
Fungal Melanin and the Mammalian Immune System


1
Department of Medicine, Albert Einstein College of Medicine, Bronx, NY 10461, USA
2
Department of Microbiology, Faculty of Medicine, Chiang Mai University, Chiang Mai 50200, Thailand
3
Department of Microbiology and Immunology, Albert Einstein College of Medicine, Bronx, NY 10461, USA
*
Author to whom correspondence should be addressed.
J. Fungi 2021, 7(4), 264; https://0-doi-org.brum.beds.ac.uk/10.3390/jof7040264
Submission received: 22 February 2021
/
Revised: 25 March 2021
/
Accepted: 26 March 2021
/
Published: 31 March 2021
(This article belongs to the Special Issue Antifungal Immunity and Fungal Vaccine Development)
Abstract
:Melanins are ubiquitous complex polymers that are commonly known in humans to cause pigmentation of our skin. Melanins are also present in bacteria, fungi, and helminths. In this review, we will describe the diverse interactions of fungal melanin with the mammalian immune system. We will particularly focus on Cryptococcus neoformans and also discuss other major melanotic pathogenic fungi. Melanin interacts with the immune system through diverse pathways, reducing the effectiveness of phagocytic cells, binding effector molecules and antifungals, and modifying complement and antibody responses.
1. Introduction
Melanins are a family of structurally complex dark pigment polymer present in all biological kingdoms [1]. The polymer is made up of covalently linked indoles, but the overall structure is granular; however, detailed structures are not well characterized because it is a collection of polymers with mixed pre-indole structures [2]. Moreover, melanins are amorphous and are not suitable for study by crystallography or cryo-electron microscopy, which has led to complicated efforts to solve the structure of these natural pigments. Mammalian melanin biogenesis happens by oxidation of L-tyrosine via tyrosinase in melanocytes, which are neural crest-derived dendritic cells [3,4]. However, melanins in bacteria, fungi, and helminths are produced through the polyketide synthase (PKS) pathway or catalyzed by phenoloxidase [5]. This review will focus on melanin in fungi and its function in pathogen–host interaction with a particular emphasis on the immune system.
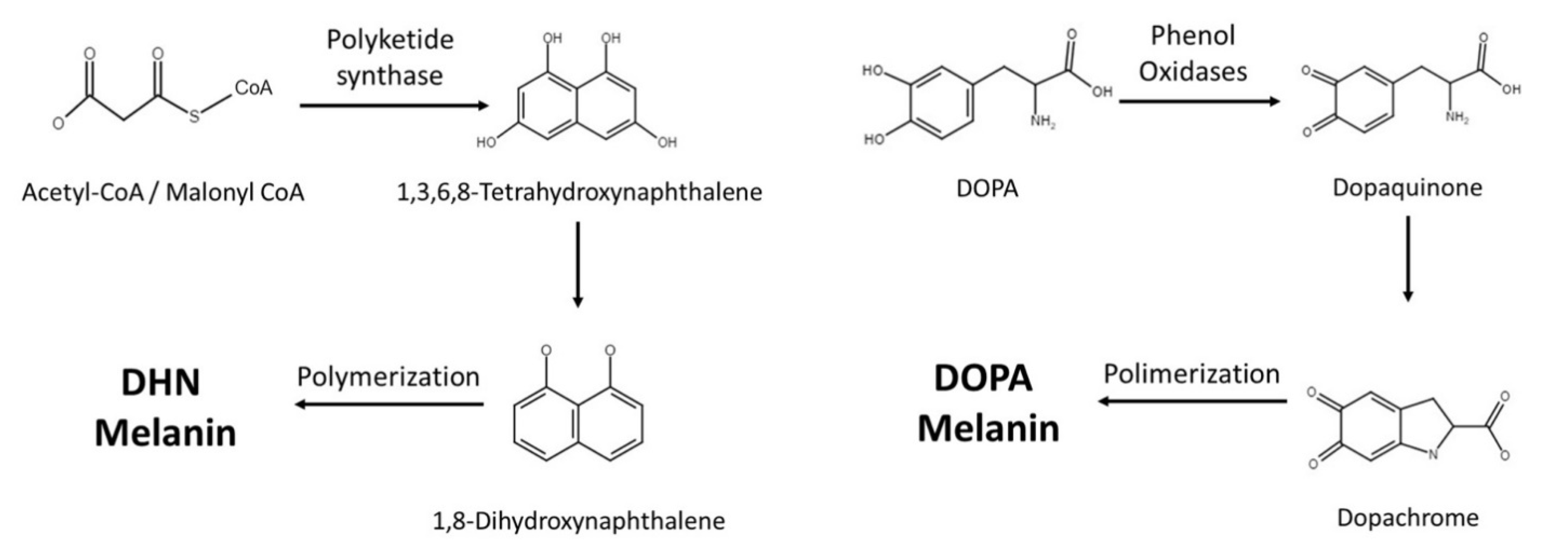
2. Melanin Synthesis
Fungi synthesize melanin via two main pathways, namely 1,8-dihydroxynaphthalene (DHN) and l-3,4-dihyroxyphenylalanine (L-DOPA) (Figure 1 and Table 1). In the DHN pathway, 1,3,6,8-tetrahydroxynaphthalene (1,3,6,8-THN) is first synthesized through PKS, which is a multi-domain enzyme complex that produces polyketides. Polyketides are products derived from acetyl-CoA or propionyl-CoA with malonyl-CoA or methylmalonyl-CoA. The condensation reactions are driven by decarboxylation, yielding a beta-keto functional group [6]. This is then followed by reduction and dehydration reactions that eventually produce DHN. It is the polymerization of DHN that leads to the formation of melanin [5,7,8]. Polyketides are a class of secondary metabolites mainly produced in bacteria, fungi, and plants, and they serve very different purposes in our society [9]. Polyketides such as macrolide, tetracycline, and amphotericin antimicrobials serve tremendous value, while aflatoxin can be lethal to mammals [10,11].
The L-DOPA pathway is similar to mammalian melanin biosynthesis, where the pathway typically uses either L-DOPA or tyrosine as starting molecules. If the pathway starts with tyrosine, tyrosinase will perform a two-step oxidation, turning tyrosine into dopaquinone. Similarly, laccase is the enzyme responsible for converting L-DOPA into dopaquinone. Dopaquinone is then turned into leucodopachrome (cyclodopa) and then oxidized to dopachrome. Dopachrome then goes through tautomerization to form dihydroxyindoles, which are simultaneously oxidized and polymerized to produce DOPA melanin [8].
3. Cryptococcus neoformans
C. neoformans is one of the most well studied pathogens for melanization (Figure 2). Cryptococcus neoformans is unique among pathogenic fungi as it solely relies on the L-DOPA pathway and requires exogenous phenolic substrates to form melanin. After the polysaccharide capsule, melanin is the second most important virulence factor in C. neoformans as it has been calculated as contributing to 14% of the pathogen’s total virulence [16]. This notable aspect of cryptococcal melanization has led to extensive study of this polymer in Cryptococcus spp. [17]. Other species can produce melanin using endogenous compounds or exogenous substrates, and some can produce more than one type of melanin [12,18,19].
In the early 1980s, Kwon-Chung et al. produced melanin-deficient strains (Mel−) via UV irradiation and observed that the mutants lacked virulence as the cells were cleared from mouse organs, whereas wild-type (WT) cells expanded in numbers, especially in the brain [21]. The same study showed that Mel− mutants were defective in the active transport system for diphenolic compounds and phenoloxidase, which is the first enzyme needed in the L-DOPA melanogenesis pathway. A subsequent study confirmed that the loss of phenoloxidase activity was responsible for the Mel− mutant phenotype [22]. Williamson next discovered that laccase, a phenoloxidase, was encoded by the lac1 gene [23]. Laccase was linked to C. neoformans virulence in vivo through disruption of the 5′ end of the lac1 gene [24]. Interestingly, C. neoformans has two laccase genes, where lac2 is 75% similar to lac1, but basal transcript levels of lac2 are much lower, and mutation of the gene only induced a mild delay in melanin formation [25].
3.1. Melanosomes and Melanin Location
Melanin synthesis needs to occur in a contained environment as its intermediates are reactive to other surrounding substances [26]. Melanin is synthesized in melanosomes within human melanocytes. In C. neoformans, melanization occurs in specialized vesicles that are also called melanosomes [27]. In C. neoformans, laccase was found to be one of many products packaged into vesicles that could be trapped in the cell wall or secreted to the extracellular milieu [28]. Indeed, RNA interference (RNAi) of sec6, which encodes a protein required for an event leading to fusion of post-Golgi vesicles with the plasma membrane, prevented melanization by blocking vesicle access [29]. Internal “melanosomes” are also observed in C. albicans, Cladosporium carrionii, and Hormoconis resinae [14,30]. Work in Aspergillus has revealed that disruption of endosomal sorting blocks the deposition of melanin in the cell wall, further supporting non-conventional secretory mechanisms driving proper polymer formation and draws marked similarities to mammalian melanosome biology [31]. The existence of cryptococcal melanosomes explains how melanin is distributed in the cell wall, with the melanosomes depositing diffusely in ring-like layers in the cell wall [19,32]. However, melanin’s distribution is pathogen-dependent. Whereas C. neoformans has melanin located in the innermost layer of the cell wall [33], melanin is located on the surface or outer wall layer in Candida albicans, Aspergillus spp., Paracoccidioides spp., Coccidioides spp., Histoplasma capsulatum, and Sporothrix schenckii [13,34,35,36,37,38].
3.2. Melanin and Cell Wall
Melanin’s role in the cell wall has been extensively studied in C. neoformans. Many of these studies utilized melanin “ghosts”, which are melanin spheres left over from enzymatic digestion and chemical breakdown of the cell wall. Ghosts formed in the presence of 13 C showed that melanin is an integrated part of the cell wall, forming covalent bonds with polysaccharides within the cell wall [26]. C. neoformans melanin ghosts are made of two to five concentric layers with thicknesses ranging from 50 to 75 nm per layer. The layers become thicker as cultures become older [39]. The presence of melanin impacts the cell wall’s properties. For example, a comparison of WT melanized and non-melanized A. fumigatus mutants showed that the conidial wall of the reference strain was composed of several superimposed layers with a thick electron transparent inner layer and two thin electron-dense outer layers, while mutant conidia showed a cell wall devoid of the outermost layer. Scanning electron microscopy (SEM) observed that pigment-less mutants have smooth-walled conidia without ornamentation, and further analysis showed that the mutant cell wall was also less electronegative and hydrophobic [40]. Interestingly, there are still pores present in the cell wall after melanization, which permit the transport of molecules across this barrier. In C. neoformans, pore sizes were markedly reduced in pigmented cells as pore radii were estimated to be 4 and 10.6 nm for melanized and non-melanized types, respectively [41]. C. neoformans cell wall pore size also correlated with the age of the culture, as day 4 melanin ghosts displayed the greatest porosity at 41 μL Å−1 g−1 at 16.4 Å, while day 10 melanin ghosts’ porosity was 15 μL Å−1 g−1 at 10 Å. The same study also showed that an antibody against melanin decreased the total porosity from 16 to 8 μL Å−1 g−1 [39], which has important implications for drug targeting.
In terms of its structure within the cell wall, C. neoformans melanin is described as being composed of irregular granules 50–80 nm in diameter, and similar granular structures are observed in other fungi such as Fonsecaea pedrosoi, Hortaea werneckii, and Agaricus bisporumand [39,42,43,44]. In C. neoformans, these granules are cross-linked with chitin, which is a polymer of β(1,4)-linked N-acetylglucosamine (GlcNAc) subunits joined in antiparallel chains by hydrogen bonding to produce strong microfibrils [45]. Chitin is synthesized by chitin synthase (CHS), which uses UDP-N-GlcNAc as a substrate to grow the polymer [32].
Through chs-deficient mutants, we were able to learn how chitin interacts with melanin in Wangiella dermatitidis, C. neoformans, and Candida albicans. In W. dermatitidis, the WdCHS4 knockout mutant allowed us to observe a decrease in melanin deposition within the cell wall concomitant with an increase in the pigment’s concentration extracellularly. In C. neoformans, a similar observation occurred in cda1, cda2, and cda3 deletion mutants. These genes are responsible for the deacetylation of chitin, producing chitosan. Chitosan not only helps in cell integrity and bud separation, but the disruption of these genes resulted in a “leaky melanin” phenotype that exhibited melanin suspended in the supernatant, while still retaining some melanin within the cell [46]. The chs3 and csr2 genes, which are a chitin synthase and a regulator of chitin synthase, respectively, were also found to be necessary to retain melanin within the cell as their mutants also demonstrated the “leaky” phenotype [47,48]. C. neoformans cda1∆2Δ3Δ mutants have decreased virulence, and mice inoculated with this mutant generate a strong inflammatory response followed by enrichment of Th1-type T cells in lung tissue, producing immunity against the WT strain [49].
C. albicans did not exhibit these leaky phenotypes but displayed repressed melanin externalization, and seemingly different structural types of chitin have different melanin localization effects. The chitin synthase (chs2)-deficient mutants showed accumulation of melanin grains and melanosomes within the cells, while chs8- and chs8/chs2-deficient mutants displayed WT behavior [14]. Similarly, melanin externalization was impaired with disruptions in the chitinase genes CHT2 and CHT3 or the chitin pathway regulator ECM22.
3.3. Survival Advantage
Melanin production enhances fungal survival in the environment. Facultative melanotic C. neoformans produces melanin while dwelling in the soil [50]. The polymer absorbs electromagnetic radiations such as UV and gamma radiation, much like its role in humans, and allows fungi to survive in temperature extremes [51,52]. It is also partially responsible for the survival of mold inside nuclear reactors [53,54]. Melanin not only protects against toxic radiation, but the polymer also enables fungal cells to harvest high-energy radiation for growth [55,56], which means that fungi are limited autotrophs. The primary mechanism for melanins being so efficient with shielding from biotic and abiotic factors relies not only on its extremely stable structure but also on the fact that it has a remarkably broad optical absorption, paramagnetism, and charge transport. These physico-chemical properties are since, as polyphenols, melanins are polymers composed by series of aromatic rings, thus allowing electron resonance and also mediating energy transfer reactions in the cells [57].
Cells with melanin are structurally stronger than non-pigmented cells as melanized fungi are more resistant to enzymatic degradation [58]. The carboxyl, phenolic, hydroxyl, and amine groups within the polymer compound are perfect sites for heavy metal binding [59], and the ability of melanized C. neoformans to resist silver compounds is linked to its binding capacity [60].
Melanin alters antimicrobial pharmacokinetics. In vitro efficacies of various antibiotics (aminoglycosides, tetracyclines, and vancomycin) and antifungals (amphotericin B and caspofungin) are significantly altered in the presence of melanin [61,62,63]. In fact, according to Barza et al., tobramycin’s efficacy decreased by 80% in the presence of melanin [62]. Binding, but no efficacy reduction, is seen with fluoroquinolones, penicillins, cephalosporins, and azoles [61,63].
3.4. Melanin and Host Effector Cells
Melanin is a negatively charged polymer [64]. Melanization increased the overall cellular negative charge by 3 to 33% in nine different encapsulated strains of C. neoformans and by 86% in an acapsular strain [65]. It has been hypothesized that cell charge might affect phagocytosis. Indeed, phagocytosis of C. neoformans can be affected by the surface hydrophobicity and charge [66]. Microbial cell charge has been shown to affect rates of phagocytosis by neutrophils and monocytes [67,68]. In fact, melanization of C. neoformans interferes with phagocytosis in vivo [69].
The mere presence of melanin, regardless of charge, improves pathogen survival. Melanized and non-melanized mutants were incubated with capsule-binding monoclonal Mab 2H1 antibodies and murine macrophages. Melanized cells conferred a 31% survival advantage over the mutant with 98.5% of the cells surviving [70]. The authors attributed the resistance to phagocytosis to melanin’s resistance to reactive oxygen species (ROS), but later research suggests that more complicated mechanisms are at play, and some seem to be pathogen-specific. Melanin’s negative charge and capacity to bind diverse molecules make cryptococcal cells less susceptible to cationic antimicrobial peptides released by phagocytes, such as defensins, than non-melanized cells [71]. C. neoformans is able to escape macrophages via nonlytic exocytosis [72]. Remarkably, laccase expression regulates both melanin formation and rates of nonlytic exocytosis [73].
Melanin-producing strains of Cryptococcus spp. suppress the host immune and inflammatory responses [74,75]. Rosas et al. induced granuloma formation in mouse peritoneal cavity after injecting isolated C. neoformans melanin particles. Granulomas were seen in the liver, spleen, and lung. Granuloma formation around melanin particles resembled a foreign body reaction [76]. This might be due to the immune system’s inability to break down foreign melanin. In chronic and latent cryptococcal infections, the yeast cells are usually encased in granulomas, similarly to those observed with melanin particles [77]. Interestingly, fungal pathogens that can induce latency and granuloma formation produce melanin [74]. This suggests that melanin might have immunomodulatory effects, preventing the pathogens’ clearance. In fact, there is early evidence that laccase protects C. neoformans from alveolar macrophages antifungal activities [78].
Since granulomas contain a wide range of macrophage morphologies, there have been numerous investigations focused on macrophages [79]. Indeed, Tajima et al. found that solubilized melanin suppresses macrophage function [80]. Solubilized melanin did not affect the survival of the macrophages, but, interestingly, induced proliferation at a very high concentration, 100 µg·mL−1. The macrophage’s phagocytosis ability was attenuated significantly, along with ROS production. Regarding cytokine production, TNF-a, IL-1b, and IL-6 were suppressed [80]. Interestingly, another study did not find differences in IL-2, IL-10, TNF-α, and IFN-γ levels, while there was twice the amount of IL-4 and an elevated monocyte chemoattractant protein-1 (MCP-1) level in lung tissues infected with melanized versus non-melanized C. neoformans [69].
Not only does melanization influence macrophage activities, but melanin also modulates T cell responses. C. neoformans strains 52 and 145 have opposite inflammatory reactions in experimental mouse models and these strains have been used to dissect out adaptive immune processes in cryptococcosis. The two strains differ in that strain 145 produces more melanin and strain 52 produces a stronger pulmonary inflammatory response [81]. In a pulmonary infection model, strain 145 had delayed responses from neutrophils and macrophages and a diminished lymphocyte response, especially CD4+ T cells, compared to strain 52. The lack of CD4+ T cell proliferation was also seen in peripheral lymph nodes [82]. Accordingly, alveolar macrophages produced 70% less TNF-α when they were incubated with the high melanin-producing strain 145 versus strain 52. These changes are also present during murine central nervous system infections. The high melanin-producing strain suppressed IL-12, IL-1β, TNF-α, IFN-γ, and inducible nitric oxide synthase (iNOS) to almost non-existent levels and resulted in a 100% mortality rate [83].
3.5. Antibody
Immunization of mice with cryptococcal melanin induces a robust antibody response with significant increases in IgM and IgG levels [84]. Moreover, antibodies to melanin naturally occur during cryptococcosis [85]. The development of cryptococcal melanin-binding monoclonal antibodies (mAbs) accelerated the study of melanin in pathogenic fungi [20], particularly as the mAbs reacted with diverse biological and synthetic melanins [35]. The melanin-binding mAbs revealed a new mechanism for antibody activity in which the mAbs interfere with properties of the polymer to inhibit growth [86]. In fact, mAbs block pores in the layered melanin [39], which likely interferes with the transport of macromolecules. Melanin-binding mAbs also prolonged the survival of mice lethally infected with C. neoformans [86].
3.6. Complement System
The complement system is a major effector arm of host defense. Complement C3 fragments are deposited in the C. neoformans capsule and enhance the clearance of these cells by phagocytes [87,88]. Interestingly, C3 also deposits on cryptococcal melanin ghosts, and this process is due to activation of the alternate complement pathway [76]. Deposition of complement fragments also occurred with ghosts injected into the lungs of mice. Notably, the presence of melanin did not affect the kinetics of C3 binding to the cryptococcal capsule. However, the C3-labeled melanin ghosts induced the formation of small granulomas in mouse lungs.
4. Aspergillus fumigatus
A. fumigatus is another pathogenic fungus that undergoes melanization, and pigment formation in this species has been the subject of extensive investigation. 1,8-dihydroxynaphthalene (DHN) melanin is the major melanin found in A. fumigatus [89]. A set of six genes are needed for melanin synthesis: pksP, ayg1, arp2, arp1, arb1, and arb2, ordered from upstream to downstream [90]. Melanin from A. fumigatus has many similarities to melanin from C. neoformans, as both provide protection from ultraviolet light and scavenge ROS generated by phagocytes [91]. Albino conidia incubated with phagocytes induced a 10-fold increase in ROS production compared with WT, which suggests that the melanin increased ROS scavenging abilities in the WT conidia. Moreover, the albino mutants were more effectively killed by monocytes than wild-types [92].
Melanin regulates host pro-inflammatory cytokine responses by physically masking fungal pathogen-associated molecular patterns (PAMPs) from immune recognition such as that observed by the rodlets layer. Chai et al. demonstrated that albino conidia were able to generate much higher IL-6, IL-10, and TNF-α levels compared to WT. IL-6 and IL-10 had roughly 12-fold and 5-fold higher levels, respectively. The authors pinpointed that albino conidia had β-glucan and other PAMPs such as mannans more readily available to bind to dectin-1, Toll-like receptor 4 (TLR4) and Mannose receptors on peripheral blood mononuclear cells (PBMCs) [93]. As conidia mature, they swell and germinate, and this process exposes β-1,3-glucan on their surface, which induces an immune response [94].
Jahn et al. observed that ΔpksP melanin-deficient conidia were more effectively phagocytosed and killed by macrophages when compared with WT conidia [95]. Thywissen et al. next demonstrated that the albino ΔpksP mutant conidia had a 3.5-fold increase in vacuolar-type ATPase-dependent phagosomal acidification observed in alveolar and monocyte-derived macrophages (~20% acidified conidia vs. 70%). There were virtually no acidified phagosomes containing WT conidia in granulocytes, but ~50% in the mutant group. The pH within the mutant-containing phagolysosome was 5, while the WT had a pH of 6, and the lower pH results in more effective actions by phagosomal enzymes. Interestingly, synthetic DOPA melanin did not prevent acidification, only DHN melanin did [96]. However, among Aspergillus spp., A. flavus was the most effective in suppressing acidification, closely followed by A. fumigatus. Microtubule-associated protein 1A/1B-light chain 3 (LC3)-associated phagocytosis (LAP) is a non-canonical autophagy pathway that is linked to certain pattern recognition receptors that trigger phagosome formation [97]. A. fumigatus melanin inhibits calcium-calmodulin signaling on the protein Rubicon, a key regulator in the LAP pathway [98,99], and Rubicon directly interacts with the p22phox subunit and facilitates NADPH oxidase activation during phagocytosis [100]. Melanin ultimately blocks the p22phox NADPH oxidase subunit from localizing on the phagosome membrane, thus blocking the assembly of the oxidase complex. Melanin’s effect on the NADPH oxidase complex is conserved in A. nidulans as well [101].
The effects of melanin vary on different immune cells. Dendritic cells (DCs) are not stimulated by A. fumigatus melanin. Bayry et al. demonstrated that DCs failed to produce cytokines TNF-α, IL-1β, IL-6, and IL-10 in response to melanized A. fumigatus conidia, and DCs treated with WT melanin ghost also failed to activate T cells [102]. The group interestingly found that ΔpksP, Δayg1, and Δarp2 mutants increased different amounts of acetyl-CoA, malonyl-CoA, and 1,3,6,8-THN, respectively. These mutants displayed altered cell walls with unmasked surface structures and were able to activate DCs.
Mammals have evolved various systems to combat melanized fungal pathogens. On the surface of mouse endothelial cells, there is a melanin-sensing C-type lectin receptor (MelLec) that recognizes DHN melanin in the conidial spores of A. fumigatus and other DHN-melanized fungi, such as Cladosporium cladosporioides and Fonsecaea pedrosoi [103]. The expression of this C-lectin is essential for protection against disseminated aspergillosis, and albino mutants are not recognized by the receptor. Macrophages change their metabolism in the presence of melanin, especially glycolysis metabolism, which is required for defense against Aspergillus. Goncalves et al. elucidated that DHN melanin blocks endoplasmic reticulum calcium/calmodulin signaling, which activates glycolysis and mammalian target of rapamycin (mTOR)-mediated defense against Aspergillus conidia [104]. Consequentially, this impairment of glycolysis, mediated by mammalian target of rapamycin (mTOR) and hypoxia-inducible factor 1 subunit alpha (HIF-1α), decreases macrophages’ conidicidal ability by lowering ROS concentration and inflammatory cytokines IL-1β, IL-6, IL-17A, TNF-α, and IFN-γ production [105].
A. fumigatus can be cleared by activating the complement system [106,107]. Tsai et al. further demonstrated that disruption of the arp1 gene leads to increased C3 deposition on the conidial cell surface [108]. Similarly, disrupting alb1/pksP that encodes polyketide synthase resulted in a significant increase in C3 binding on conidial surfaces, with an expected increase in phagocytosis by neutrophils and a decrease in virulence [89,109]. Direct binding of C3 fragments in normal human serum has been shown with A. niger melanin [76].
5. Other Melanotic Fungi and Their Interactions with the Immune System
Fonseca spp. are causative agents of chromoblastomycosis, and these fungi produce large quantities of melanin. Melanized Fonsecaea monophora and cell wall-containing extracted melanin significantly decrease the expression of inducible nitric oxide synthase gene and the production of nitric oxide and enhanced non-protective Th2 responses [110]. Macrophages infected with pigmented F. monophora enhanced the differential expression of genes related to immune responses, including the MAPK signaling pathway, demonstrating how melanization modifies pathogenesis [111]. As with C. neoformans and A. niger melanin, melanized Fonsecaea pedrosoi was quickly labeled with C3, C4, and C9 complement components [112].
Several endemic dimorphic pathogenic fungi produce melanin, including Histoplasma capsulatum [38], Paracoccidoides spp. [36], Coccidioides immitis [37], Blastomyces dermatitidis [15], and Talaromyces marneffei [113]. Melanin production is associated with pathogenesis in Paracoccidioides spp. through complex processes that extend beyond pigment production. Different Paracoccidioides species resist phagocytosis of yeast cells by macrophages, and this effect is associated with the degree of melanization in each strain [114,115]. Melanized P. brasiliensis is also highly resistant to NO, ROS, hypochlorite, and H2O2 [116]. A recent proteomic analysis comparing melanized and non-melanized P. brasiliensis and P. lutzii revealed that melanization leads to an abundance of virulence-associated proteins, including heat-shock proteins, vesicular transport proteins, adhesins, superoxide dismutases, proteases, and phospholipases, which further underscores the complex mechanisms that occur along with melanin production to subvert the host [117]. As with cryptococcosis, melanin-binding antibodies are generated during murine as well as human infection with P. brasiliensis [114].
T. marneffei (formerly Penicillium marneffei) also utilizes melanin to avoid host defenses. Remarkably, the T. marneffei genome has 23 polyketide synthase genes and additional non-ribosomal polyketide synthase hybrid genes [118]. Mutants unable to form the polymer are more sensitive to antifungals, H2O2, and sodium dodecyl sulfate (SDS). Furthermore, melanized cells were significantly more resistant to phagocytosis and killing compared to melanin-deficient mutants [119]. A second study corroborated the capacity of melanized fungal cells to resist antifungals [120]. There is an interesting link to melanin in T. marneffei with tyrosine catabolism, which is essential for survival in the host cells [121].
Melanin is well described in Sporothrix spp., and the production of the polymer is closely linked to virulence [13,122,123]. Melanization of S. globosa leads to a reduction in antigen presentation by macrophages and facilitates the dissemination of the pathogen [124]. Melanin formation has been linked to increased dissemination in several Sporothrix species [125]. However, melanization of some S. schenckii strains may, instead, induce the formation of granuloma, which facilitates the survival of the yeast [126]. Melanin production in Sporothrix complex species is protective against diverse antifungal compounds [127,128].
Melanin is purported to be a major factor in the pathogenicity of Mucorales spp. A recent paper from the Ibrahim laboratory identified compounds that could selectively inhibit eumelanin production by Rhizopus sp. [129]. Moreover, the inhibition of melanin by one blocking compound, UOSC-2, led to the formation of spores that were more efficiently phagocytosed and killed in mouse lungs compared to melanized spores, and the albino spores were similarly more efficiently killed by human macrophages, verifying the importance of melanin in protection against host effector responses against this pigmented species.
6. Open Questions in Melanin Biology
The enigmatic polymer’s structure remains elusive. Although there are synthetic melanins and certain precursors may mimic aspects of melanin’s activities, the complexities of melanin’s structure suggest that these are poor surrogates and do not represent the different forms of melanin. In fact, melanin formation in vivo is markedly different from that generated in vitro [130,131]. Future dissection of the structural unit of melanin, as approached by Dr. Stark and Dr. Cassadevall, will continue to unravel the mystery of melanin’s form [132]. Moreover, differences between types of melanin may reveal important variables in their biology as well as which molecules are combined with their formation and scaffolding. The evolution of different melanin systems also remains mysterious—why do some fungi have a single mechanism for forming melanin and others different avenues for formation? How did this evolve differently in human pathogenic fungi versus environmental species, and did this contribute to their abilities to cause diseases?
Although the formation of melanin is mysterious, its degradation is similarly unknown. The capacity for fungal melanin to persist in tissues even after the pathogen has been killed can challenge the host’s responses to nearby viable cells. Additionally, the effect of fungal melanin on other immune and non-immune cells is still elusive and should be experimentally addressed. Melanin has been suggested to have applications that could impact diverse aspects of life, ranging from remediation of nuclear contamination [133] to space travel [134].
7. Summary
Melanin plays a major role in the pathogenesis of a wide variety of human pathogens such as A. fumigatus, C. neoformans, A. fumigatus, and dimorphic fungi such as Histoplasma capsulatum, Paracoccidioides spp., Coccidioides immitis, and Blastomyces dermatitidis. The polymer can be compared to a Swiss army knife as fungi use melanin to improve their survival in a variety of environments through an array of mechanisms. Since its discovery, our view on melanin has gradually evolved. Melanin not only helps fungi to resist radiation and enzymatic degradation in the environment, but it also helps fungi to wreak havoc in the human body. Melanin renders our antifungals less effective while preventing our innate immune system from actively clearing the fungal invaders by lowering ROS production and phagocytosis. In A. fumigatus, immune evasion also involves modulating dendritic cells. Though melanin seems like an all-powerful tool, our immune system has also created our own counter measures such as the MelLec (melanin-sensing C-type lectin receptor). Meanwhile, our macrophages are also tuned in to detect melanin. It is exciting to see what the future holds as we unravel more about the interplay between melanin and our immune system, which will likely lead to the discovery of new therapeutic targets to aid us in our ongoing struggle with pathogenic fungi.
Author Contributions
Conceptualization, J.D.N.; writing—original draft preparation, S.L.; writing—review and editing, S.Y., D.Z.-M., J.D.N. funding acquisition, J.D.N. All authors have read and agreed to the published version of the manuscript.
Funding
J.D.N. was supported in part by NIH AI52733. S.Y. was supported by a research grant from Chiang Mai University, Chiang Mai, Thailand.
Institutional Review Board Statement
Not applicable.
Informed Consent Statement
Not applicable.
Data Availability Statement
Not applicable.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Suwannarach, N.; Kumla, J.; Watanabe, B.; Matsui, K.; Lumyong, S. Characterization of melanin and optimal conditions for pigment production by an endophytic fungus, Spissiomyces endophytica SDBR-CMU319. PLoS ONE 2019, 14, e0222187. [Google Scholar] [CrossRef] [Green Version]
- Riley, P.A. Melanin. Int. J. Biochem. Cell Biol. 1997, 29, 1235–1239. [Google Scholar] [CrossRef]
- Eisen, T.G. The control of gene expression in melanocytes and melanomas. Melanoma Res. 1996, 6, 277–284. [Google Scholar] [CrossRef]
- Sánchez-Ferrer, Á.; Neptuno Rodríguez-López, J.; García-Cánovas, F.; García-Carmona, F. Tyrosinase: A comprehensive review of its mechanism. Biochim. Biophys. Acta (BBA) Protein Struct. Mol. Enzymol. 1995, 1247, 1–11. [Google Scholar] [CrossRef]
- Wheeler, M.H.; Bell, A.A. Melanins and their importance in pathogenic fungi. In Current Topics in Medical Mycology; Springer: Berlin/Heidelberg, Germany, 1988; pp. 338–387. [Google Scholar]
- Staunton, J.; Weissman, K.J. Polyketide biosynthesis: A millennium review. Nat. Prod. Rep. 2001, 18, 380–416. [Google Scholar] [CrossRef]
- Butler, M.; Day, A. Fungal melanins: A review. J. Can. J. Microbiol. 1998, 44, 1115–1136. [Google Scholar] [CrossRef]
- Langfelder, K.; Streibel, M.; Jahn, B.; Haase, G.; Brakhage, A.A. Biosynthesis of fungal melanins and their importance for human pathogenic fungi. Fungal Genet. Biol. 2003, 38, 143–158. [Google Scholar] [CrossRef]
- Katz, L. Manipulation of Modular Polyketide Synthases. Chem. Rev. 1997, 97, 2557–2576. [Google Scholar] [CrossRef]
- Huffman, J.; Gerber, R.; Du, L. Recent advancements in the biosynthetic mechanisms for polyketide-derived mycotoxins. Biopolymers 2010, 93, 764–776. [Google Scholar] [CrossRef] [PubMed]
- Hopwood, D.A. Complex enzymes in microbial natural product biosynthesis, part B: Polyketides, aminocoumarins and carbohydrates. Preface. Methods Enzymol. 2009, 459, xvii–xix. [Google Scholar] [CrossRef] [PubMed]
- Eisenman, H.C.; Greer, E.M.; McGrail, C.W. The role of melanins in melanotic fungi for pathogenesis and environmental survival. Appl. Microbiol. Biotechnol. 2020, 104, 4247–4257. [Google Scholar] [CrossRef]
- Morris-Jones, R.; Youngchim, S.; Gomez, B.L.; Aisen, P.; Hay, R.J.; Nosanchuk, J.D.; Casadevall, A.; Hamilton, A.J. Synthesis of Melanin-Like Pigments by Sporothrix schenckiix In Vitro and during Mammalian Infection. Infect. Immun. 2003, 71, 4026–4033. [Google Scholar] [CrossRef] [Green Version]
- Walker, C.A.; Gómez, B.L.; Mora-Montes, H.M.; Mackenzie, K.S.; Munro, C.A.; Brown, A.J.P.; Gow, N.A.R.; Kibbler, C.C.; Odds, F.C. Melanin externalization in Candida albicans depends on cell wall chitin structures. Eukaryot Cell 2010, 9, 1329–1342. [Google Scholar] [CrossRef] [Green Version]
- Nosanchuk, J.D.; Van Duin, D.; Mandal, P.; Aisen, P.; Legendre, A.M.; Casadevall, A. Blastomyces dermatitidis produces melanin in vitro and during infection. FEMS Microbiol. Lett. 2004, 239, 187–193. [Google Scholar] [CrossRef]
- McClelland, E.E.; Bernhardt, P.; Casadevall, A. Estimating the relative contributions of virulence factors for pathogenic microbes. Infect. Immun. 2006, 74, 1500–1504. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chatterjee, S.; Prados-Rosales, R.; Frases, S.; Itin, B.; Casadevall, A.; Stark, R.E. Using solid-state NMR to monitor the molecular consequences of Cryptococcus neoformans melanization with different catecholamine precursors. Biochemistry 2012, 51, 6080–6088. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Almeida-Paes, R.; Frases, S.; Fialho Monteiro, P.C.; Gutierrez-Galhardo, M.C.; Zancope-Oliveira, R.M.; Nosanchuk, J.D. Growth conditions influence melanization of Brazilian clinical Sporothrix schenckii isolates. Microbes Infect. 2009, 11, 554–562. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Eisenman, H.C.; Casadevall, A. Synthesis and assembly of fungal melanin. Appl. Microbiol. Biotechnol. 2012, 93, 931–940. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rosas, A.L.; Nosanchuk, J.D.; Feldmesser, M.; Cox, G.M.; McDade, H.C.; Casadevall, A. Synthesis of polymerized melanin by Cryptococcus neoformans in infected rodents. Infect. Immun. 2000, 68, 2845–2853. [Google Scholar] [CrossRef] [Green Version]
- Kwon-Chung, K.J.; Polacheck, I.; Popkin, T.J. Melanin-lacking mutants of Cryptococcus neoformans and their virulence for mice. J. Bacteriol. 1982, 150, 1414–1421. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rhodes, J.C.; Polacheck, I.; Kwon-Chung, K.J. Phenoloxidase activity and virulence in isogenic strains of Cryptococcus neoformans. Infect. Immun. 1982, 36, 1175–1184. [Google Scholar] [CrossRef] [Green Version]
- Williamson, P.R. Biochemical and molecular characterization of the diphenol oxidase of Cryptococcus neoformans: Identification as a laccase. J. Bacteriol. 1994, 176, 656–664. [Google Scholar] [CrossRef] [Green Version]
- Salas, S.D.; Bennett, J.E.; Kwon-Chung, K.J.; Perfect, J.R.; Williamson, P.R. Effect of the laccase gene CNLAC1, on virulence of Cryptococcus neoformans. J. Exp. Med. 1996, 184, 377–386. [Google Scholar] [CrossRef]
- Pukkila-Worley, R.; Gerrald, Q.D.; Kraus, P.R.; Boily, M.-J.; Davis, M.J.; Giles, S.S.; Cox, G.M.; Heitman, J.; Alspaugh, J.A. Transcriptional network of multiple capsule and melanin genes governed by the Cryptococcus neoformans cyclic AMP cascade. Eukaryot Cell 2005, 4, 190–201. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhong, J.; Frases, S.; Wang, H.; Casadevall, A.; Stark, R.E. Following fungal melanin biosynthesis with solid-state NMR: Biopolymer molecular structures and possible connections to cell-wall polysaccharides. Biochemistry 2008, 47, 4701–4710. [Google Scholar] [CrossRef]
- Eisenman, H.C.; Frases, S.; Nicola, A.M.; Rodrigues, M.L.; Casadevall, A. Vesicle-associated melanization in Cryptococcus neoformans. Microbiology 2009, 155, 3860–3867. [Google Scholar] [CrossRef] [Green Version]
- Rodrigues, M.L.; Nakayasu, E.S.; Oliveira, D.L.; Nimrichter, L.; Nosanchuk, J.D.; Almeida, I.C.; Casadevall, A. Extracellular Vesicles Produced by Cryptococcus neoformans Contain Protein Components Associated with Virulence. J. Eukaryotic. Cell 2008, 7, 58–67. [Google Scholar] [CrossRef] [Green Version]
- Panepinto, J.; Komperda, K.; Frases, S.; Park, Y.-D.; Djordjevic, J.T.; Casadevall, A.; Williamson, P.R. Sec6-dependent sorting of fungal extracellular exosomes and laccase of Cryptococcus neoformans. Mol. Microbiol. 2009, 71, 1165–1176. [Google Scholar] [CrossRef] [PubMed]
- San-Blas, G.; Guanipa, O.; Moreno, B.; Pekerar, S.; San-Blas, F. Cladosporium carrionii and Hormoconis resinae (C. resinae): Cell wall and melanin studies. Curr. Microbiol. 1996, 32, 11–16. [Google Scholar] [CrossRef] [PubMed]
- Upadhyay, S.; Xu, X.; Lowry, D.; Jackson, J.C.; Roberson, R.W.; Lin, X. Subcellular Compartmentalization and Trafficking of the Biosynthetic Machinery for Fungal Melanin. Cell Rep. 2016, 14, 2511–2518. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nosanchuk, J.D.; Stark, R.E.; Casadevall, A. Fungal Melanin: What do We Know About Structure? Front. Microbiol. 2015, 6. [Google Scholar] [CrossRef] [Green Version]
- Nosanchuk, J.D.; Casadevall, A. Budding of melanized Cryptococcus neoformans in the presence or absence of L-dopa. Microbiology 2003, 149, 1945–1951. [Google Scholar] [CrossRef] [Green Version]
- Morris-Jones, R.; Gomez, B.L.; Diez, S.; Uran, M.; Morris-Jones, S.D.; Casadevall, A.; Nosanchuk, J.D.; Hamilton, A.J. Synthesis of Melanin Pigment by Candida albicans In Vitro and during Infection. Infect. Immun. 2005, 73, 6147–6150. [Google Scholar] [CrossRef] [Green Version]
- Rosas, Á.L.; Nosanchuk, J.D.; Gómez, B.L.; Edens, W.A.; Henson, J.M.; Casadevall, A. Isolation and serological analyses of fungal melanins. J. Immunol. Methods 2000, 244, 69–80. [Google Scholar] [CrossRef]
- Gómez, B.L.; Nosanchuk, J.D.; Díez, S.; Youngchim, S.; Aisen, P.; Cano, L.E.; Restrepo, A.; Casadevall, A.; Hamilton, A.J. Detection of Melanin-Like Pigments in the Dimorphic Fungal Pathogen Paracoccidioides brasiliensis In Vitro and during Infection. J. Infect. Immun. 2001, 69, 5760–5767. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nosanchuk, J.D.; Yu, J.-J.; Hung, C.-Y.; Casadevall, A.; Cole, G.T. Coccidioides posadasii produces melanin in vitro and during infection. Fungal Genet. Biol. 2007, 44, 517–520. [Google Scholar] [CrossRef] [PubMed]
- Nosanchuk, J.D.; Gómez, B.L.; Youngchim, S.; Díez, S.; Aisen, P.; Zancopé-Oliveira, R.M.; Restrepo, A.; Casadevall, A.; Hamilton, A.J. Histoplasma capsulatum synthesizes melanin-like pigments in vitro and during mammalian infection. Infect. Immun. 2002, 70, 5124–5131. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Eisenman, H.C.; Nosanchuk, J.D.; Webber, J.B.; Emerson, R.J.; Camesano, T.A.; Casadevall, A. Microstructure of cell wall-associated melanin in the human pathogenic fungus Cryptococcus neoformans. Biochemistry 2005, 44, 3683–3693. [Google Scholar] [CrossRef] [PubMed]
- Pihet, M.; Vandeputte, P.; Tronchin, G.; Renier, G.; Saulnier, P.; Georgeault, S.; Mallet, R.; Chabasse, D.; Symoens, F.; Bouchara, J.P. Melanin is an essential component for the integrity of the cell wall of Aspergillus fumigatus conidia. BMC Microbiol. 2009, 9, 177. [Google Scholar] [CrossRef]
- Jacobson, E.S.; Ikeda, R. Effect of melanization upon porosity of the cryptococcal cell wall. Med. Mycol. 2005, 43, 327–333. [Google Scholar] [CrossRef] [Green Version]
- Franzen, A.J.; Cunha, M.M.; Batista, E.J.; Seabra, S.H.; De Souza, W.; Rozental, S. Effects of tricyclazole (5-methyl-1,2,4-triazol [3,4] benzothiazole), a specific DHN-melanin inhibitor, on the morphology of Fonsecaea pedrosoi conidia and sclerotic cells. Microsc. Res. Tech. 2006, 69, 729–737. [Google Scholar] [CrossRef] [PubMed]
- Hegnauer, H.; Nyhle´n, L.E.; Rast, D.M. Ultrastructure of native and synthetic Agaricus bisporus melanins—Implications as to the compartmentation of melanogenesis in fungi. Exp. Mycol. 1985, 9, 1–29. [Google Scholar] [CrossRef]
- Kogej, T.; Stein, M.; Volkmann, M.; Gorbushina, A.A.; Galinski, E.A.; Gunde-Cimerman, N. Osmotic adaptation of the halophilic fungus Hortaea werneckii: Role of osmolytes and melanization. Microbiology 2007, 153, 4261–4273. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Munro, C.A.; Gow, N.A. Chitin synthesis in human pathogenic fungi. Med. Mycol. 2001, 39 (Suppl. 1), 41–53. [Google Scholar] [CrossRef]
- Baker, L.G.; Specht, C.A.; Donlin, M.J.; Lodge, J.K. Chitosan, the deacetylated form of chitin, is necessary for cell wall integrity in Cryptococcus neoformans. Eukaryot Cell 2007, 6, 855–867. [Google Scholar] [CrossRef] [Green Version]
- Banks, I.R.; Specht, C.A.; Donlin, M.J.; Gerik, K.J.; Levitz, S.M.; Lodge, J.K. A Chitin Synthase and Its Regulator Protein Are Critical for Chitosan Production and Growth of the Fungal Pathogen Cryptococcus neoformans. J. Eukaryot. Cell 2005, 4, 1902–1912. [Google Scholar] [CrossRef] [Green Version]
- Walton, F.J.; Idnurm, A.; Heitman, J. Novel gene functions required for melanization of the human pathogen Cryptococcus neoformans. Mol. Microbiol. 2005, 57, 1381–1396. [Google Scholar] [CrossRef] [PubMed]
- Upadhya, R.; Lam, W.C.; Maybruck, B.; Specht, C.A.; Levitz, S.M.; Lodge, J.K. Induction of Protective Immunity to Cryptococcal Infection in Mice by a Heat-Killed, Chitosan-Deficient Strain of Cryptococcus neoformans. mBio 2016, 7. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nosanchuk, J.D.; Rudolph, J.; Rosas, A.L.; Casadevall, A. Evidence That Cryptococcus neoformans Is Melanized in Pigeon Excreta: Implications for Pathogenesis. Infect. Immun. 1999, 67, 5477–5479. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rosas, Á.L.; Casadevall, A. Melanization affects susceptibility of Cryptococcus neoformans to heat and cold. FEMS Microbiol. Lett. 1997, 153, 265–272. [Google Scholar] [CrossRef]
- Zhdanova, N.; Gavriushina, A.; Vasilevskaia, A. Effect of gamma and UV irradiation on the survival of Cladosporium sp. and Oidiodendron cerealis. Mikrobiolohichnyi Zhurnal 1973, 35, 449–452. [Google Scholar] [PubMed]
- Mironenko, N.V.; Alekhina, I.A.; Zhdanova, N.N.; Bulat, S.A. Intraspecific variation in gamma-radiation resistance and genomic structure in the filamentous fungus Alternaria alternata: A case study of strains inhabiting Chernobyl reactor no. 4. Ecotoxicol. Environ. Saf. 2000, 45, 177–187. [Google Scholar] [CrossRef]
- Nosanchuk, J.D.; Casadevall, A. Impact of melanin on microbial virulence and clinical resistance to antimicrobial compounds. Antimicrob. Agents Chemother. 2006, 50, 3519–3528. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dadachova, E.; Bryan, R.A.; Huang, X.; Moadel, T.; Schweitzer, A.D.; Aisen, P.; Nosanchuk, J.D.; Casadevall, A. Ionizing radiation changes the electronic properties of melanin and enhances the growth of melanized fungi. PLoS ONE 2007, 2, e457. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Robertson, K.L.; Mostaghim, A.; Cuomo, C.A.; Soto, C.M.; Lebedev, N.; Bailey, R.F.; Wang, Z. Adaptation of the black yeast Wangiella dermatitidis to ionizing radiation: Molecular and cellular mechanisms. PLoS ONE 2012, 7, e48674. [Google Scholar] [CrossRef] [Green Version]
- Cordero, R.J.; Casadevall, A. Functions of fungal melanin beyond virulence. Fungal Biol. Rev. 2017, 31, 99–112. [Google Scholar] [CrossRef]
- Rosas, A.L.; Casadevall, A. Melanization decreases the susceptibility of Cryptococcus neoformans to enzymatic degradation. Mycopathologia 2001, 151, 53–56. [Google Scholar] [CrossRef] [PubMed]
- Fogarty, R.V.; Tobin, J.M. Fungal melanins and their interactions with metals. Enzym. Microb. Technol. 1996, 19, 311–317. [Google Scholar] [CrossRef]
- Garcia-Rivera, J.; Casadevall, A. Melanization of Cryptococcus neoformans reduces its susceptibility to the antimicrobial effects of silver nitrate. Sabouraudia 2001, 39, 353–357. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ikeda, R.; Sugita, T.; Jacobson, E.S.; Shinoda, T. Effects of melanin upon susceptibility of Cryptococcus to antifungals. Microbiol. Immunol. 2003, 47, 271–277. [Google Scholar] [CrossRef]
- Barza, M.; Baum, J.; Kane, A. Inhibition of antibiotic activity in vitro by synthetic melanin. Antimicrob. Agents Chemother. 1976, 10, 569. [Google Scholar] [CrossRef] [Green Version]
- Fukuda, M.; Sasaki, K. Changes in the antibacterial activity of melanin-bound drugs. Ophthalmic Res. 1990, 22, 123–127. [Google Scholar] [CrossRef]
- White, L.P. Melanin: A Naturally Occurring Cation Exchange Material. Nature 1958, 182, 1427–1428. [Google Scholar] [CrossRef] [PubMed]
- Nosanchuk, J.D.; Casadevall, A. Cellular charge of Cryptococcus neoformans: Contributions from the capsular polysaccharide, melanin, and monoclonal antibody binding. Infect. Immun. 1997, 65, 1836–1841. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kozel, T.R. Dissociation of a hydrophobic surface from phagocytosis of encapsulated and non-encapsulated Cryptococcus neoformans. Infect. Immun. 1983, 39, 1214–1219. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Walter, H.; Graham, L.L.; Krob, E.J.; Hill, M. Correlation between phagocytic and membrane surface properties reflected by partitioning of human peripheral blood monocytes in two-polymer aqueous phases. Biochim. Biophys. Acta. 1980, 602, 309–322. [Google Scholar] [CrossRef]
- Eggleton, P.; Crawford, N.; Fisher, D. Fractionation of human neutrophils into subpopulations by countercurrent distribution: Surface charge and functional heterogeneity. Eur. J. Cell Biol. 1992, 57, 265–272. [Google Scholar]
- Mednick, A.J.; Nosanchuk, J.D.; Casadevall, A. Melanization of Cryptococcus neoformans affects lung inflammatory responses during cryptococcal infection. Infect. Immun. 2005, 73, 2012–2019. [Google Scholar] [CrossRef] [Green Version]
- Wang, Y.; Aisen, P.; Casadevall, A. Cryptococcus neoformans melanin and virulence: Mechanism of action. Infect. Immun. 1995, 63, 3131–3136. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Doering, T.L.; Nosanchuk, J.D.; Roberts, W.K.; Casadevall, A. Melanin as a potential cryptococcal defence against microbicidal proteins. Med. Mycol. 1999, 37, 175–181. [Google Scholar] [CrossRef]
- Alvarez, M.; Casadevall, A. Phagosome extrusion and host-cell survival after Cryptococcus neoformans phagocytosis by macrophages. Curr. Biol. 2006, 16, 2161–2165. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Frazão, S.d.O.; Sousa, H.R.d.; Silva, L.G.d.; Folha, J.d.S.; Gorgonha, K.C.d.M.; Oliveira, G.P.d.; Felipe, M.S.S.; Silva-Pereira, I.; Casadevall, A.; Nicola, A.M.; et al. Laccase Affects the Rate of Cryptococcus neoformans Nonlytic Exocytosis from Macrophages. mBio 2020, 11, e02085-20. [Google Scholar] [CrossRef] [PubMed]
- Nosanchuk, J.D.; Casadevall, A. The contribution of melanin to microbial pathogenesis. Cell Microbiol. 2003, 5, 203–223. [Google Scholar] [CrossRef] [PubMed]
- Sabiiti, W.; Robertson, E.; Beale, M.A.; Johnston, S.A.; Brouwer, A.E.; Loyse, A.; Jarvis, J.N.; Gilbert, A.S.; Fisher, M.C.; Harrison, T.S.; et al. Efficient phagocytosis and laccase activity affect the outcome of HIV-associated cryptococcosis. J. Clin. Investig. 2014, 124, 2000–2008. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rosas, A.L.; MacGill, R.S.; Nosanchuk, J.D.; Kozel, T.R.; Casadevall, A. Activation of the alternative complement pathway by fungal melanins. Clin. Diagn Lab. Immunol. 2002, 9, 144–148. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Goldman, D.L.; Lee, S.C.; Mednick, A.J.; Montella, L.; Casadevall, A. Persistent Cryptococcus neoformans pulmonary infection in the rat is associated with intracellular parasitism, decreased inducible nitric oxide synthase expression, and altered antibody responsiveness to cryptococcal polysaccharide. Infect. Immun. 2000, 68, 832–838. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liu, L.; Tewari, R.P.; Williamson, P.R. Laccase Protects Cryptococcus neoformans from Antifungal Activity of Alveolar Macrophages. Infect. Immun. 1999, 67, 6034–6039. [Google Scholar] [CrossRef] [Green Version]
- Minton, K. Granuloma macrophage differentiation. Nat. Rev. Immunol. 2016, 16, 719. [Google Scholar] [CrossRef]
- Tajima, K.; Yamanaka, D.; Ishibashi, K.I.; Adachi, Y.; Ohno, N. Solubilized melanin suppresses macrophage function. FEBS Open Bio 2019, 9, 791–800. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mitchell, T.G.; Friedman, L. In vitro phagocytosis and intracellular fate of variously encapsulated strains of Cryptococcus neoformans. Infect. Immun. 1972, 5, 491–498. [Google Scholar] [CrossRef] [Green Version]
- Huffnagle, G.B.; Chen, G.H.; Curtis, J.L.; McDonald, R.A.; Strieter, R.M.; Toews, G.B. Down-regulation of the afferent phase of T cell-mediated pulmonary inflammation and immunity by a high melanin-producing strain of Cryptococcus neoformans. J. Immunol. 1995, 155, 3507–3516. [Google Scholar] [PubMed]
- Barluzzi, R.; Brozzetti, A.; Mariucci, G.; Tantucci, M.; Neglia, R.G.; Bistoni, F.; Blasi, E. Establishment of protective immunity against cerebral cryptococcosis by means of an avirulent, non melanogenic Cryptococcus neoformans strain. J. Neuroimmunol. 2000, 109, 75–86. [Google Scholar] [CrossRef]
- Nosanchuk, J.D.; Rosas, A.L.; Casadevall, A. The antibody response to fungal melanin in mice. J. Immunol. 1998, 160, 6026–6031. [Google Scholar]
- Nosanchuk, J.D.; Valadon, P.; Feldmesser, M.; Casadevall, A. Melanization of Cryptococcus neoformans in murine infection. Mol. Cell Biol. 1999, 19, 745–750. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rosas, A.L.; Nosanchuk, J.D.; Casadevall, A. Passive immunization with melanin-binding monoclonal antibodies prolongs survival of mice with lethal Cryptococcus neoformans infection. Infect. Immun. 2001, 69, 3410–3412. [Google Scholar] [CrossRef] [Green Version]
- Kozel, T.R.; Highison, B.; Stratton, C.J. Localization on encapsulated Cryptococcus neoformans of serum components opsonic for phagocytosis by macrophages and neutrophils. Infect. Immun. 1984, 43, 574–579. [Google Scholar] [CrossRef] [Green Version]
- Davies, S.F.; Clifford, D.P.; Hoidal, J.R.; Repine, J.E. Opsonic requirements for the uptake of Cryptococcus neoformans by human polymorphonuclear leukocytes and monocytes. J. Infect. Dis. 1982, 145, 870–874. [Google Scholar] [CrossRef]
- Tsai, H.F.; Chang, Y.C.; Washburn, R.G.; Wheeler, M.H.; Kwon-Chung, K.J. The developmentally regulated alb1 gene of Aspergillus fumigatus: Its role in modulation of conidial morphology and virulence. J. Bacteriol. 1998, 180, 3031–3038. [Google Scholar] [CrossRef] [Green Version]
- Tsai, H.F.; Wheeler, M.H.; Chang, Y.C.; Kwon-Chung, K.J. A developmentally regulated gene cluster involved in conidial pigment biosynthesis in Aspergillus fumigatus. J. Bacteriol. 1999, 181, 6469–6477. [Google Scholar] [CrossRef] [Green Version]
- Abad, A.; Fernández-Molina, J.V.; Bikandi, J.; Ramírez, A.; Margareto, J.; Sendino, J.; Hernando, F.L.; Pontón, J.; Garaizar, J.; Rementeria, A. What makes Aspergillus fumigatus a successful pathogen? Genes and molecules involved in invasive aspergillosis. Rev. Iberoam. Micol. 2010, 27, 155–182. [Google Scholar] [CrossRef]
- Jahn, B.; Boukhallouk, F.; Lotz, J.; Langfelder, K.; Wanner, G.; Brakhage, A.A. Interaction of human phagocytes with pigmentless Aspergillus conidia. Infect. Immun. 2000, 68, 3736–3739. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chai, L.Y.; Netea, M.G.; Sugui, J.; Vonk, A.G.; Van de Sande, W.W.; Warris, A.; Kwon-Chung, K.J.; Kullberg, B.J. Aspergillus fumigatus conidial melanin modulates host cytokine response. Immunobiology 2010, 215, 915–920. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Luther, K.; Torosantucci, A.; Brakhage, A.A.; Heesemann, J.; Ebel, F. Phagocytosis of Aspergillus fumigatus conidia by murine macrophages involves recognition by the dectin-1 beta-glucan receptor and Toll-like receptor 2. Cell. Microbiol. 2007, 9, 368–381. [Google Scholar] [CrossRef]
- Jahn, B.; Langfelder, K.; Schneider, U.; Schindel, C.; Brakhage, A.A. PKSP-dependent reduction of phagolysosome fusion and intracellular kill of Aspergillus fumigatus conidia by human monocyte-derived macrophages. Cell. Microbiol. 2002, 4, 793–803. [Google Scholar] [CrossRef]
- Thywißen, A.; Heinekamp, T.; Dahse, H.M.; Schmaler-Ripcke, J.; Nietzsche, S.; Zipfel, P.F.; Brakhage, A.A. Conidial Dihydroxynaphthalene Melanin of the Human Pathogenic Fungus Aspergillus fumigatus Interferes with the Host Endocytosis Pathway. Front. Microbiol. 2011, 2, 96. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sanjuan, M.A.; Dillon, C.P.; Tait, S.W.; Moshiach, S.; Dorsey, F.; Connell, S.; Komatsu, M.; Tanaka, K.; Cleveland, J.L.; Withoff, S.; et al. Toll-like receptor signalling in macrophages links the autophagy pathway to phagocytosis. Nature 2007, 450, 1253–1257. [Google Scholar] [CrossRef]
- Kyrmizi, I.; Ferreira, H.; Carvalho, A.; Figueroa, J.A.L.; Zarmpas, P.; Cunha, C.; Akoumianaki, T.; Stylianou, K.; Deepe, G.S.; Samonis, G.; et al. Calcium sequestration by fungal melanin inhibits calcium–calmodulin signalling to prevent LC3-associated phagocytosis. Nat. Microbiol. 2018, 3, 791–803. [Google Scholar] [CrossRef] [PubMed]
- Martinez, J.; Malireddi, R.S.; Lu, Q.; Cunha, L.D.; Pelletier, S.; Gingras, S.; Orchard, R.; Guan, J.-L.; Tan, H.; Peng, J. Molecular characterization of LC3-associated phagocytosis reveals distinct roles for Rubicon, NOX2 and autophagy proteins. Nat. Cell Biol. 2015, 17, 893–906. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yang, C.-S.; Lee, J.-S.; Rodgers, M.; Min, C.-K.; Lee, J.-Y.; Kim, H.J.; Lee, K.-H.; Kim, C.-J.; Oh, B.; Zandi, E. Autophagy protein Rubicon mediates phagocytic NADPH oxidase activation in response to microbial infection or TLR stimulation. Cell Host Microbe 2012, 11, 264–276. [Google Scholar] [CrossRef] [Green Version]
- Akoumianaki, T.; Kyrmizi, I.; Valsecchi, I.; Gresnigt, M.S.; Samonis, G.; Drakos, E.; Boumpas, D.; Muszkieta, L.; Prevost, M.-C.; Kontoyiannis, D.P. Aspergillus cell wall melanin blocks LC3-associated phagocytosis to promote pathogenicity. Cell Host Microbe 2016, 19, 79–90. [Google Scholar] [CrossRef] [Green Version]
- Bayry, J.; Beaussart, A.; Dufrêne, Y.F.; Sharma, M.; Bansal, K.; Kniemeyer, O.; Aimanianda, V.; Brakhage, A.A.; Kaveri, S.V.; Kwon-Chung, K.J.; et al. Surface structure characterization of Aspergillus fumigatus conidia mutated in the melanin synthesis pathway and their human cellular immune response. Infect. Immun. 2014, 82, 3141–3153. [Google Scholar] [CrossRef] [Green Version]
- Stappers, M.H.T.; Clark, A.E.; Aimanianda, V.; Bidula, S.; Reid, D.M.; Asamaphan, P.; Hardison, S.E.; Dambuza, I.M.; Valsecchi, I.; Kerscher, B.; et al. Recognition of DHN-melanin by a C-type lectin receptor is required for immunity to Aspergillus. Nature 2018, 555, 382–386. [Google Scholar] [CrossRef]
- Gonçalves, S.M.; Duarte-Oliveira, C.; Campos, C.F.; Aimanianda, V.; Ter Horst, R.; Leite, L.; Mercier, T.; Pereira, P.; Fernández-García, M.; Antunes, D.; et al. Phagosomal removal of fungal melanin reprograms macrophage metabolism to promote antifungal immunity. Nat. Commun. 2020, 11, 2282. [Google Scholar] [CrossRef] [PubMed]
- Majumder, P.K.; Febbo, P.G.; Bikoff, R.; Berger, R.; Xue, Q.; McMahon, L.M.; Manola, J.; Brugarolas, J.; McDonnell, T.J.; Golub, T.R.; et al. mTOR inhibition reverses Akt-dependent prostate intraepithelial neoplasia through regulation of apoptotic and HIF-1-dependent pathways. Nat. Med. 2004, 10, 594–601. [Google Scholar] [CrossRef]
- Sturtevant, J.; Latgé, J.P. Participation of complement in the phagocytosis of the conidia of Aspergillus fumigatus by human polymorphonuclear cells. J. Infect. Dis. 1992, 166, 580–586. [Google Scholar] [CrossRef] [PubMed]
- Behnsen, J.; Hartmann, A.; Schmaler, J.; Gehrke, A.; Brakhage, A.A.; Zipfel, P.F. The Opportunistic Human Pathogenic Fungus Aspergillus fumigatus Evades the Host Complement System. J. Infect. Immun. 2008, 76, 820–827. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tsai, H.F.; Washburn, R.G.; Chang, Y.C.; Kwon-Chung, K.J. Aspergillus fumigatus arp1 modulates conidial pigmentation and complement deposition. Mol. Microbiol. 1997, 26, 175–183. [Google Scholar] [CrossRef]
- Langfelder, K.; Jahn, B.; Gehringer, H.; Schmidt, A.; Wanner, G.; Brakhage, A.A. Identification of a polyketide synthase gene (pksP) of Aspergillus fumigatus involved in conidial pigment biosynthesis and virulence. Med. Microbiol. Immunol. 1998, 187, 79–89. [Google Scholar] [CrossRef]
- Zhang, J.; Wang, L.; Xi, L.; Huang, H.; Hu, Y.; Li, X.; Huang, X.; Lu, S.; Sun, J. Melanin in a meristematic mutant of Fonsecaea monophora inhibits the production of nitric oxide and Th1 cytokines of murine macrophages. Mycopathologia 2013, 175, 515–522. [Google Scholar] [CrossRef] [PubMed]
- Shi, M.; Sun, J.; Lu, S.; Qin, J.; Xi, L.; Zhang, J. Transcriptional profiling of macrophages infected with Fonsecaea monophora. Mycoses 2019, 62, 374–383. [Google Scholar] [CrossRef]
- Pinto, L.; Granja, L.F.Z.; Almeida, M.A.d.; Alviano, D.S.; Silva, M.H.d.; Ejzemberg, R.; Rozental, S.; Alviano, C.S. Melanin particles isolated from the fungus Fonsecaea pedrosoi activates the human complement system. Memórias do Instituto Oswaldo Cruz 2018, 113. [Google Scholar] [CrossRef] [PubMed]
- Youngchim, S.; Hay, R.J.; Hamilton, A.J. Melanization of Penicillium marneffei in vitro and in vivo. Microbiology 2005, 151, 291–299. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Uran, M.E.; Nosanchuk, J.D.; Restrepo, A.; Hamilton, A.J.; Gomez, B.L.; Cano, L.E. Detection of antibodies against Paracoccidioides brasiliensis melanin in in vitro and in vivo studies during infection. Clin. Vaccine Immunol. 2011, 18, 1680–1688. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Emidio, E.C.P.; Uran, M.E.; Silva, L.B.R.; Dias, L.S.; Doprado, M.; Nosanchuk, J.D.; Taborda, C.P. Melanin as a Virulence Factor in Different Species of Genus Paracoccidioides. J. Fungi. 2020, 6, 291. [Google Scholar] [CrossRef]
- Silva, M.B.; Thomaz, L.; Marques, A.F.; Svidzinski, A.E.; Nosanchuk, J.D.; Casadevall, A.; Travassos, L.R.; Taborda, C.P. Resistance of melanized yeast cells of Paracoccidioides brasiliensis to antimicrobial oxidants and inhibition of phagocytosis using carbohydrates and monoclonal antibody to CD18. Memórias do Instituto Oswaldo Cruz 2009, 104, 644–648. [Google Scholar] [CrossRef] [Green Version]
- Almeida-Paes, R.; Almeida, M.A.; Baeza, L.C.; Marmello, L.A.M.; Trugilho, M.R.O.; Nosanchuk, J.D.; Soares, C.M.A.; Valente, R.H.; Zancopé-Oliveira, R.M. Beyond Melanin: Proteomics Reveals Virulence-Related Proteins in Paracoccidioides brasiliensis and Paracoccidioides lutzii Yeast Cells Grown in the Presence of L-Dihydroxyphenylalanine. J. Fungi. 2020, 6, 328. [Google Scholar] [CrossRef]
- Tam, E.W.; Tsang, C.C.; Lau, S.K.; Woo, P.C. Polyketides, toxins and pigments in Penicillium marneffei. Toxins 2015, 7, 4421–4436. [Google Scholar] [CrossRef] [Green Version]
- Sapmak, A.; Kaewmalakul, J.; Nosanchuk, J.D.; Vanittanakom, N.; Andrianopoulos, A.; Pruksaphon, K.; Youngchim, S. Talaromyces marneffei laccase modifies THP-1 macrophage responses. Virulence 2016, 7, 702–717. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kaewmalakul, J.; Nosanchuk, J.D.; Vanittanakom, N.; Youngchim, S. Melanization and morphological effects on antifungal susceptibility of Penicillium marneffei. Antonie Van Leeuwenhoek 2014, 106, 1011–1020. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Boyce, K.J.; McLauchlan, A.; Schreider, L.; Andrianopoulos, A. Intracellular growth is dependent on tyrosine catabolism in the dimorphic fungal pathogen Penicillium marneffei. PLoS Pathog. 2015, 11, e1004790. [Google Scholar] [CrossRef] [Green Version]
- Almeida-Paes, R.; Frases, S.; Araújo Gde, S.; De Oliveira, M.M.; Gerfen, G.J.; Nosanchuk, J.D.; Zancopé-Oliveira, R.M. Biosynthesis and functions of a melanoid pigment produced by species of the Sporothrix complex in the presence of L-tyrosine. Appl. Environ. Microbiol. 2012, 78, 8623–8630. [Google Scholar] [CrossRef] [Green Version]
- Cruz, I.L.R.; Figueiredo-Carvalho, M.H.G.; Zancopé-Oliveira, R.M.; Almeida-Paes, R. Evaluation of melanin production by Sporothrix luriei. Memórias do Instituto Oswaldo Cruz 2018, 113, 68–70. [Google Scholar] [CrossRef] [Green Version]
- Song, Y.; Yao, L.; Zhen, Y.; Cui, Y.; Zhong, S.; Liu, Y.; Li, S. Sporothrix globosa melanin inhibits antigen presentation by macrophages and enhances deep organ dissemination. Braz J. Microbiol. 2020. [Google Scholar] [CrossRef]
- Almeida-Paes, R.; de Oliveira, L.C.; Oliveira, M.M.; Gutierrez-Galhardo, M.C.; Nosanchuk, J.D.; Zancope-Oliveira, R.M. Phenotypic characteristics associated with virulence of clinical isolates from the Sporothrix complex. BioMed Res. Int. 2015, 2015, 212308. [Google Scholar] [CrossRef] [Green Version]
- Madrid, I.M.; Xavier, M.O.; Mattei, A.S.; Fernandes, C.G.; Guim, T.N.; Santin, R.; Schuch, L.F.; Nobre Mde, O.; Araújo Meireles, M.C. Role of melanin in the pathogenesis of cutaneous sporotrichosis. Microbes Infect. 2010, 12, 162–165. [Google Scholar] [CrossRef]
- Mario, D.A.; Santos, R.C.; Denardi, L.B.; Vaucher Rde, A.; Santurio, J.M.; Alves, S.H. Interference of melanin in the susceptibility profile of Sporothrix species to amphotericin B. Rev. Iberoam. Micol. 2016, 33, 21–25. [Google Scholar] [CrossRef]
- Almeida-Paes, R.; Figueiredo-Carvalho, M.H.; Brito-Santos, F.; Almeida-Silva, F.; Oliveira, M.M.; Zancopé-Oliveira, R.M. Melanins Protect Sporothrix brasiliensis and Sporothrix schenckii from the Antifungal Effects of Terbinafine. PLoS ONE 2016, 11, e0152796. [Google Scholar] [CrossRef]
- Soliman, S.S.M.; Hamdy, R.; Elseginy, S.A.; Gebremariam, T.; Hamoda, A.M.; Madkour, M.; Venkatachalam, T.; Ershaid, M.N.; Mohammad, M.G.; Chamilos, G.; et al. Selective inhibition of Rhizopus eumelanin biosynthesis by novel natural product scaffold-based designs caused significant inhibition of fungal pathogenesis. Biochem. J. 2020, 477, 2489–2507. [Google Scholar] [CrossRef]
- Chatterjee, S.; Prados-Rosales, R.; Tan, S.; Phan, V.C.; Chrissian, C.; Itin, B.; Wang, H.; Khajo, A.; Magliozzo, R.S.; Casadevall, A.; et al. The melanization road more traveled by: Precursor substrate effects on melanin synthesis in cell-free and fungal cell systems. J. Biol. Chem. 2018, 293, 20157–20168. [Google Scholar] [CrossRef] [Green Version]
- Cordero, R.J.B.; Casadevall, A. Melanin. Curr. Biol. 2020, 30, R142–R143. [Google Scholar] [CrossRef]
- Camacho, E.; Vij, R.; Chrissian, C.; Prados-Rosales, R.; Gil, D.; O’Meally, R.N.; Cordero, R.J.B.; Cole, R.N.; McCaffery, J.M.; Stark, R.E.; et al. The structural unit of melanin in the cell wall of the fungal pathogen Cryptococcus neoformans. J. Biol. Chem. 2019, 294, 10471–10489. [Google Scholar] [CrossRef] [PubMed]
- Galeb, H.A.; Wilkinson, E.L.; Stowell, A.F.; Lin, H.; Murphy, S.T.; Martin-Hirsch, P.L.; Mort, R.L.; Taylor, A.M.; Hardy, J.G. Melanins as Sustainable Resources for Advanced Biotechnological Applications. Glob. Chall. 2020, 5, 2000102. [Google Scholar] [CrossRef] [PubMed]
- Cordero, R.J.B. Melanin for space travel radioprotection. Environ. Microbiol. 2017, 19, 2529–2532. [Google Scholar] [CrossRef] [PubMed]
Figure 1.
Current knowledge of melanin synthetic pathways in fungi.

Figure 2.
Cryptococcus neoformans melanin “ghosts” obtained from lungs of infected mice as described in [20]. The red arrows show bud scars on the melanin.
Figure 2.
Cryptococcus neoformans melanin “ghosts” obtained from lungs of infected mice as described in [20]. The red arrows show bud scars on the melanin.


{kind=link}
{kind=link}
| Species | Isolate Environment | Melanin Types |
|---|---|---|
| Aspergillus fumigatus | Clinical | DHN and pyo-melanin |
| Aspergillus niger | Industrial fermentation | DHN and L-DOPA |
| Blastomyces dermatitidis | Clinical | DHN |
| Candida Albicans | Clinical | L-DOPA |
| Cryptococcus neoformans | Clinical | L-DOPA |
| Histoplasma capsulatum | Clinical | DHN and L-DOPA |
| Paracoccidioides brasiliensis | Clinical | DHN and L-DOPA |
| Fonsecaea monophora | Clinical | DHN and L-DOPA |
| Fonsecaea pedrosoi | Clinical | DHN |
| Sporothrix schenckii | Clinical | DHN |
Abbreviations: DHN is 1,8-dihydroxynaphthalene and L-DOPA is l-3,4-dihyroxyphenylalanine.
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Liu, S.; Youngchim, S.; Zamith-Miranda, D.; Nosanchuk, J.D. Fungal Melanin and the Mammalian Immune System. J. Fungi 2021, 7, 264. https://0-doi-org.brum.beds.ac.uk/10.3390/jof7040264
AMA Style
Liu S, Youngchim S, Zamith-Miranda D, Nosanchuk JD. Fungal Melanin and the Mammalian Immune System. Journal of Fungi. 2021; 7(4):264. https://0-doi-org.brum.beds.ac.uk/10.3390/jof7040264
Chicago/Turabian StyleLiu, Sichen, Sirida Youngchim, Daniel Zamith-Miranda, and Joshua D. Nosanchuk. 2021. "Fungal Melanin and the Mammalian Immune System" Journal of Fungi 7, no. 4: 264. https://0-doi-org.brum.beds.ac.uk/10.3390/jof7040264
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.