
Potential Probiotic Strains of Saccharomyces and Non-Saccharomyces: Functional and Biotechnological Characteristics


Abstract
:1. Introduction
2. Materials and Methods
2.1. Yeast Strains
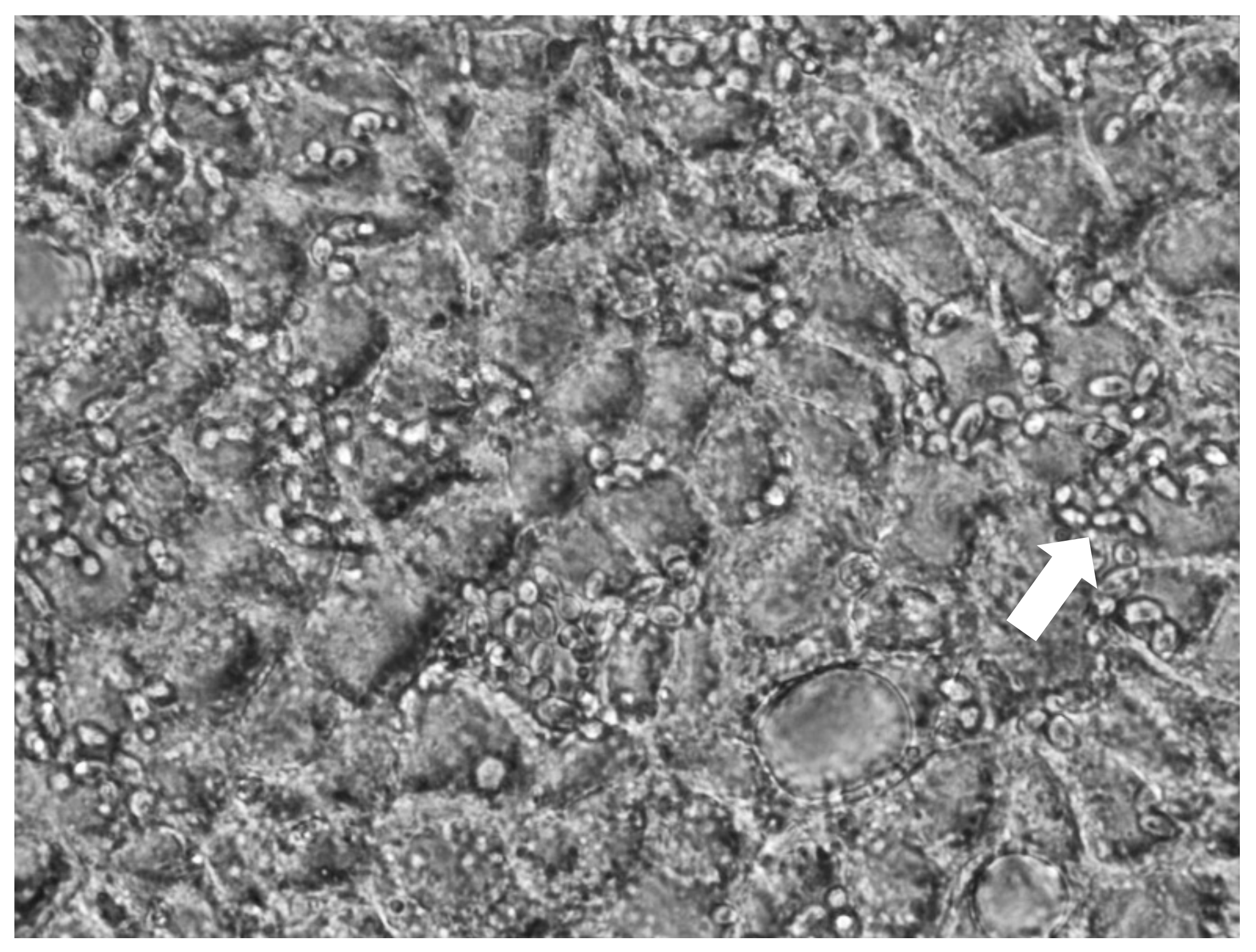
2.2. Yeast Adhesion to Caco-2/TC7 Cells
2.3. Prebiotic Metabolisms
2.3.1. Aerobic Conditions
2.3.2. Anaerobic Conditions
2.4. Assimilation of Cholesterol
2.5. Enzymatic Activity
2.6. Antioxidant Activity
2.7. Antifungal Resistance
2.8. Attenuation
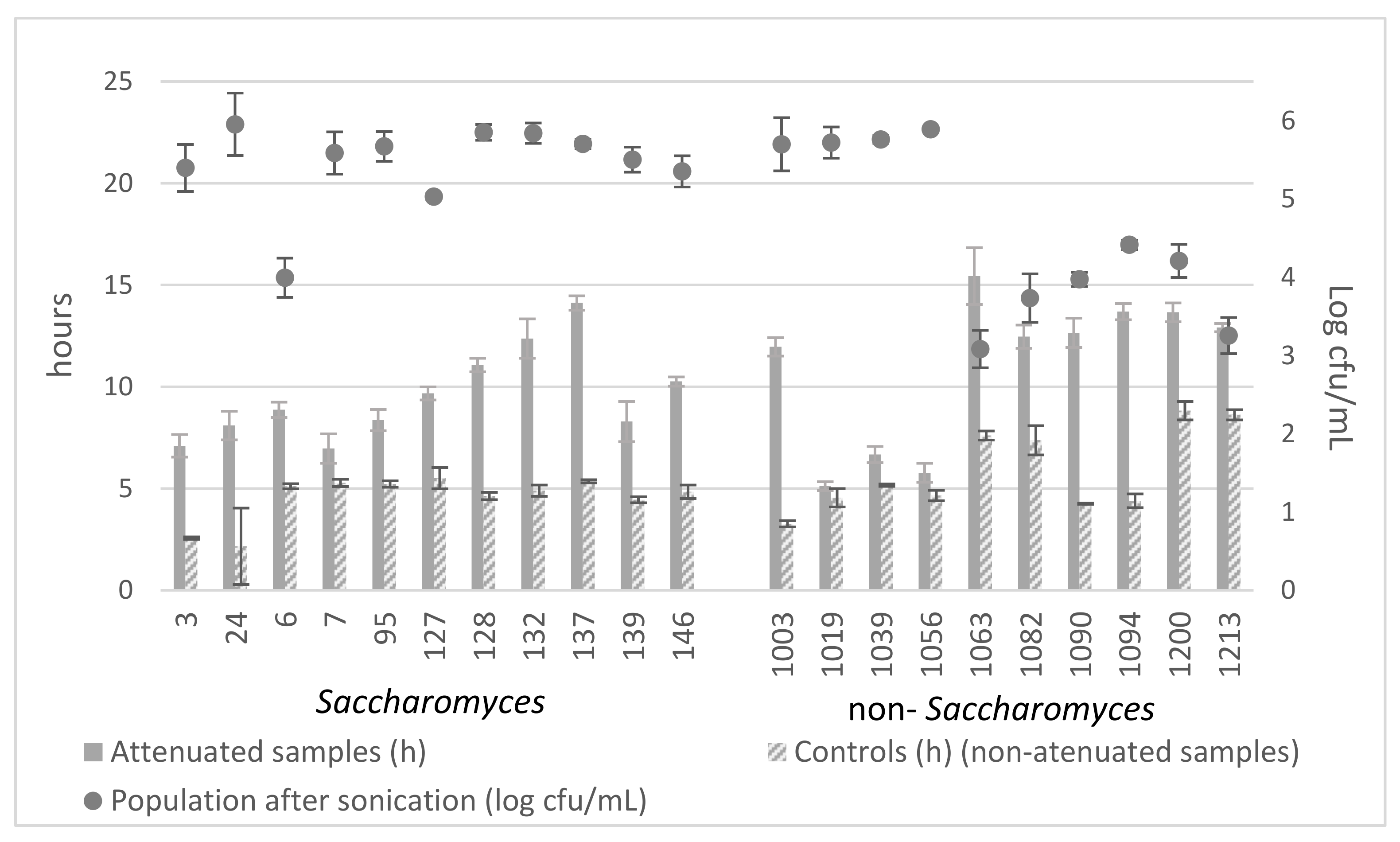
2.8.1. Setup of Method
2.8.2. Attenuation Effects
2.9. Statistical Analysis
3. Results and Discussion
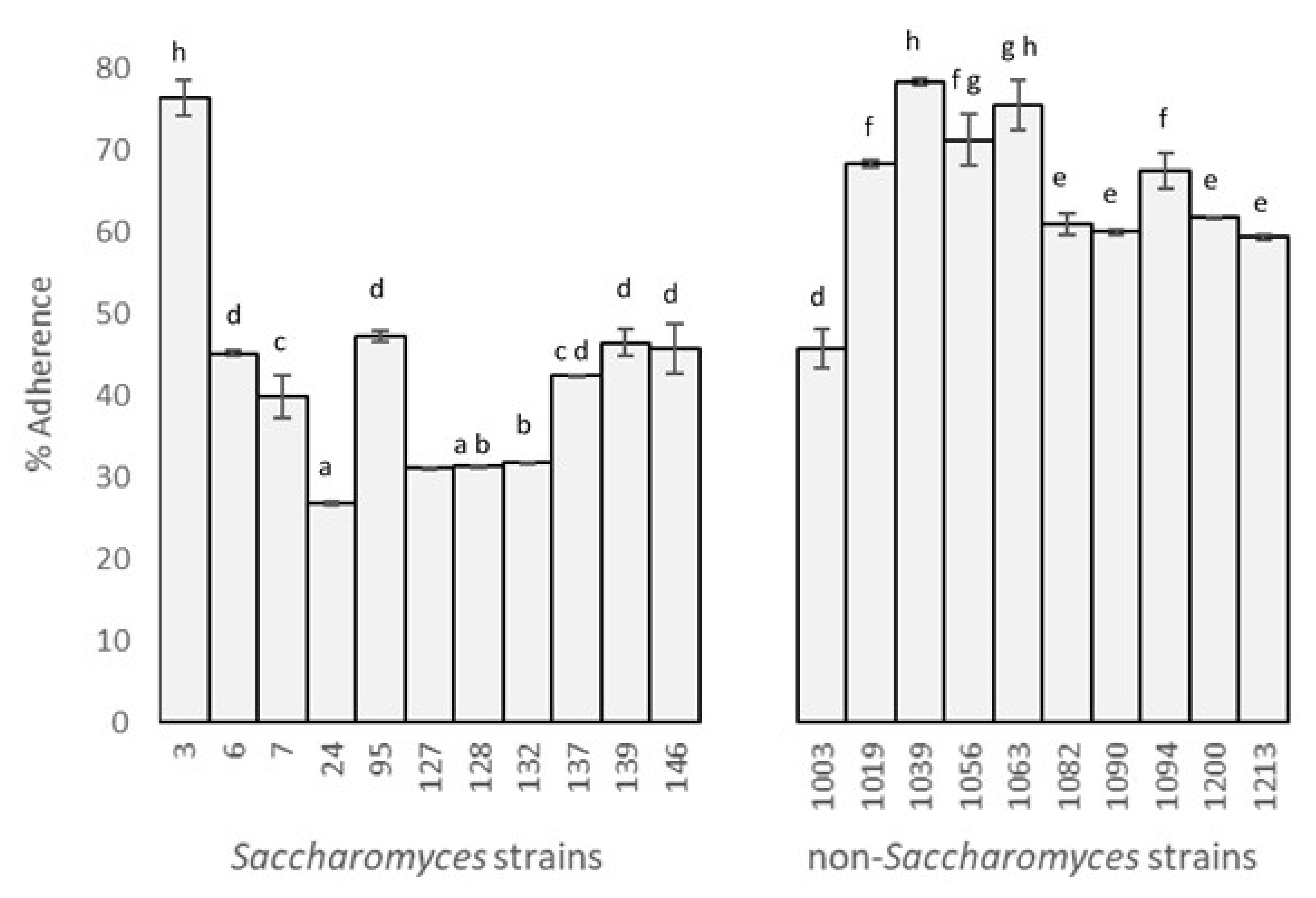
3.1. Yeast Adhesion to Caco-2/TC7 Cells
3.2. Prebiotics Metabolism
3.2.1. Aerobic Conditions
3.2.2. Anaerobic Conditions
3.3. Assimilation of Cholesterol
3.4. Enzymatic Activity
3.5. Antioxidant Activity
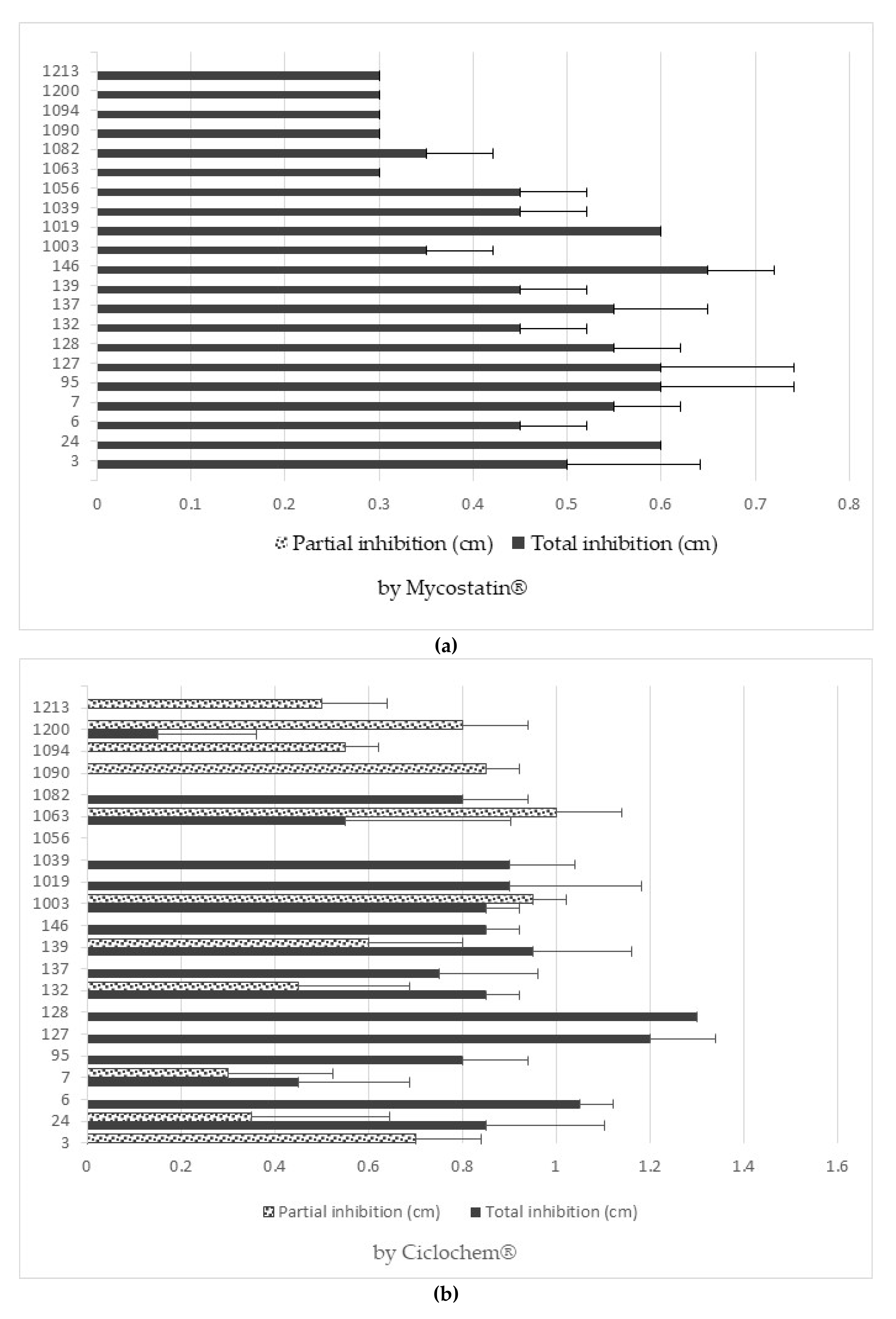
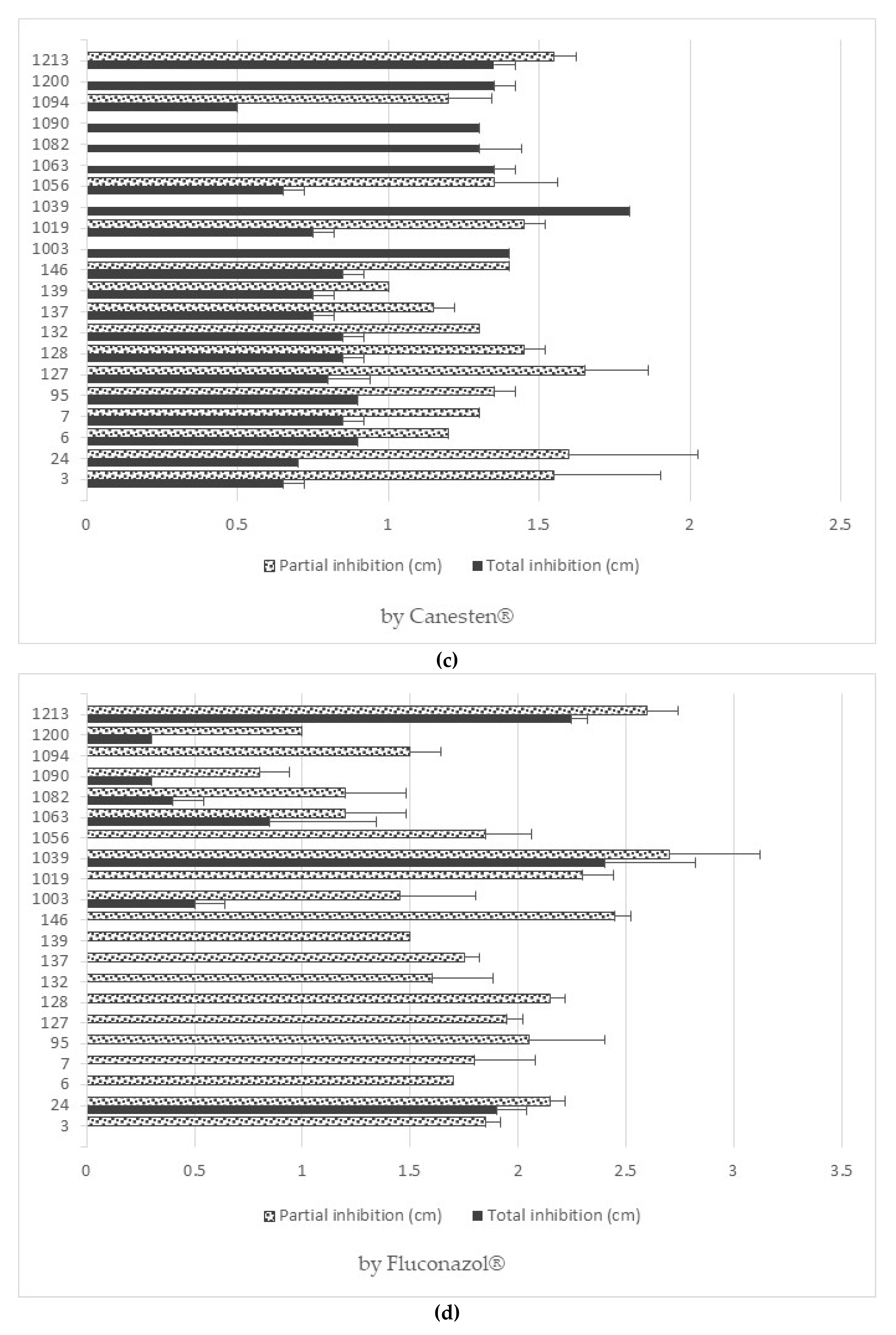
3.6. Antifungal Resistance
3.7. Attenuation
3.7.1. Setup of the Method
3.7.2. Attenuation Effects
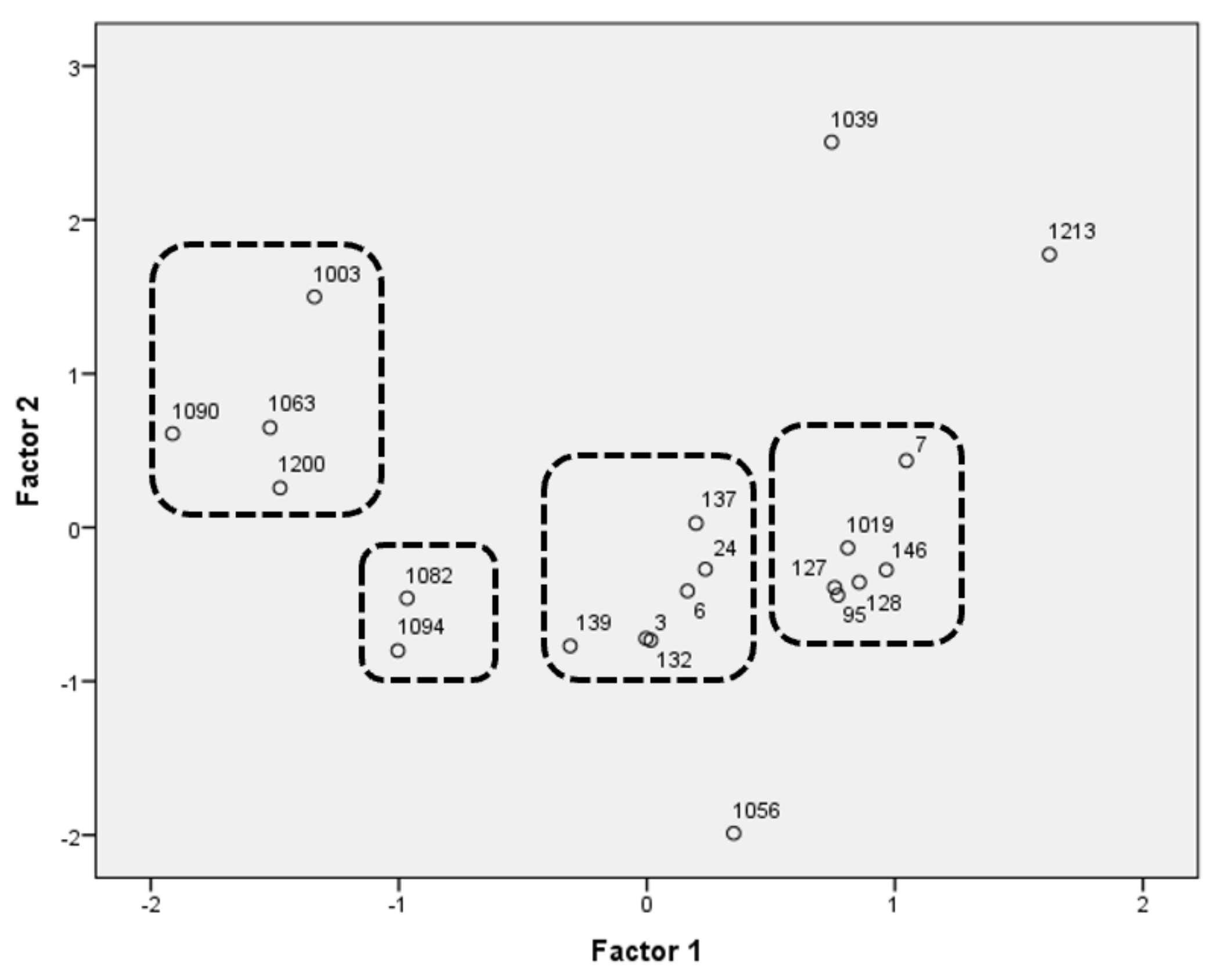
3.8. Selection of Strains
4. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Binetti, A.; Carrasco, M.; Reinheimer, J.; Suárez, V. Yeasts from autochthonous cheese starters: Technological and functional properties. J. Appl. Microbiol. 2013, 115, 434–444. [Google Scholar] [CrossRef] [PubMed]
- Chelliah, R.; Ramakrishnan, S.R.; Prabhu, P.R.; Antony, U. Evaluation of antimicrobial activity and probiotic properties of wild-strainPichia kudriavzeviiisolated from frozenidlibatter. Yeast 2016, 33, 385–401. [Google Scholar] [CrossRef]
- Silva, T.; Reto, M.; Sol, M.; Peito, A.; Peres, C.M.; Peres, C.; Malcata, F.X. Characterization of yeasts from Portuguese brined olives, with a focus on their potentially probiotic behavior. LWT-Food Sci. Technol. 2011, 44, 1349–1354. [Google Scholar] [CrossRef]
- Ogunremi, O.; Sanni, A.; Agrawal, R. Probiotic potentials of yeasts isolated from some cereal-based Nigerian traditional fermented food products. J. Appl. Microbiol. 2015, 119, 797–808. [Google Scholar] [CrossRef] [PubMed]
- Adams, M. Safety of industrial lactic acid bacteria. J. Biotechnol. 1999, 68, 171–178. [Google Scholar] [CrossRef]
- Moslehi-Jenabian, S.; Lindegaard, L.; Jespersen, L. Beneficial Effects of Probiotic and Food Borne Yeasts on Human Health. Nutrients 2010, 2, 449–473. [Google Scholar] [CrossRef] [PubMed]
- Meriggi, N.; Di Paola, M.; Vitali, F.; Rivero, D.; Cappa, F.; Turillazzi, F.; Gori, A.; Dapporto, L.; Beani, L.; Turillazzi, S.; et al. Saccharomyces cerevisiae Induces Immune Enhancing and Shapes Gut Microbiota in Social Wasps. Front. Microbiol. 2019, 10, 2320. [Google Scholar] [CrossRef] [Green Version]
- Li, Y.; Sadiq, F.A.; Liu, T.; Chen, J.; He, G. Purification and identification of novel peptides with inhibitory effects against angiotensin I-converting enzyme and optimization of process conditions in milk fermented with the yeast Kluyveromyces marxianus. J. Funct. Foods 2015, 16, 278–288. [Google Scholar] [CrossRef]
- FAO; WHO. Guidelines for the Evaluation of Probiotics in Food. In Food and Agriculture Organization of the United Nations and World Health Organization Working Group Report; FAO: Rome, Italy; WHO: Geneva, Switzerland, 2002. [Google Scholar]
- Salminen, S.; von Wright, A.; Morelli, L.; Marteau, P.; Brassart, D.; de Vos, W.M.; Fondén, R.; Saxelin, M.; Collins, K.; Mogensen, G.; et al. Demonstration of safety of probiotics—A review. Int. J. Food Microbiol. 1998, 44, 93–106. [Google Scholar] [CrossRef]
- Foligné, B.; Daniel, C.; Pot, B. Probiotics from research to market: The possibilities, risks and challenges. Curr. Opin. Microbiol. 2013, 16, 284–292. [Google Scholar] [CrossRef]
- Manson, J.E.; Tosteson, H.; Ridker, P.M.; Satterfield, S.; Hebert, P.; O’Connor, G.T.; Buring, J.E.; Hennekens, C.H. The Primary Prevention of Myocardial Infarction. N. Engl. J. Med. 1992, 326, 1406–1416. [Google Scholar] [CrossRef]
- Nita, M.; Grzybowski, A. The Role of the Reactive Oxygen Species and Oxidative Stress in the Pathomechanism of the Age-Related Ocular Diseases and Other Pathologies of the Anterior and Posterior Eye Segments in Adults. Oxidative Med. Cell. Longev. 2016, 2016, 1–23. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Suárez, J.E. Microbiota autóctona, probióticos y prebióticos. Nutr. Hosp. 2015, 31, 3–9. [Google Scholar] [PubMed]
- Bevilacqua, A.; Casanova, F.P.; Petruzzi, L.; Sinigaglia, M.; Corbo, M.R. Using physical approaches for the attenuation of lactic acid bacteria in an organic rice beverage. Food Microbiol. 2016, 53, 1–8. [Google Scholar] [CrossRef]
- Dai, C.; Xiong, F.; He, R.; Zhang, W.; Ma, H. Effects of low-intensity ultrasound on the growth, cell membrane permeability and ethanol tolerance of Saccharomyces cerevisiae. Ultrason. Sonochem. 2017, 36, 191–197. [Google Scholar] [CrossRef] [PubMed]
- Samani, B.H.; Khoshtaghaza, M.H.; Lorigooini, Z.; Minaei, S.; Zareiforoush, H. Analysis of the combinative effect of ultrasound and microwave power on Saccharomyces cerevisiae in orange juice processing. Innov. Food Sci. Emerg. Technol. 2015, 32, 110–115. [Google Scholar] [CrossRef]
- Arévalo-Villena, M.; Fernández-Pacheco, P.; Castillo, N.; Bevilacqua, A.; Briones, A. Probiotic yeast. Set up of a method for screening. LWT-Food Sci. Technol. 2018, 89, 657–665. [Google Scholar] [CrossRef]
- Fernandez-Pacheco, P.; Arévalo-Villena, M.; Bevilacqua, A.; Corbo, M.R.; Pérez, A.B. Probiotic characteristics in Saccharomyces cerevisiae strains: Properties for application in food industries. LWT-Food Sci. Technol. 2018, 97, 332–340. [Google Scholar] [CrossRef]
- Fernández-Pacheco Rodríguez, P.; Arévalo-Villena, M.; Zaparoli Rosa, I.; Briones Pérez, A. Selection of potential non-Sacharomyces probiotic yeasts from food origin by a step-by-step approach. Food Res. Int. 2018, 112, 143–151. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fernández-Pacheco, P.; Cueva, C.; Arévalo-Villena, M.; Moreno-Arribas, M.V.; Pérez, A.B. Saccharomyces cerevisiae and Hanseniaspora osmophila strains as yeast active cultures for potential probiotic applications. Food Funct. 2019, 10, 4924–4931. [Google Scholar] [CrossRef]
- Rodríguez, J.M. Probióticos: Del laboratorio al consumidor. Nutr. Hosp. 2015, 31, 33–47. [Google Scholar] [PubMed]
- Diosma, G.; Romanin, D.E.; Rey-Burusco, M.F.; Londero, A.; Garrote, G.L. Yeasts from kefir grains: Isolation, identification, and probiotic characterization. World J. Microbiol. Biotechnol. 2014, 30, 43–53. [Google Scholar] [CrossRef]
- Golowczyc, M.; Mobili, P.; Garrote, G.; Abraham, A.; De Antoni, G. Protective action of Lactobacillus kefir carrying S-layer protein against Salmonella enterica serovar Enteritidis. Int. J. Food Microbiol. 2007, 118, 264–273. [Google Scholar] [CrossRef] [PubMed]
- Kumar, M.; Nagpal, R.; Kumar, R.; Hemalatha, R.; Verma, V.; Kumar, A.; Chakraborty, C.; Singh, B.; Marotta, F.; Jain, S.; et al. Cholesterol-Lowering Probiotics as Potential Biotherapeutics for Metabolic Diseases. Exp. Diabetes Res. 2012, 2012, 1–14. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gilliland, S.E.; Nelson, C.R.; Maxwell, C. Assimilation of cholesterol by Lactobacillus acidophilus. Appl. Environ. Microbiol. 1985, 49, 377–381. [Google Scholar] [CrossRef] [Green Version]
- Brand-Williams, W.; Cuvelier, M.; Berset, C. Use of a free radical method to evaluate antioxidant activity. LWT-Food Sci. Technol. 1995, 28, 25–30. [Google Scholar] [CrossRef]
- Trotta, F.; Caldini, G.; Dominici, L.; Federici, E.; Tofalo, R.; Schirone, M.; Corsetti, A.; Suzzi, G.; Cenci, G. Food borne yeasts as DNA-bioprotective agents against model genotoxins. Int. J. Food Microbiol. 2012, 153, 275–280. [Google Scholar] [CrossRef]
- Whittenbury, R. Hydrogen Peroxide Formation and Catalase Activity in the Lactic Acid Bacteria. Microbiology 1964, 35, 13–26. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kumura, H.; Tanoue, Y.; Tsukahara, M.; Tanaka, T.; Shimazaki, K. Screening of Dairy Yeast Strains for Probiotic Applications. J. Dairy Sci. 2004, 87, 4050–4056. [Google Scholar] [CrossRef]
- Kurtzman, C.P.; Fell, J.W.; Boekhout, T. The Yeasts: A Taxonomic Study, 5th ed.; Elsevier: San Diego, CA, USA, 2011; Volume 2. [Google Scholar]
- Psomas, E.; Fletouris, D.; Litopoulou-Tzanetaki, E.; Tzanetakis, N. Assimilation of Cholesterol by Yeast Strains Isolated from Infant Feces and Feta Cheese. J. Dairy Sci. 2003, 86, 3416–3422. [Google Scholar] [CrossRef] [Green Version]
- Chen, L.-S.; Ma, Y.; Maubois, J.-L.; He, S.-H.; Li, H.-M.; Chen, L.-J. Screening for the potential probiotic yeast strains from raw milk to assimilate cholesterol. Dairy Sci. Technol. 2010, 90, 537–548. [Google Scholar] [CrossRef] [Green Version]
- Psomas, E.; Andrighetto, C.; Litopoulou-Tzanetaki, E.; Lombardi, A.; Tzanetakis, N. Some probiotic properties of yeast isolates from infant faeces and Feta cheese. Int. J. Food Microbiol. 2001, 69, 125–133. [Google Scholar] [CrossRef]
- Lee, N.K.; Yun, C.W.; Kim, S.W.; Chang, H.I.; Kang, C.W.; Paik, H.D. Screening of Lactobacilli derived from chicken feces and partial characterization of Lactobacillus acidophilus A12 as an animal probiotic. J. Microbiol. Biotechnol. 2008, 18, 338–342. [Google Scholar] [PubMed]
- Bilski, J.; Mazur-Bialy, A.; Wojcik, D.; Zahradnik-Bilska, J.; Brzozowski, B.; Magierowski, M.; Mach, T.; Magierowska, K.; Brzozowski, T. The Role of Intestinal Alkaline Phosphatase in Inflammatory Disorders of Gastrointestinal Tract. Mediat. Inflamm. 2017, 2017, 1–9. [Google Scholar] [CrossRef]
- De Lima, M.D.S.F.; De Souza, K.M.S.; Albuquerque, W.W.C.; Teixeira, J.A.C.; Cavalcanti, M.T.H.; Porto, A.L.F. Saccharomyces cerevisiae from Brazilian kefir-fermented milk: An in vitro evaluation of probiotic properties. Microb. Pathog. 2017, 110, 670–677. [Google Scholar] [CrossRef] [Green Version]
- França, M.; Panek, A.; Eleutherio, E. Oxidative stress and its effects during dehydration. Comp. Biochem. Physiol. Part A Mol. Integr. Physiol. 2007, 146, 621–631. [Google Scholar] [CrossRef] [PubMed]
- Herrero, E.; Ros, J.; Bellí, G.; Cabiscol, E. Redox control and oxidative stress in yeast cells. Biochim. Biophys. Acta (BBA)-Gen. Subj. 2008, 1780, 1217–1235. [Google Scholar] [CrossRef] [PubMed]
- Tomusiak-Plebanek, A.; Heczko, P.; Skowron, B.; Baranowska, A.; Okoń, K.; Thor, P.J.; Strus, M. Lactobacilli with superoxide dismutase-like or catalase activity are more effective in alleviating inflammation in an inflammatory bowel disease mouse model. Drug Des. Dev. Ther. 2018, 12, 3221. [Google Scholar] [CrossRef] [PubMed] [Green Version]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Species | Strains | Source |
|---|---|---|
| Saccharomyces boulardii | 24 | CP |
| Saccharomyces cerevisiae | 3, 6, 7, 95, 127, 128, 132, 137, 139, 146 | W |
| Pichia kudriavzevii | 1003, 1200 | W |
| Pichia membranifaciens | 1019 | W |
| Lachancea thermotolerans | 1039 | W |
| Hanseniaspora osmophila | 1056, 1094 | D |
| Candida vini | 1063 | W |
| Pichia anomala | 1082, 1090 | D |
| Zygosaccharomyces bailii | 1213 | FV |
| STRAINS | Prebiotic/Assimilation Time | |||||||||
|---|---|---|---|---|---|---|---|---|---|---|
| Aerobic Conditions | ||||||||||
| Cellulose | Inulin | Melibiose | Raffinose | Xylan | Trehalose | Pectin | Cellobiose | Beta-Glucans | ||
| Saccharomyces | 3 | - | - | - | 0.56 (0–24] | - | - | 0.50 (48–2] | - | - |
| 24 | - | - | - | 0.83 (0–24] | - | - | 0.53 (48–72] | 0.51 (48–72] | - | |
| 6 | - | - | - | 0.93 (0–24] | - | 0.53 > 72 | 0.55 (24–48] | 0.50 (24–48] | 0.54 (24–48] | |
| 7 | - | - | - | 0.81 (0–24] | - | 0.51 > 72 | 0.58 > 72 | - | - | |
| 95 | - | - | - | 0.84 (0–24] | - | - | 0.53 > 72 | - | - | |
| 127 | - | - | 0.61 (24–48] | 0.92 (0–24] | 0.52 (48–72] | 0.72 > 72 | 0.52 (48–72] | 0.51 (48–72] | - | |
| 128 | - | - | 0.52 (24–48] | 0.99 (0–24] | - | 0.58 > 72 | 0.53 (24–48] | 0.61 (48–72] | - | |
| 132 | - | - | - | 0.59 (0–24] | - | - | 0.55 (48-–72] | - | - | |
| 137 | - | 0.52 > 72 | 0.56 > 72 | 0.73 (0–24] | - | 0.55 > 72 | 0.52 > 72 | - | - | |
| 139 | - | 0.61 (48–72] | 0.52 > 72 | 0.97 (0–24] | - | 0.67 (0–24] | 0,.52 (48–72] | 0.70 > 72 | - | |
| 146 | - | - | 0.50 > 72 | 1.16 (0–24] | - | - | 0.50 > 72 | 0.60 > 72 | - | |
| Non-Saccharomyces | 1003 | - | - | 0.69 (0–24] | 0.72 (0–24] | 0.58 (0–24] | 0.74 (0–24] | 0.89 (0–24] | 0.59 (0–24] | 1.14 (0–24] |
| 1019 | - | - | - | 0.69 (0–24] | - | - | - | - | - | |
| 1039 | - | - | 0.75 (24–48] | 0.86 (0–24] | 0.72 (24–48] | 0.70 (24–48] | 0.74 > 72 | 0.54 (48–72] | 0.58 (48–72] | |
| 1056 | - | - | - | 0.56 (0–24] | - | - | - | - | 0.76 (0–24] | |
| 1063 | - | - | - | 0.53 (0–24] | 0.56 (0–24] | 0.54 (0–24] | 0.55 (0–24] | 0.86 (0–24] | 0.95 (0–24] | |
| 1082 | - | 0.56 (0–24] | - | 0.53 (0–24] | 0.55 (0–24] | - | 0.80 (0–24] | 0.89 (0–24] | 0.89 (0–24] | |
| 1090 | - | 0.88 (0–24] | 0.66 (0–24] | - | 0.55 (0–24] | 0.63 (0–24] | 0.96 (0–24] | 1.08 (0–24] | 1.12 (0–24] | |
| 1094 | 0.6 (0–24] | 0.73 (0–24] | 0.94 (0–24] | 0.86 (0–24] | 0.77 (0–24] | 1.56 (0–24] | 0.94 (0–24] | 1.60 (0–24] | 1.27 (0–24] | |
| 1200 | - | 0.61 > 72 | - | - | - | 0.50 (0–24] | 0.82 (0–24] | 0,.2 (0–24] | 1.06 (0–24] | |
| 1213 | - | - | - | - | - | - | - | - | 0.52 (24–48] | |
| Anaerobic Conditions | ||||||||||
| Cellulose | Melibiose | Raffinose | Xylan | Trehalose | Cellobiose | Beta-Glucans | ||||
| Saccharomyces | 3 | - | - | - | - | - | - | - | ||
| 24 | - | - | - | - | 0.56 > 72 | 0.57 (24–48] | - | |||
| 6 | - | - | - | - | - | - | - | |||
| 7 | - | - | - | - | 0.50 > 72 | - | - | |||
| 95 | - | - | - | - | 0.53 (48–72] | - | - | |||
| 127 | - | - | - | - | 0.58 > 72 | 0.57 (48–72] | - | |||
| 128 | - | - | - | - | 0.72 > 72 | 0.58 > 72 | - | |||
| 132 | - | - | 0.53 > 72 | - | 0.55 (48–72] | - | - | |||
| 137 | - | - | - | - | 0.56 (48–72] | - | - | |||
| 139 | - | - | 0.56 > 72 | - | 0.56 (48–72] | 0.55 (0–24] | - | |||
| 146 | - | - | - | - | 0.70 > 72 | 0.58 (0–24] | - | |||
| Non-Saccharomyces | 1003 | - | - | - | - | - | 0.50 (24–48] | - | ||
| 1019 | - | - | - | 0.72 (24–48] | - | - | - | |||
| 1039 | 0.50 (0–24] | - | - | - | - | 0.51 (24–48] | 0.68 (24–48] | |||
| 1056 | 0.57 (24–48] | - | 0.91 (0–24] | 0.76 (48–72] | 0.74 (24–48] | - | - | |||
| 1063 | - | - | - | 0.64 (24–48] | - | 0.54 (24–48] | 0.64 (24–48] | |||
| 1082 | - | - | 0.51 (24–48] | 0.56 (0–24] | - | - | - | |||
| 1090 | - | - | - | 0.58 (48–72] | - | - | - | |||
| 1094 | - | - | - | - | - | 0.58 (0–24] | - | |||
| 1200 | - | - | - | - | 0.54 > 72 | - | - | |||
| 1213 | - | 0,65 (48–72] | - | 0.50 (24–48] | - | - | - | |||
| Strain | Enzyme Studied * | |||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| 02 | 03 | 04 | 05 | 06 | 07 | 08 | 09 | 10 | 11 | 12 | ||
| Saccharomyces cerevisiae | 3 | - | + | + | - | + | + | + | - | - | + | + |
| 6 | - | - | - | - | + | - | - | - | - | - | + | |
| 7 | - | + | + | - | + | - | - | - | - | + | + | |
| 24 | - | + | + | - | + | - | - | - | - | + | + | |
| 95 | - | + | + | - | + | - | - | - | - | - | + | |
| 127 | - | + | + | - | + | - | - | - | - | + | + | |
| 128 | - | + | + | - | + | - | - | - | - | + | + | |
| 132 | - | - | - | - | + | - | - | - | - | + | + | |
| 137 | - | + | - | - | + | - | - | - | - | + | + | |
| 139 | - | + | + | - | + | - | - | - | - | + | + | |
| 146 | - | - | - | - | + | - | - | - | - | + | + | |
| Non-Saccharomyces | 1003 | - | + | + | - | + | - | - | - | - | + | + |
| 1019 | + | + | + | - | + | - | - | - | - | + | + | |
| 1039 | - | - | - | - | + | - | - | - | - | - | + | |
| 1056 | - | - | - | - | + | - | - | - | - | + | + | |
| 1063 | - | + | + | + | + | - | - | - | - | + | + | |
| 1082 | - | + | - | - | + | - | - | - | - | + | + | |
| 1090 | - | + | + | - | + | - | - | - | - | + | + | |
| 1094 | - | + | + | - | + | - | - | - | - | + | + | |
| 1200 | - | + | + | - | + | - | - | - | - | - | + | |
| Amplitude (%) | Total Time (min) | Pulses (s) | Initial Population 107 cfu/mL | Initial Population 106 cfu/mL | |
|---|---|---|---|---|---|
| On | Off | ||||
| 80 | 3 | - | - | 58.5% | 60.9% |
| 80 | 2 | 2 | 10 | 47.1% | 45.4% |
| 80 | 1 | 2 | 10 | 3.1% | 2.1% |
| 60 | 3 | - | - | 37.7% | 42.6% |
| 60 | 3 | 2 | 10 | 26.9% | 30.2% |
| 60 | 2 | 2 | 10 | 6.9% | 6.1% |
| 60 | 1 | 2 | 10 | 0% | 2.2% |
| Principal Component | Variance Explained (%) | Total Variance (%) | Most Highly Correlated | Loading |
|---|---|---|---|---|
| 1 | 27.38 | 27.38 | Fluconazol PI Canesten PI Mycostatin TI Ciclochem PI Pectin | 0.87 0.80 0.71 −0.70 −0.55 |
| 2 | 20.38 | 47.77 | Fluconazol TI Raffinose Canesten TI Trehalose | 0.81 −0.80 0.79 −0.69 |
| 3 | 11.34 | 59.11 | β-glucan Xylan Cellulose DPPH Attenuation vital. | 0.79 0.78 0.77 −0.73 −0.56 |
| 4 | 10.04 | 69.15 | Ciclochem TI Cellobiose Melibiose | 0.85 0.72 −0.55 |
| 5 | 8.46 | 77.61 | Cholesterol Attenuation | 0.88 −0.62 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Fernández-Pacheco, P.; Pintado, C.; Briones Pérez, A.; Arévalo-Villena, M. Potential Probiotic Strains of Saccharomyces and Non-Saccharomyces: Functional and Biotechnological Characteristics. J. Fungi 2021, 7, 177. https://0-doi-org.brum.beds.ac.uk/10.3390/jof7030177
Fernández-Pacheco P, Pintado C, Briones Pérez A, Arévalo-Villena M. Potential Probiotic Strains of Saccharomyces and Non-Saccharomyces: Functional and Biotechnological Characteristics. Journal of Fungi. 2021; 7(3):177. https://0-doi-org.brum.beds.ac.uk/10.3390/jof7030177
Chicago/Turabian StyleFernández-Pacheco, Pilar, Cristina Pintado, Ana Briones Pérez, and María Arévalo-Villena. 2021. "Potential Probiotic Strains of Saccharomyces and Non-Saccharomyces: Functional and Biotechnological Characteristics" Journal of Fungi 7, no. 3: 177. https://0-doi-org.brum.beds.ac.uk/10.3390/jof7030177