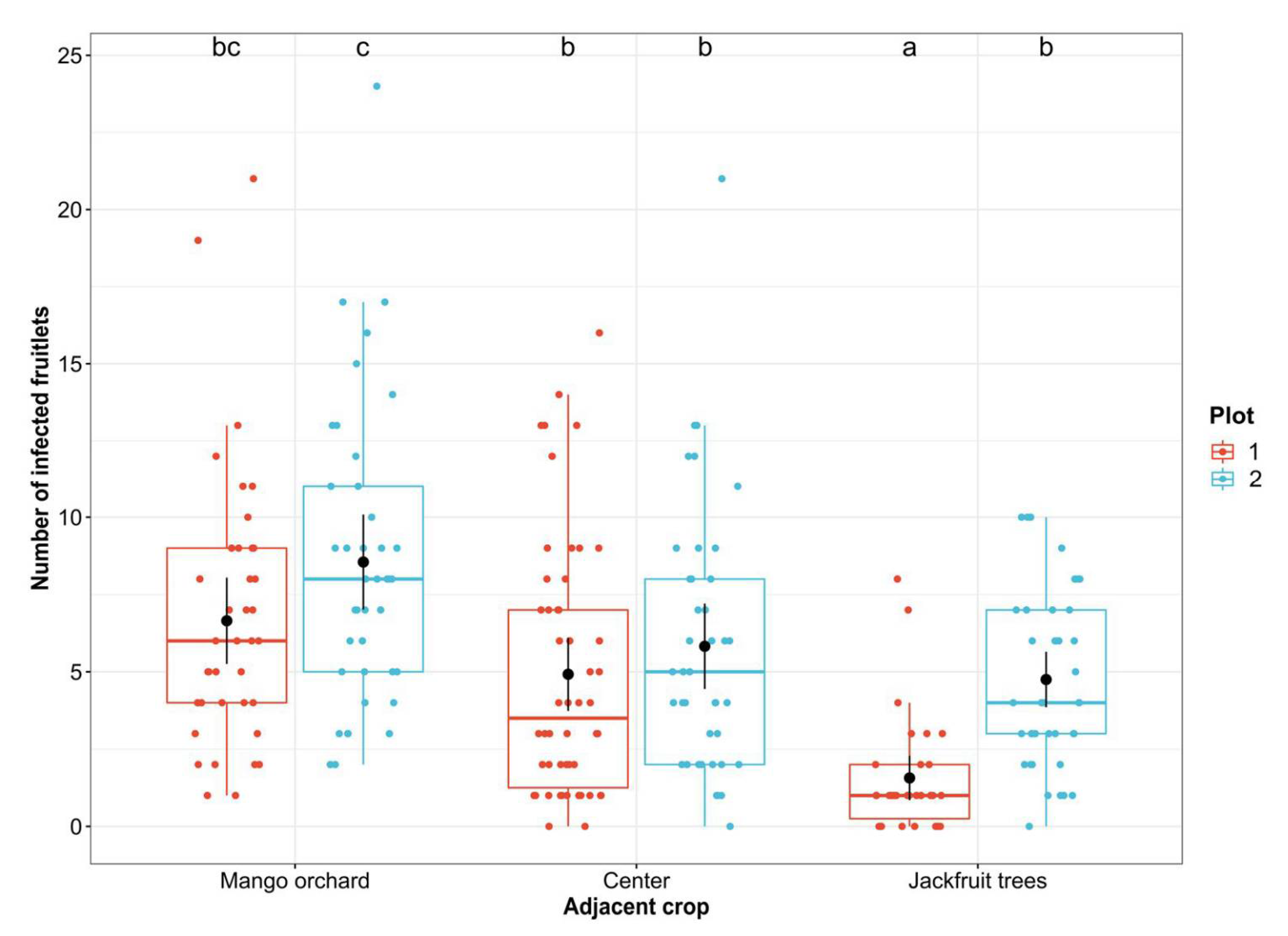
3.2. Correlation between the Diversity of Fruitlet Mycobiomes and FCR Incidence
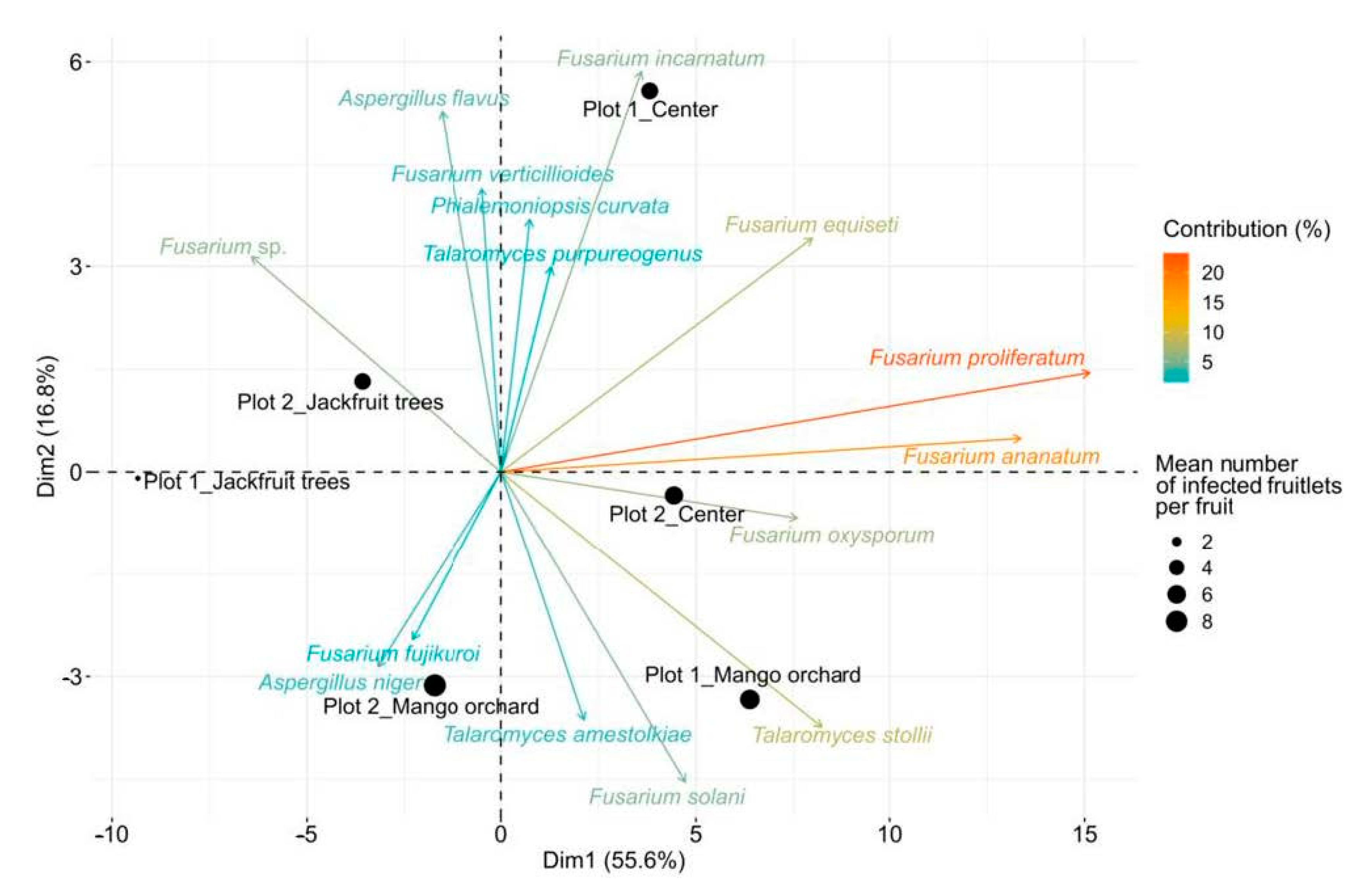
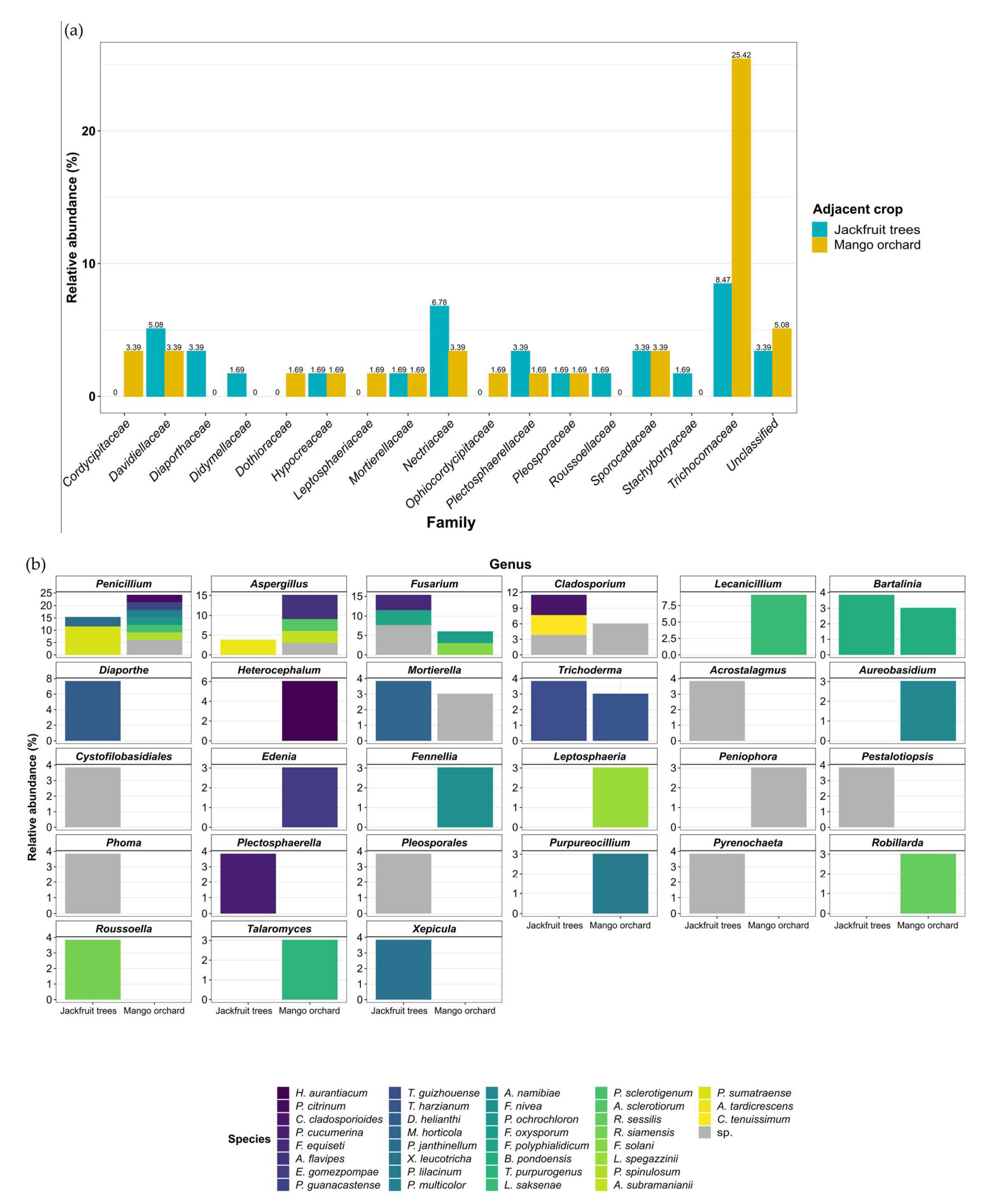
3.3. Potential Sources of Inoculum and Dispersion Patterns
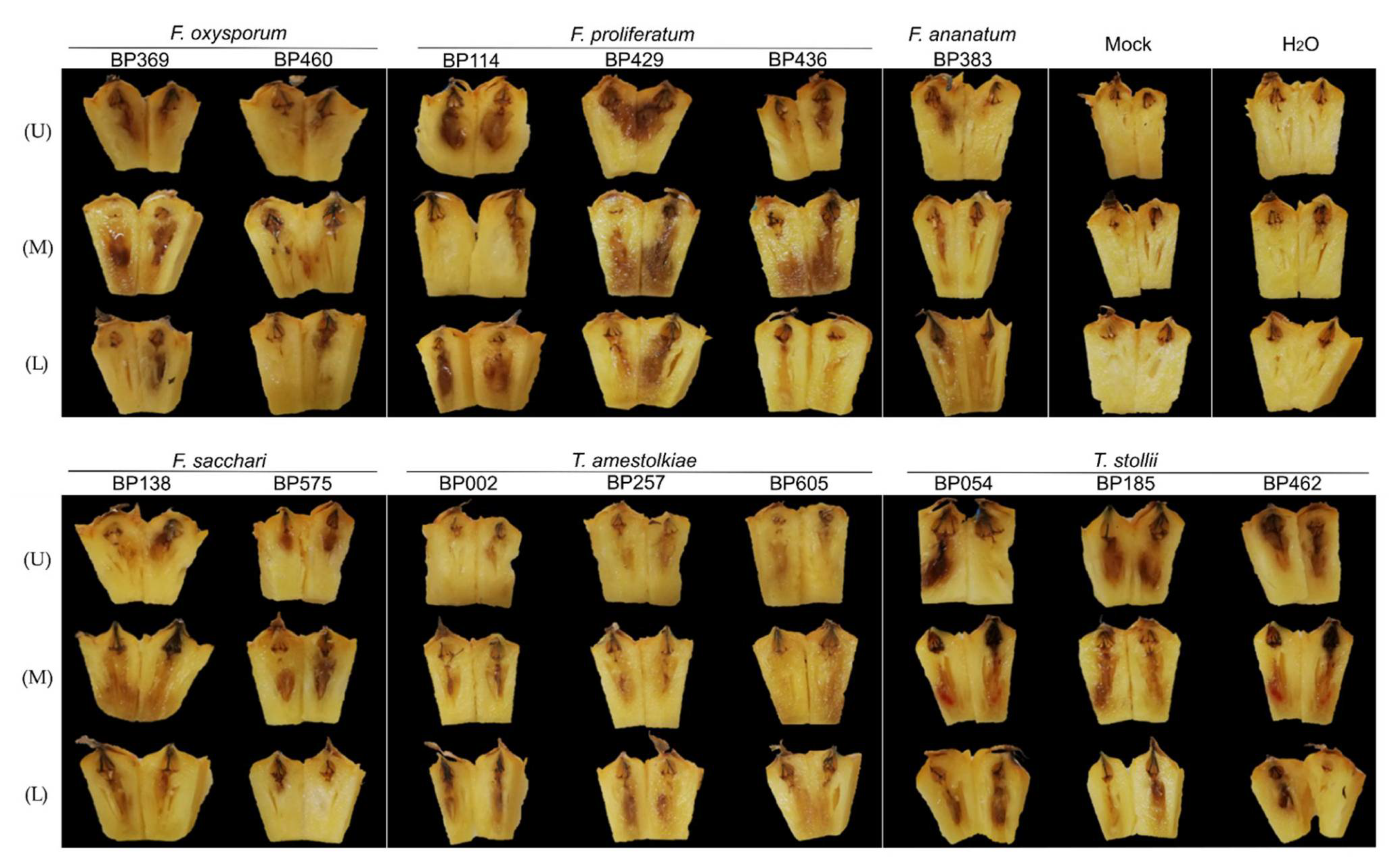
3.4. Koch’s Postulates for the Determination of New FCR Pathogens
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- FAO. Major Tropical Fruits—Preliminary Market Results 2019; Food and Agriculture Organization of the United Nations: Rome, Italy, 2020; p. 24. [Google Scholar]
- Bartholomew, D.P.; Hawkins, R.A.; Lopez, J.A. Hawaii Pineapple: The Rise and Fall of an Industry. HortScience 2012, 47, 1390–1398. [Google Scholar] [CrossRef] [Green Version]
- Nassr, M.S.; Abu-Naser, S.S. Knowledge Based System for Diagnosing Pineapple Diseases. Int. J. Acad. Pedagog. Res. 2018, 2, 12–17. [Google Scholar]
- Gu, H.; Zhan, R.L.; Zhan, L.B.; Gong, D.Q.; Jia, Z.W. First Report of Fusarium Ananatum Causing Pineapple Fruitlet Core Rot in China. Plant Dis. 2015, 99, 1653. [Google Scholar] [CrossRef]
- Rohrbach, K.G.; Pfeiffer, J.B. Susceptibility of Pineapple Cultivars to Fruit Diseases Incited by Penicillium Funiculosum and Fusarium Moniliforme. Phytopathology 1976, 66, 1386–1390. [Google Scholar] [CrossRef]
- Rohrbach, K.G.; Johnson, M.W. Pests, Diseases and Weeds. In The Pineapple: Botany, Production and Uses; Department of Tropical Plant and Soil Science, CTAHR, University of Hawaii at Manoa: Honolulu, HI, USA, 2003; pp. 203–251. [Google Scholar] [CrossRef]
- Seifert, K.A.; Aoki, T.; Baayen, R.P.; Brayford, D.; Burgess, L.W.; Chulze, S.; Gams, W.; Geiser, D.; de Gruyter, J.; Leslie, J.F.; et al. The Name Fusarium Moniliforme Should No Longer Be Used. Mycol. Res. 2003, 107, 643–644. [Google Scholar] [CrossRef]
- Lim, T.-K. Role of Penicillium Funiculosum Strains in the Development of Pineapple Fruit Diseases. Phytopathology 1980, 70, 663. [Google Scholar] [CrossRef]
- Petty, G.J.; Tustin, H.A.; Dicks, H.M. Control of black spot disease / fruitlet core rot in queen pineapple with integrated mealybug, pineapple fruit mite and fungus control programmes. Acta Hortic. 2006, 143–149. [Google Scholar] [CrossRef]
- Jacobs, A.; Van Wyk, P.S.; Marasas, W.F.O.; Wingfield, B.D.; Wingfield, M.J.; Coutinho, T.A. Fusarium Ananatum Sp. Nov. in the Gibberella Fujikuroi Species Complex from Pineapples with Fruit Rot in South Africa. Fungal Biol. 2010, 114, 515–527. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Barral, B.; Chillet, M.; Minier, J.; Léchaudel, M.; Schorr-Galindo, S. Evaluating the Response to Fusarium Ananatum Inoculation and Antifungal Activity of Phenolic Acids in Pineapple. Fungal Biol. 2017, 121, 1045–1053. [Google Scholar] [CrossRef] [PubMed]
- Mourichon, X. Pineapple Fruitlet Core Rot (Black Spot) and Leathery Pocket: Review and Prospects. Acta Hortic. 1997, 501–508. [Google Scholar] [CrossRef]
- Maltz, M.R.; Treseder, K.K.; McGuire, K.L. Links between Plant and Fungal Diversity in Habitat Fragments of Coastal Shrubland. PLoS ONE 2017, 12, e0184991. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hugoni, M.; Luis, P.; Guyonnet, J.; el Zahar Haichar, F. Plant Host Habitat and Root Exudates Shape Fungal Diversity. Mycorrhiza 2018, 28, 451–463. [Google Scholar] [CrossRef] [PubMed]
- Hawksworth, D.L.; Lücking, R. Fungal Diversity Revisited: 2.2 to 3.8 Million Species. In The Fungal Kingdom; Heitman, J., Howlett, B.J., Crous, P.W., Stukenbrock, E.H., James, T.Y., Gow, N.A.R., Eds.; ASM Press: Washington, DC, USA, 2017; pp. 79–95. ISBN 978-1-68367-082-7. [Google Scholar]
- Dos Santos Souza, C.R.; de Oliveira Barbosa, A.C.; Fortes Ferreira, C.; Vidigal Duarte Souza, F.; de Souza Rocha, L.; de Souza, E.H.; de Oliveira, S.A.S. Diversity of Microorganisms Associated to Ananas Spp. from Natural Environment, Cultivated and Ex Situ Conservation Areas. Sci. Hortic. 2019, 243, 544–551. [Google Scholar] [CrossRef]
- Rohrbach, K.G.; Taniguchi, G. Effects of Temperature, Moisture, and Stage of Inflorescence Development on Infection of Pineapple by Penicillium Funiculosum and Fusarium Moniliforme Var. Subglutinans. Phytopathology 1984, 74, 995–1000. [Google Scholar] [CrossRef]
- Fournier, P.; Benneveau, A.; Hardy, C.; Chillet, M.; Léchaudel, M. A Predictive Model Based on a Pluviothermic Index for Leathery Pocket and Fruitlet Core Rot of Pineapple Cv. ‘Queen’. Eur. J. Plant Pathol. 2015, 142, 449–460. [Google Scholar] [CrossRef]
- Gamliel-Atinsky, E.; Sztejnberg, A.; Maymon, M.; Shtienberg, D.; Freeman, S. Inoculum Availability and Conidial Dispersal Patterns of Fusarium Mangiferae, the Causal Agent of Mango Malformation Disease. Phytopathology 2009, 99, 160–166. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rieux, A.; Soubeyrand, S.; Bonnot, F.; Klein, E.K.; Ngando, J.E.; Mehl, A.; Ravigne, V.; Carlier, J.; de Lapeyre de Bellaire, L. Long-Distance Wind-Dispersal of Spores in a Fungal Plant Pathogen: Estimation of Anisotropic Dispersal Kernels from an Extensive Field Experiment. PLoS ONE 2014, 9, e103225. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Aylor, D.E. SPREAD OF PLANT DISEASE ON A CONTINENTAL SCALE: ROLE OF AERIAL DISPERSAL OF PATHOGENS. Ecology 2003, 84, 1989–1997. [Google Scholar] [CrossRef]
- Rieux, A.; Lenormand, T.; Carlier, J.; de Lapeyre de Bellaire, L.; Ravigné, V. Using Neutral Cline Decay to Estimate Contemporary Dispersal: A Generic Tool and Its Application to a Major Crop Pathogen. Ecol. Lett. 2013, 16, 721–730. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fredrikson, L.; Skinkis, P.A.; Peachey, E. Cover Crop and Floor Management Affect Weed Coverage and Density in an Establishing Oregon Vineyard. HortTechnology 2011, 21, 208–216. [Google Scholar] [CrossRef] [Green Version]
- Charest, J.; Dewdney, M.; Paulitz, T.; Philion, V.; Carisse, O. Spatial Distribution of Venturia Inaequalis Airborne Ascospores in Orchards. Phytopathology 2002, 92, 769–779. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Osborne, L.E.; Stein, J.M. Epidemiology of Fusarium Head Blight on Small-Grain Cereals. Int. J. Food Microbiol. 2007, 119, 103–108. [Google Scholar] [CrossRef] [PubMed]
- Dita, M.; Barquero, M.; Heck, D.; Mizubuti, E.S.G.; Staver, C.P. Fusarium Wilt of Banana: Current Knowledge on Epidemiology and Research Needs Toward Sustainable Disease Management. Front. Plant Sci. 2018, 9, 1468. [Google Scholar] [CrossRef] [Green Version]
- Van der Does, H.C.; Constantin, M.E.; Houterman, P.M.; Takken, F.L.W.; Cornelissen, B.J.C.; Haring, M.A.; van den Burg, H.A.; Rep, M. Fusarium Oxysporum Colonizes the Stem of Resistant Tomato Plants, the Extent Varying with the R-Gene Present. Eur. J. Plant Pathol. 2019, 154, 55–65. [Google Scholar] [CrossRef] [Green Version]
- Fournier, P. La Culture de l’ananas Victoria à La Réunion Pour l’exportation. In Recueil de Bonnes Pratiques; CIRAD: Reunion Island, France, 2011. [Google Scholar]
- Smart Information System (SmartIS)-METEOR-CIRAD-UR AÏDA. Available online: https://smartis.re/METEOR (accessed on 25 November 2020).
- Météo France. Available online: https://donneespubliques.meteofrance.fr/?fond=produit&id_produit=129&id_rubrique=52 (accessed on 30 September 2020).
- O’Donnell, K.; Cigelnik, E.; Nirenberg, H.I. Molecular Systematics and Phylogeography of the Gibberella Fujikuroi Species Complex. Mycologia 1998, 90, 465–493. [Google Scholar] [CrossRef]
- O’Donnell, K.; Kistler, H.C.; Cigelnik, E.; Ploetz, R.C. Multiple Evolutionary Origins of the Fungus Causing Panama Disease of Banana: Concordant Evidence from Nuclear and Mitochondrial Gene Genealogies. Proc. Natl. Acad. Sci. USA 1998, 95, 2044–2049. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liu, X.; Xing, M.; Kong, C.; Fang, Z.; Yang, L.; Zhang, Y.; Wang, Y.; Ling, J.; Yang, Y.; Lv, H. Genetic Diversity, Virulence, Race Profiling, and Comparative Genomic Analysis of the Fusarium Oxysporum f. Sp. Conglutinans Strains Infecting Cabbages in China. Front. Microbiol. 2019, 10, 1373. [Google Scholar] [CrossRef] [PubMed]
- Glass, N.L.; Donaldson, G.C. Development of Primer Sets Designed for Use with the PCR to Amplify Conserved Genes from Filamentous Ascomycetes. Appl. Environ. Microbiol. 1995, 61, 1323–1330. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- White, T.J.; Bruns, T.; Lee, S.; Taylor, J. Amplification And Direct Sequencing Of Fungal Ribosomal Rna Genes For Phylogenetics. In PCR Protocols; Academic Press: Cambridge, MA, USA, 1990; pp. 315–322. ISBN 978-0-12-372180-8. [Google Scholar]
- Gardes, M.; Bruns, T.D. ITS Primers with Enhanced Specificity for Basidiomycetes—Application to the Identification of Mycorrhizae and Rusts. Mol. Ecol. 1993, 2, 113–118. [Google Scholar] [CrossRef]
- Sheffield, V.C.; Cox, D.R.; Lerman, L.S.; Myers, R.M. Attachment of a 40-Base-Pair G + C-Rich Sequence (GC-Clamp) to Genomic DNA Fragments by the Polymerase Chain Reaction Results in Improved Detection of Single-Base Changes. Proc. Natl. Acad. Sci. USA 1989, 86, 232–236. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hamdouche, Y.; Meile, J.C.; Nganou, D.N.; Durand, N.; Teyssier, C.; Montet, D. Discrimination of Post-Harvest Coffee Processing Methods by Microbial Ecology Analyses. Food Control 2016, 65, 112–120. [Google Scholar] [CrossRef]
- Kumar, S.; Stecher, G.; Li, M.; Knyaz, C.; Tamura, K. MEGA X: Molecular Evolutionary Genetics Analysis across Computing Platforms. Mol. Biol. Evol. 2018, 35, 1547–1549. [Google Scholar] [CrossRef] [PubMed]
- Edgar, R.C. MUSCLE: Multiple Sequence Alignment with High Accuracy and High Throughput. Nucleic Acids Res. 2004, 32, 1792–1797. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Darriba, D.; Taboada, G.L.; Doallo, R.; Posada, D. JModelTest 2: More Models, New Heuristics and Parallel Computing. Nat. Methods 2012, 9, 772. [Google Scholar] [CrossRef] [Green Version]
- Guindon, S.; Dufayard, J.-F.; Lefort, V.; Anisimova, M.; Hordijk, W.; Gascuel, O. New Algorithms and Methods to Estimate Maximum-Likelihood Phylogenies: Assessing the Performance of PhyML 3.0. Syst. Biol. 2010, 59, 307–321. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Paradis, E.; Schliep, K. Ape 5.0: An Environment for Modern Phylogenetics and Evolutionary Analyses in R. Bioinformatics 2019, 35, 526–528. [Google Scholar] [CrossRef] [PubMed]
- Lê, S.; Josse, J.; Husson, F. FactoMineR: An R Package for Multivariate Analysis. J. Stat. Softw. 2008, 25. [Google Scholar] [CrossRef] [Green Version]
- Kassambara, A.; Mundt, F. Factoextra. 2020. Available online: https://cran.r-project.org/package=factoextra (accessed on 13 January 2021).
- Oksanen, J.; Blanchet, F.G.; Friendly, M.; Kindt, R.; Legendre, P.; McGlinn, D.; Minchin, P.R.; O’Hara, R.B.; Simpson, G.L.; Solymos, P.; et al. Vegan: Community Ecology Package. 2020. Available online: http://outputs.worldagroforestry.org/cgi-bin/koha/opac-detail.pl?biblionumber=39504# (accessed on 13 January 2021).
- Veech, J.A. A Probabilistic Model for Analysing Species Co-Occurrence: Probabilistic Model. Glob. Ecol. Biogeogr. 2013, 22, 252–260. [Google Scholar] [CrossRef]
- Griffith, D.M.; Veech, J.A.; Marsh, C.J. Cooccur: Probabilistic Species Co-Occurrence Analysis in R. J. Stat. Softw. 2016, 69. [Google Scholar] [CrossRef]
- Barral, B.; Chillet, M.; Doizy, A.; Grassi, M.; Ragot, L.; Léchaudel, M.; Durand, N.; Rose, L.J.; Viljoen, A.; Schorr-Galindo, S. Diversity and Toxigenicity of Fungi That Cause Pineapple Fruitlet Core Rot. Toxins 2020, 12, 339. [Google Scholar] [CrossRef] [PubMed]
- Wigmann, É.F.; Behr, J.; Vogel, R.F.; Niessen, L. MALDI-TOF MS Fingerprinting for Identification and Differentiation of Species within the Fusarium Fujikuroi Species Complex. Appl. Microbiol. Biotechnol. 2019, 103, 5323–5337. [Google Scholar] [CrossRef] [PubMed]
- Yilmaz, N.; Houbraken, J.; Hoekstra, E.S.; Frisvad, J.C.; Visagie, C.M.; Samson, R.A. Delimitation and Characterisation of Talaromyces Purpurogenus and Related Species. Persoonia Mol. Phylogeny Evol. Fungi 2012, 29, 39–54. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Oxenham, B. Etiology of Fruitlet Core Rot of Pineapple in Queensland. Qld. J. Agric. Sci. 1962, 19, 27–31. [Google Scholar]
- Barral, B.; Chillet, M.; Léchaudel, M.; Lartaud, M.; Verdeil, J.-L.; Conéjéro, G.; Schorr-Galindo, S. An Imaging Approach to Identify Mechanisms of Resistance to Pineapple Fruitlet Core Rot. Front. Plant Sci. 2019, 10, 1065. [Google Scholar] [CrossRef] [PubMed]
- Leneveu-Jenvrin, C.; Quentin, B.; Assemat, S.; Hoarau, M.; Meile, J.-C.; Remize, F. Changes of Quality of Minimally-Processed Pineapple (Ananas Comosus, Var. ‘Queen Victoria’) during Cold Storage: Fungi in the Leading Role. Microorganisms 2020, 8, 185. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ou, Y.; Penton, C.R.; Geisen, S.; Shen, Z.; Sun, Y.; Lv, N.; Wang, B.; Ruan, Y.; Xiong, W.; Li, R.; et al. Deciphering Underlying Drivers of Disease Suppressiveness Against Pathogenic Fusarium Oxysporum. Front. Microbiol. 2019, 10, 2535. [Google Scholar] [CrossRef] [Green Version]
- Lima, C.S.; Pfenning, L.H.; Costa, S.S.; Campos, M.A.; Leslie, J.F. A New Fusarium Lineage within the Gibberella Fujikuroi Species Complex Is the Main Causal Agent of Mango Malformation Disease in Brazil. Plant Pathol. 2009, 58, 33–42. [Google Scholar] [CrossRef]
- Zhan, R.-L.; Yang, S.-J.; Ho, H.-H.; Liu, F.; Zhao, Y.-L.; Chang, J.-M.; He, Y.-B. Mango Malformation Disease in South China Caused by Fusarium Proliferatum: Mango Abnormal Growth. J. Phytopathol. 2010, 158, 721–725. [Google Scholar] [CrossRef]
- García-Guzmán, G.; Heil, M. Life Histories of Hosts and Pathogens Predict Patterns in Tropical Fungal Plant Diseases. New Phytol. 2014, 201, 1106–1120. [Google Scholar] [CrossRef] [PubMed]
- Chen, L.; Zhou, J.; Zeng, T.; Miao, Y.; Mei, L.; Yao, G.; Fang, K.; Dong, X.; Sha, T.; Yang, M.; et al. Quantifying the Sharing of Foliar Fungal Pathogens by the Invasive Plant Ageratina Adenophora and Its Neighbours. New Phytol. 2020, 227, 1493–1504. [Google Scholar] [CrossRef] [PubMed]
- Parnell, S.; Gottwald, T.R.; Gilligan, C.A.; Cunniffe, N.J.; van den Bosch, F. The Effect of Landscape Pattern on the Optimal Eradication Zone of an Invading Epidemic. Phytopathology 2010, 100, 638–644. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Stępień, Ł.; Koczyk, G.; Waśkiewicz, A. Diversity of Fusarium Species and Mycotoxins Contaminating Pineapple. J. Appl. Genet. 2013, 54, 367–380. [Google Scholar] [CrossRef] [Green Version]
- Ibrahim, N.F.; Mohd, M.H.; Mohamed Nor, N.M.I.; Zakaria, L. Characterization of Fusarium Spp. Associated with Pineapple Fruit Rot and Leaf Spot in Peninsular Malaysia. J. Phytopathol. 2017, 165, 718–726. [Google Scholar] [CrossRef]
- Leneveu-Jenvrin, C.; Quentin, B.; Assemat, S.; Remize, F. Maintaining Physicochemical, Microbiological, and Sensory Quality of Pineapple Juice (Ananas Comosus, Var. ‘Queen Victoria’) through Mild Heat Treatment. Processes 2020, 8, 1186. [Google Scholar] [CrossRef]
- Rodriguez Estrada, A.E.; Hegeman, A.; Corby Kistler, H.; May, G. In Vitro Interactions between Fusarium Verticillioides and Ustilago Maydis through Real-Time PCR and Metabolic Profiling. Fungal Genet. Biol. 2011, 48, 874–885. [Google Scholar] [CrossRef] [PubMed]
- Hynes, J.; Müller, C.T.; Jones, T.H.; Boddy, L. Changes in Volatile Production During the Course of Fungal Mycelial Interactions Between Hypholoma Fasciculare and Resinicium Bicolor. J. Chem. Ecol. 2006, 33, 43–57. [Google Scholar] [CrossRef] [PubMed]
- Combès, A.; Ndoye, I.; Bance, C.; Bruzaud, J.; Djediat, C.; Dupont, J.; Nay, B.; Prado, S. Chemical Communication between the Endophytic Fungus Paraconiothyrium Variabile and the Phytopathogen Fusarium Oxysporum. PLoS ONE 2012, 7, e47313. [Google Scholar] [CrossRef]
- Marmann, A.; Aly, A.; Lin, W.; Wang, B.; Proksch, P. Co-Cultivation—A Powerful Emerging Tool for Enhancing the Chemical Diversity of Microorganisms. Mar. Drugs 2014, 12, 1043–1065. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Barriuso, J.; Hogan, D.A.; Keshavarz, T.; Martínez, M.J. Role of Quorum Sensing and Chemical Communication in Fungal Biotechnology and Pathogenesis. FEMS Microbiol. Rev. 2018, 42, 627–638. [Google Scholar] [CrossRef] [PubMed]
- Lu, Z.; Tombolini, R.; Woo, S.; Zeilinger, S.; Lorito, M.; Jansson, J.K. In Vivo Study of Trichoderma-Pathogen-Plant Interactions, Using Constitutive and Inducible Green Fluorescent Protein Reporter Systems. Appl. Environ. Microbiol. 2004, 70, 3073–3081. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lagashetti, A.C.; Dufossé, L.; Singh, S.K.; Singh, P.N. Fungal Pigments and Their Prospects in Different Industries. Microorganisms 2019, 7, 604. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Osmanova, N.; Schultze, W.; Ayoub, N. Azaphilones: A Class of Fungal Metabolites with Diverse Biological Activities. Phytochem. Rev. 2010, 9, 315–342. [Google Scholar] [CrossRef]
- Wei, Z.; Yang, T.; Friman, V.-P.; Xu, Y.; Shen, Q.; Jousset, A. Trophic Network Architecture of Root-Associated Bacterial Communities Determines Pathogen Invasion and Plant Health. Nat. Commun. 2015, 6, 8413. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Schouten, A.; van den Berg, G.; Edel-Hermann, V.; Steinberg, C.; Gautheron, N.; Alabouvette, C.; de Vos, C.H.(Ric); Lemanceau, P.; Raaijmakers, J.M. Defense Responses of Fusarium Oxysporum to 2,4-Diacetylphloroglucinol, a Broad-Spectrum Antibiotic Produced by Pseudomonas Fluorescens. Mol. Plant-Microbe Interact. 2004, 17, 1201–1211. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Santos, C.; Ventura, J.A.; Costa, H.; Fernandes, P.M.B.; Lima, N. MALDI-TOF MS to Identify the Pineapple Pathogen Fusarium Guttiforme and Its Antagonist Trichoderma Asperellum on Decayed Pineapple. Trop. Plant Pathol. 2015, 40, 227–232. [Google Scholar] [CrossRef]
- Crandall, S.G.; Gilbert, G.S. Meteorological Factors Associated with Abundance of Airborne Fungal Spores over Natural Vegetation. Atmos. Environ. 2017, 162, 87–99. [Google Scholar] [CrossRef]
- Dorey, E.; Fournier, P.; Léchaudel, M.; Tixier, P. Modeling Sugar Content of Pineapple under Agro-Climatic Conditions on Reunion Island. Eur. J. Agron. 2016, 73, 64–72. [Google Scholar] [CrossRef]
- Zheng, Q.; Ploetz, R. Genetic Diversity in the Mango Malformation Pathogen and Development of a PCR Assay: PCR Assay for Mango Malformation Pathogen. Plant Pathol. 2002, 51, 208–216. [Google Scholar] [CrossRef]
- Lin, Z.; Xu, S.; Que, Y.; Wang, J.; Comstock, J.C.; Wei, J.; McCord, P.H.; Chen, B.; Chen, R.; Zhang, M. Species-Specific Detection and Identification of Fusarium Species Complex, the Causal Agent of Sugarcane Pokkah Boeng in China. PLoS ONE 2014, 9, e104195. [Google Scholar] [CrossRef] [PubMed]
- Filières Végétales—DAAF de La Réunion. Available online: https://daaf.reunion.agriculture.gouv.fr/Filieres-Vegetales (accessed on 11 December 2020).
- Smart Information System (SmartIS)-VICTORIA-CIRAD-UR AÏDA. Available online: https://smartis.re/VICTORIA (accessed on 23 October 2020).
- Dorey, E.; Fournier, P.; Lechaudel, M.; Tixier, P. Validity of the Pineapple Crop Model SIMPIÑA across the Climatic Gradient in Réunion Island. Eur. J. Agron. 2015, 62, 1–12. [Google Scholar] [CrossRef]












| Min Temp (°C) | Average Temp (°C) | Max Temp (°C) | Average Rainfall (mm) | Number of Day(s) with Rain | Wind Direction | Max Gust Speed (km h−1) | |
|---|---|---|---|---|---|---|---|
| April | 20.12 | 23.19 | 27.85 | 15.78 * | 10 | south | 116 * |
| May | 18.42 | 22.20 | 27.48 | 0.66 | 5 | east-south-east | 85 |
| June | 17.10 | 20.68 | 25.72 | 2.47 | 8 | south-east | 73 |
| July | 15.53 | 19.50 | 24.64 | 4.46 | 12 | south-east | 80 |
| August | 17.07 | 21.10 | 26.60 | 0.00 | 0 | south-east | 75 |
| September | 16.64 | 21.15 | 27.60 | 0.15 | 1 | south-east | 80 |
| October | 17.65 | 21.75 | 27.79 | 3.85 | 8 | east-south-east | 88 |
| November | 18.98 | 23.49 | 28.66 | 0.03 | 1 | south-east | 69 |
| Genus | Species | Occurrence | Top Match GenBank Accession Number * (Frequency in Data) | |
|---|---|---|---|---|
| Healthy Fruitlet (n = 96) | Infected Fruitlet (n = 96) | |||
| Fusarium | proliferatum | 28 | 22 | KF993985.1 (12) |
| Fusarium | ananatum | 19 | 18 | MT010996.1 (36) |
| Talaromyces | stollii | 8 | 16 | JX315634.1 (24) |
| Fusarium | verticillioides | 12 | 12 | MT594370.1 (10) |
| Talaromyces | amestolkiae | 13 | 9 | KJ413360.1 (13) |
| Fusarium | equiseti | 10 | 11 | MN589630.1 (4) |
| Phialemoniopsis | curvata | 6 | 12 | AB278180.1 (15) |
| Fusarium | oxysporum | 9 | 9 | CP053267.1 (4) |
| Fusarium | sp. | 10 | 5 | JF740861.1 (5) |
| Fusarium | solani | 5 | 8 | MT594367.1 (3)/MK968891.1 (3) |
| Talaromyces | purpureogenus | 5 | 5 | KJ528885.1 (3)/MF476006.1 (3) |
| Aspergillus | flavus | 2 | 6 | MN955851.1 (3) |
| Fusarium | incarnatum | 3 | 5 | MK752398.1 (2)/MN882829.1 (2) |
| Fusarium | fujikuroi | 6 | 1 | MF281281.2 (4) |
| Aspergillus | niger | 3 | 4 | KY357318.1 (2)/MN788116.1 (2) |
| Fusarium | chlamydosporum | 3 | 1 | KJ125830.1 (3) |
| Talaromyces | funiculosus | 2 | 2 | AB893941.1 (3) |
| Penicillium | sp. | 3 | 1 | EU330619.1 (2) |
| Pestalotiopsis | trachicarpicola | 2 | 2 | MN295594.1 (4) |
| Fusarium | sacchari | 0 | 3 | MN193868.1 (2) |
| Trichoderma | erinaceum | 0 | 2 | MK109820.1 (2) |
| Fusarium | napiforme | 1 | 1 | MH862670.1 (2) |
| Trichoderma | paraviridescens | 1 | 1 | MF782827.1 (1)/MK418756.1 (1) |
| Epicoccum | sorghinum | 1 | 1 | MF782827.1 (1)/MK418756.1 (1) |
| Trichoderma | sp. | 0 | 2 | KX449479.1 (1)/MK870964.1 (1) |
| Aspergillus | awamori | 2 | 0 | KY416558.1 (2) |
| Fusarium | falciforme | 2 | 0 | MT251175.1 (2) |
| Lasiodiplodia | mahajangana | 2 | 0 | MH057188.1 (2) |
| Trichoderma | asperellum | 0 | 1 | KX538815.1 (1) |
| Fusarium | circinatum | 0 | 1 | MK334369.1 (1) |
| Fusarium | cortaderiae | 0 | 1 | AH012626.2 (1) |
| Aspergillus | fumigatus | 0 | 1 | MH844690.1 (1) |
| Fusarium | graminearum | 0 | 1 | MK460853.1 (1) |
| Trichoderma | harzianum | 0 | 1 | JN116710.1 (1) |
| Diaporthe | kongii | 0 | 1 | KR024740.1 (1) |
| Curvularia | lunata | 0 | 1 | MN971669.1 (1) |
| Diaporthe | masirevicii | 0 | 1 | MF668289.1 (1) |
| Aspergillus | novoparasiticus | 0 | 1 | MH279415.1 (1) |
| Aspergillus | oryzae | 0 | 1 | MN648727.1 (1) |
| Rhizopus | stolonifer | 0 | 1 | MF461025.1 (1) |
| Trichoderma | trixiae | 0 | 1 | MN889512.1 (1) |
| Pestalotiopsis | vismiae | 0 | 1 | KP747694.1 (1) |
| Clonostachys | wenpingii | 0 | 1 | NR_119651.1 (1) |
| Cosmospora | butyri | 1 | 0 | KU204560.1 (1) |
| Lasiodiplodia | citricola | 1 | 0 | KU530119.1 (1) |
| Glomus | clarum | 1 | 0 | AY035654.1 (1) |
| Fusarium | dlamini | 1 | 0 | MN173109.1 (1) |
| Fusarium | ficicrescens | 1 | 0 | KP662895.1 (1) |
| Bionectria | ochroleuca | 1 | 0 | EU552110.1 (1) |
| Clonostachys | rosea | 1 | 0 | MH047188.1 (1) |
| Clonostachys | sp. | 1 | 0 | MH681594.1 (1) |
| Davidiella | sp. | 1 | 0 | KX621979.1 (1) |
| Diaporthe | sp. | 1 | 0 | MH220834.1 (1) |
| Lasiodiplodia | theobromae | 1 | 0 | KR260829.1 (1) |
| Aspergillus | welwitschiae | 1 | 0 | MH374611.1 (1) |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Vignassa, M.; Meile, J.-C.; Chiroleu, F.; Soria, C.; Leneveu-Jenvrin, C.; Schorr-Galindo, S.; Chillet, M. Pineapple Mycobiome Related to Fruitlet Core Rot Occurrence and the Influence of Fungal Species Dispersion Patterns. J. Fungi 2021, 7, 175. https://0-doi-org.brum.beds.ac.uk/10.3390/jof7030175
Vignassa M, Meile J-C, Chiroleu F, Soria C, Leneveu-Jenvrin C, Schorr-Galindo S, Chillet M. Pineapple Mycobiome Related to Fruitlet Core Rot Occurrence and the Influence of Fungal Species Dispersion Patterns. Journal of Fungi. 2021; 7(3):175. https://0-doi-org.brum.beds.ac.uk/10.3390/jof7030175
Chicago/Turabian StyleVignassa, Manon, Jean-Christophe Meile, Frédéric Chiroleu, Christian Soria, Charlène Leneveu-Jenvrin, Sabine Schorr-Galindo, and Marc Chillet. 2021. "Pineapple Mycobiome Related to Fruitlet Core Rot Occurrence and the Influence of Fungal Species Dispersion Patterns" Journal of Fungi 7, no. 3: 175. https://0-doi-org.brum.beds.ac.uk/10.3390/jof7030175