
Dexamethasone Creates a Suppressive Microenvironment and Promotes Aspergillus fumigatus Invasion in a Human 3D Epithelial/Immune Respiratory Model

 , , and
, , and 
Abstract
:1. Introduction
2. Materials and Methods
2.1. Dexamethasone Preparation and Treatment
2.2. Isolation of Monocytes and Macrophage Differentiation
2.3. Respiratory MODEL
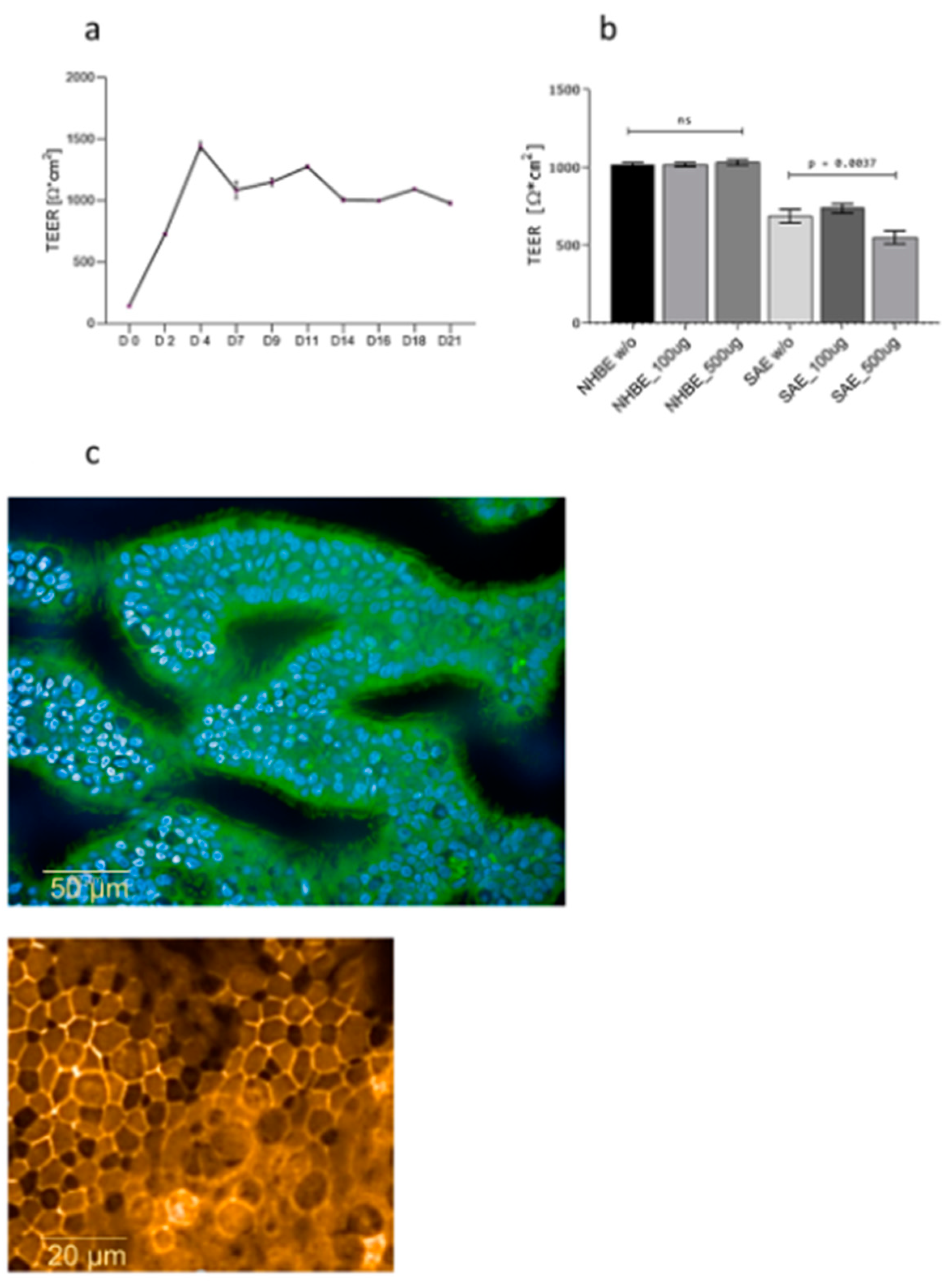
2.4. TEER
2.5. Macrophage–NHBE Coculture
2.6. Chemotaxis and Transmigration Assays
2.7. Aspergillus (A.) fumigatus Culture
2.8. Multi-Color Flow Cytometry
2.9. A. fumigatus Infection of the 3D Respiratory Model
2.10. NHBE Membrane Immunostaining, Mounting and HCS (High Content Screening)
2.11. ELISA for Cytokine Assessment
2.12. Statistical Analysis
3. Results
3.1. Dexamethasone Does Not Disrupt Epithelial Integrity
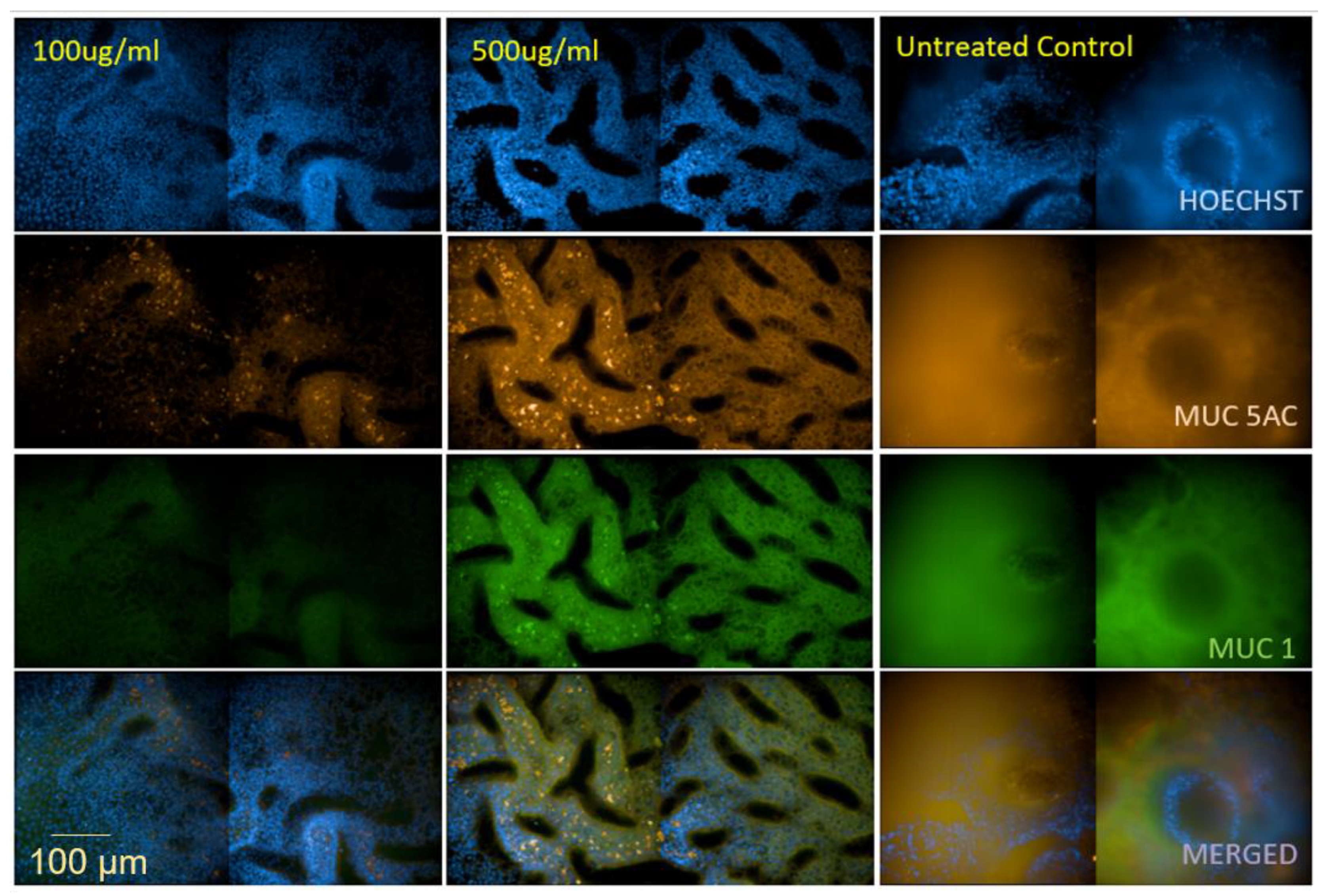
3.2. Dexamethasone Exerts an Impact on Mucus Production
3.3. Dexamethasone Reduces Ciliary Beat Frequency within the 3D Lung Model Independent of A. fumigatus Infection
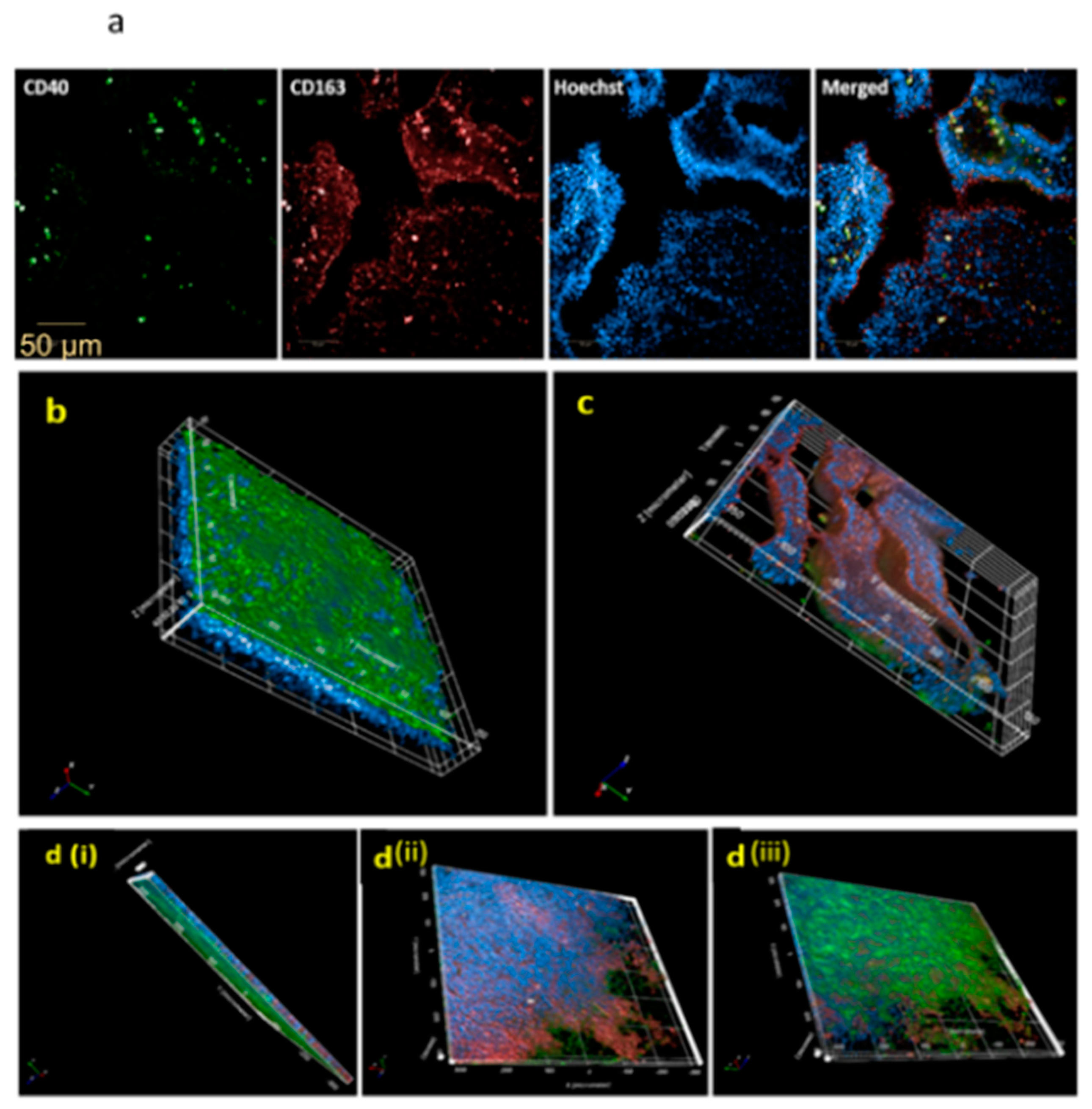
3.4. Dexamethasone Induces an Immunosuppressed Microenvironment Characterized by CD163 Expression and Obstructed Macrophage Trans-Epithelial Migration
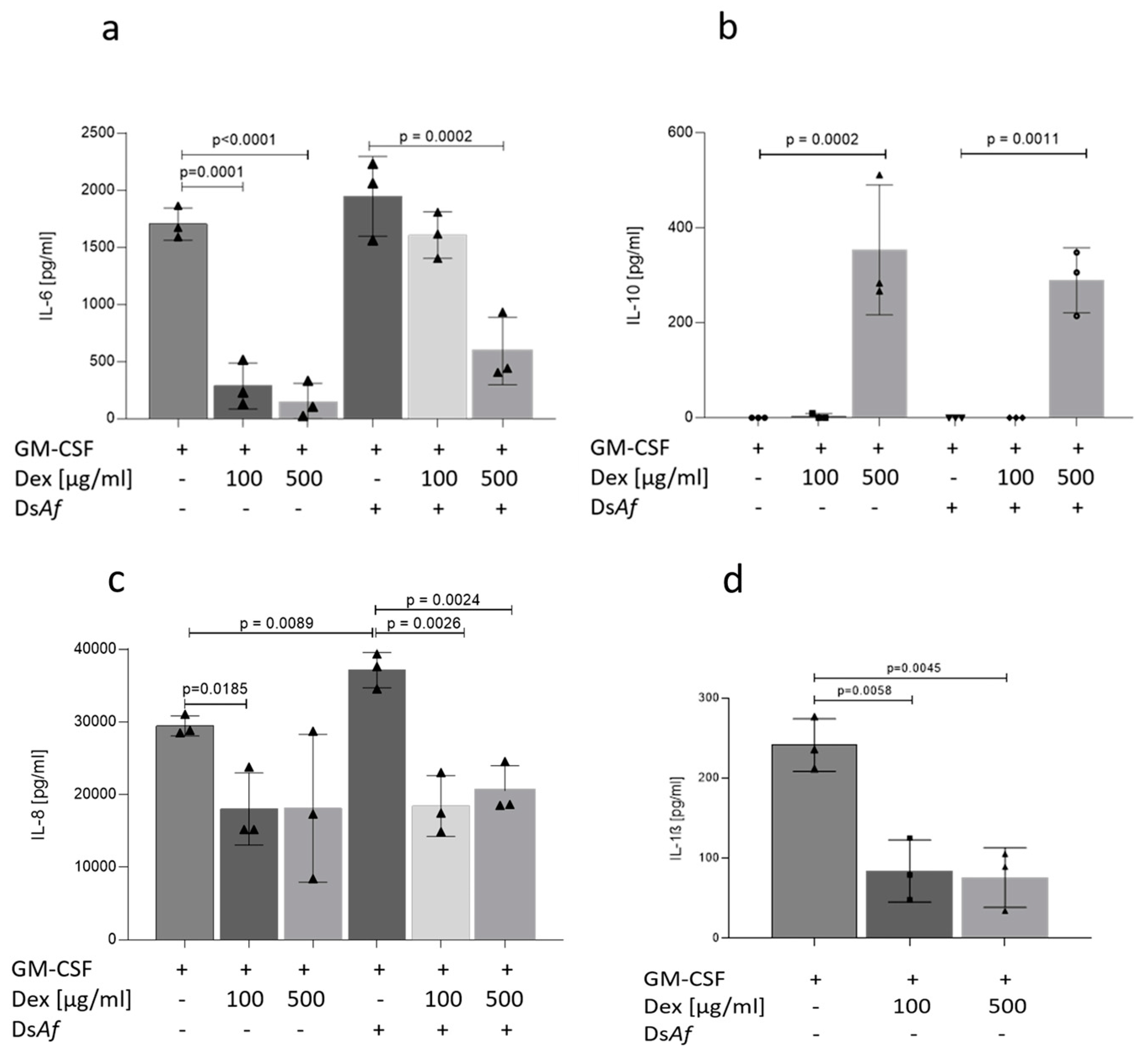
3.5. Dexamethasone Treatment Skews Epithelial/Immune Lung Tissues to an Anti-Inflammatory Pattern Independent on A. fumigatus Infection
3.6. Dexamethasone Enhances Intra-Epithelial Fungal Invasion within the 3D Lung Model
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Diehl, R.; Ferrara, F.; Müller, C.; Dreyer, A.Y.; McLeod, D.D.; Fricke, S.; Boltze, J. Immunosuppression for in vivo research: State-of-the-art protocols and experimental approaches. Cell. Mol. Immunol. 2017, 14, 146–179. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kawalkowska, J.Z.; Hemmerle, T.; Pretto, F.; Matasci, M.; Neri, D.; Williams, R.O. Targeted IL-4 therapy synergizes with dexamethasone to induce a state of tolerance by promoting Treg cells and macrophages in mice with arthritis. Eur. J. Immunol. 2016, 46, 1246–1257. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Suntharalingam, G.; Perry, M.R.; Ward, S.; Brett, S.J.; Castello-Cortes, A.; Brunner, M.D.; Panoskaltsis, N. Cytokine Storm in a Phase 1 Trial of the Anti-CD28 Monoclonal Antibody TGN1412. N. Engl. J. Med. 2006, 355, 1018–1028. [Google Scholar] [CrossRef] [PubMed]
- Dagenais, T.R.T.; Keller, N.P. Pathogenesis of Aspergillus fumigatus in Invasive Aspergillosis. Clin. Microbiol. Rev. 2009, 22, 447–465. [Google Scholar] [CrossRef] [Green Version]
- Zaidman, N.A.; Panoskaltsis-Mortari, A.; O’Grady, S.M. Differentiation of human bronchial epithelial cells: Role of hydrocortisone in development of ion transport pathways involved in mucociliary clearance. Am. J. Physiol. Physiol. 2016, 311, C225–C236. [Google Scholar] [CrossRef] [Green Version]
- Rayner, R.E.; Makena, P.; Prasad, G.L.; Cormet-Boyaka, E. Optimization of Normal Human Bronchial Epithelial (NHBE) Cell 3D Cultures for in vitro Lung Model Studies. Sci. Rep. 2019, 9, 1–10. [Google Scholar] [CrossRef]
- Hauber, H.-P.; Goldmann, T.; Vollmer, E.; Wollenberg, B.; Zabel, P. Effect of Dexamethasone and ACC on Bacteria-Induced Mucin Expression in Human Airway Mucosa. Am. J. Respir. Cell Mol. Biol. 2007, 37, 606–616. [Google Scholar] [CrossRef] [Green Version]
- Michaeloudes, C.; Bhavsar, P.K.; Mumby, S.; Xu, B.; Hui, C.K.M.; Chung, K.F.; Adcock, I.M. Role of Metabolic Reprogramming in Pulmonary Innate Immunity and Its Impact on Lung Diseases. J. Innate Immun. 2020, 12, 31–46. [Google Scholar] [CrossRef]
- Leopold Wager, C.M.; Wormley, F.L., Jr. Classical versus alternative macrophage activation: The Ying and the Yang in host defense against pulmonary fungal infections. Mucosal Immunol. 2014, 7, 1023–1035. [Google Scholar] [CrossRef] [Green Version]
- Crystal, R.G.; Randell, S.H.; Engelhardt, J.F.; Voynow, J.; Sunday, M.E. Airway Epithelial Cells: Current Concepts and Challenges. Proc. Am. Thorac. Soc. 2008, 5, 772–777. [Google Scholar] [CrossRef]
- van de Veerdonk, F.L.; Gresnigt, M.S.; Romani, L.; Netea, M.G.; Latge, J.P. Aspergillus fumigatus morphology and dynamic host interactions. Nat. Rev. Microbiol. 2017, 15, 661–674. [Google Scholar] [CrossRef]
- Lewis, R.E.; Kontoyiannis, D.P. Invasive aspergillosis in glucocorticoid-treated patients. Med Mycol. 2009, 47 (Suppl. 1), S271–S281. [Google Scholar] [CrossRef]
- Latgé, J.-P. Aspergillus fumigatus and Aspergillosis. Clin. Microbiol. Rev. 1999, 12, 310–350. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lin, K.-T.; Wang, L.-H. New dimension of glucocorticoids in cancer treatment. Steroids 2016, 111, 84–88. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Luvanda, M.; Posch, W.; Vosper, J.; Zaderer, V.; Noureen, A.; Lass-Flörl, C.; Wilflingseder, D. Dexamethasone Promotes Aspergillus fumigatus Growth in Macrophages by Triggering M2 Repolarization via Targeting PKM2. J. Fungi 2021, 7, 70. [Google Scholar] [CrossRef]
- Kadmiel, M.; Cidlowski, J.A. Glucocorticoid receptor signaling in health and disease. Trends Pharmacol. Sci. 2013, 34, 518–530. [Google Scholar] [CrossRef] [Green Version]
- Ehrchen, J.M.; Roth, J.; Barczyk-Kahlert, K. More than Suppression: Glucocorticoid Action on Monocytes and Macrophages. Front. Immunol. 2019, 10, 2028. [Google Scholar] [CrossRef] [Green Version]
- Chandorkar, P.; Posch, W.; Zaderer, V.; Blatzer, M.; Steger, M.; Ammann, C.G.; Binder, U.; Hermann, M.; Hörtnagl, P.; Lass-Flörl, C.; et al. Fast-track development of an in vitro 3D lung/immune cell model to study Aspergillus infections. Sci. Rep. 2017, 7, 1–13. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lescoat, A.; Ballerie, A.; Augagneur, Y.; Morzadec, C.; Vernhet, L.; Fardel, O.; Jégo, P.; Jouneau, S.; Lecureur, V. Distinct Properties of Human M-CSF and GM-CSF Monocyte-Derived Macrophages to Simulate Pathological Lung Conditions In Vitro: Application to Systemic and Inflammatory Disorders with Pulmonary Involvement. Int. J. Mol. Sci. 2018, 19, 894. [Google Scholar] [CrossRef] [Green Version]
- Blom, R.A.M.; Erni, S.T.; Krempaská, K.; Schaerer, O.; Van Dijk, R.M.; Amacker, M.; Moser, C.; Hall, S.R.R.; Von Garnier, C.; Blank, F. A Triple Co-Culture Model of the Human Respiratory Tract to Study Immune-Modulatory Effects of Liposomes and Virosomes. PLoS ONE 2016, 11, e0163539. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kim, B.-Y.; Son, Y.; Lee, J.; Choi, J.; Kim, C.D.; Bae, S.S.; Eo, S.-K.; Kim, K. Dexamethasone inhibits activation of monocytes/macrophages in a milieu rich in 27-oxygenated cholesterol. PLoS ONE 2017, 12, e0189643. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hermanns, M.I.; Unger, R.E.; Kehe, K.; Peters, K.; Kirkpatrick, C.J. Lung epithelial cell lines in coculture with human pulmonary microvascular endothelial cells: Development of an alveolo-capillary barrier in vitro. Lab. Investig. 2004, 84, 736–752. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Millius, A.; Weiner, O.D. Manipulation of neutrophil-like HL-60 cells for the study of directed cell migration. Methods Mol. Biol. 2010, 591, 147–158. [Google Scholar] [PubMed] [Green Version]
- Czock, D.; Keller, F.; Rasche, F.M.; Häussler, U. Pharmacokinetics and Pharmacodynamics of Systemically Administered Glucocorticoids. Clin. Pharmacokinet. 2005, 44, 61–98. [Google Scholar] [CrossRef] [PubMed]
- Cronin, J.; Kennedy, U.; McCoy, S.; Fhailí, S.N.A.; Crispino-O’Connell, G.; Hayden, J.; Wakai, A.; Walsh, S.; O’Sullivan, R. Single dose oral dexamethasone versus multi-dose prednisolone in the treatment of acute exacerbations of asthma in children who attend the emergency department: Study protocol for a randomized controlled trial. Trials 2012, 13, 141. [Google Scholar] [CrossRef] [Green Version]
- Wang, J.; Wang, C.; Li, X.; Kong, L.; Gao, K.; Liu, R.-Y. The effects of anti-asthma drugs on the phagocytic clearance of apoptotic eosinophils by A549 cells. Respir. Med. 2009, 103, 1693–1699. [Google Scholar] [CrossRef] [Green Version]
- Xiang, L.; Hu, Y.-F.; Wu, J.-S.; Wang, L.; Huang, W.-G.; Xu, C.-S.; Meng, X.-L.; Wang, P. Semi-Mechanism-Based Pharmacodynamic Model for the Anti-Inflammatory Effect of Baicalein in LPS-Stimulated RAW264.7 Macrophages. Front. Pharmacol. 2018, 9. [Google Scholar] [CrossRef]
- Lass-Flörl, C.; Roilides, E.; Löffler, J.; Wilflingseder, D.; Romani, L. Minireview: Host defence in invasive aspergillosis. Mycoses 2013, 56, 403–413. [Google Scholar] [CrossRef]
- Drummond, R.A.; Swamydas, M.; Oikonomou, V.; Zhai, B.; Dambuza, I.M.; Schaefer, B.C.; Bohrer, A.C.; Mayer-Barber, K.D.; Lira, S.A.; Iwakura, Y.; et al. CARD9+ microglia promote antifungal immunity via IL-1β- and CXCL1-mediated neutrophil recruitment. Nat. Immunol. 2019, 20, 559–570. [Google Scholar] [CrossRef]
- Alonso, M.F.; Gow, N.A.R.; Erwig, L.P.; Bain, J.M. Macrophage Migration Is Impaired within Candida albicans Biofilms. J. Fungi 2017, 3, 31. [Google Scholar] [CrossRef] [Green Version]
- Byrne, A.J.; Mathie, S.A.; Gregory, L.G.; Lloyd, C.M. Pulmonary macrophages: Key players in the innate defence of the airways. Thorax 2015, 70, 1189–1196. [Google Scholar] [CrossRef] [Green Version]
- Zhang, Z.; Wang, Z.; Ren, H.; Yue, M.; Huang, K.; Gu, H.; Liu, M.; Du, B.; Qian, M. P2Y6 Agonist Uridine 5′-Diphosphate Promotes Host Defense against Bacterial Infection via Monocyte Chemoattractant Protein-1–Mediated Monocytes/Macrophages Recruitment. J. Immunol. 2011, 186, 5376–5387. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Parker, D.; Prince, A. Innate Immunity in the Respiratory Epithelium. Am. J. Respir. Cell Mol. Biol. 2011, 45, 189–201. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kai, H.; Yoshitake, K.; Hisatsune, A.; Kido, T.; Isohama, Y.; Takahama, K.; Miyata, T. Dexamethasone suppresses mucus production and MUC-2 and MUC-5AC gene expression by NCI-H292 cells. Am. J. Physiol. Cell. Mol. Physiol. 1996, 271 Pt 1, L484–L488. [Google Scholar] [CrossRef]
- Kitano, M.; Ishinaga, H.; Shimizu, T.; Takeuchi, K.; Majima, Y. Effects of Clarithromycin and Dexamethasone on Mucus Production in Isografted Rat Trachea. Pharmacology 2011, 87, 56–62. [Google Scholar] [CrossRef]
- Leiva-Juárez, M.M.; Kolls, J.K.; Evans, S.E. Lung epithelial cells: Therapeutically inducible effectors of antimicrobial defense. Mucosal Immunol. 2018, 11, 21–34. [Google Scholar] [CrossRef] [PubMed]
- Workman, A.D.; Cohen, N.A. The Effect of Drugs and Other Compounds on the Ciliary Beat Frequency of Human Respiratory Epithelium. Am. J. Rhinol. Allergy 2014, 28, 454–464. [Google Scholar] [CrossRef]
- Wasylnka, J.A.; Moore, M.M. Uptake of Aspergillus fumigatus Conidia by Phagocytic and Nonphagocytic Cells In Vitro: Quantitation Using Strains Expressing Green Fluorescent Protein. Infect. Immun. 2002, 70, 3156–3163. [Google Scholar] [CrossRef] [PubMed] [Green Version]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Antibody/Stain | Source | Cat Number | Concentration Applied |
|---|---|---|---|
| Alexa Fluor® 488 anti -human CD40 | Biolegend, San Diego, CA | 334318 | 40 µg/mL |
| Alexa Fluor® 488 Wheat Germ Agglutinin | Biotium, Hayward, CA | 29022-1 | 5 µg/mL |
| Anti-Pan Cytokeratin | Invitrogen, Carlsbad, CA | 53-9003-82 | 0.5 mg/mL |
| Alexa Fluor® 647 anti -human CD163 | Biolegend, San Diego, CA | 326508 | 10 µg/mL |
| Hoechst 33342 | Sigma, St. Louis, MO | B1155 | 2 µg/mL |
| Mitotracker® Orange | Invitrogen, Carlsbad, CA | M7511 | 100–500 nM |
| Mouse anti-E-cadherin Alexa Fluor® 647 | BD Pharmingen | 560062 | 2.5 µg/mL |
| Mouse anti-Occludin Alexa Fluor® 594 | Invitrogen, Carlsbad, CA | 331594 | 5 µg/mL |
| MUC 1 488 | Abcam, Cambdridge, UK | Ab196443 | 0.5 mg/mL |
| MUC 5 AC 555 | Abcam, Cambdridge, UK | Ab218714 | 0.5 mg/mL |
| PE anti-human PKM2 (D78A4) | Cell Signaling Technology, Danvers, MA | 983675 | 2.5 µg/mL |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Luvanda, M.K.; Posch, W.; Noureen, A.; Lafon, E.; Zaderer, V.; Lass-Flörl, C.; Wilflingseder, D. Dexamethasone Creates a Suppressive Microenvironment and Promotes Aspergillus fumigatus Invasion in a Human 3D Epithelial/Immune Respiratory Model. J. Fungi 2021, 7, 221. https://0-doi-org.brum.beds.ac.uk/10.3390/jof7030221
Luvanda MK, Posch W, Noureen A, Lafon E, Zaderer V, Lass-Flörl C, Wilflingseder D. Dexamethasone Creates a Suppressive Microenvironment and Promotes Aspergillus fumigatus Invasion in a Human 3D Epithelial/Immune Respiratory Model. Journal of Fungi. 2021; 7(3):221. https://0-doi-org.brum.beds.ac.uk/10.3390/jof7030221
Chicago/Turabian StyleLuvanda, Maureen K., Wilfried Posch, Asma Noureen, Eliott Lafon, Viktoria Zaderer, Cornelia Lass-Flörl, and Doris Wilflingseder. 2021. "Dexamethasone Creates a Suppressive Microenvironment and Promotes Aspergillus fumigatus Invasion in a Human 3D Epithelial/Immune Respiratory Model" Journal of Fungi 7, no. 3: 221. https://0-doi-org.brum.beds.ac.uk/10.3390/jof7030221