
Soil Metabarcoding Offers a New Tool for the Investigation and Hunting of Truffles in Northern Thailand



1
Research Center of Microbial Diversity and Sustainable Utilization, Chiang Mai University, Chiang Mai 50200, Thailand
2
Department of Biology, Faculty of Science, Chiang Mai University, Chiang Mai 50200, Thailand
3
Bioinformatic and Systems Biology Program, King Mongkut’s University of Technology Thonburi, Bang Kun Thian, Bangkok 10150, Thailand
4
Academy of Science, The Royal Society of Thailand, Bangkok 10300, Thailand
*
Author to whom correspondence should be addressed.
J. Fungi 2021, 7(4), 293; https://0-doi-org.brum.beds.ac.uk/10.3390/jof7040293
Submission received: 24 March 2021
/
Revised: 9 April 2021
/
Accepted: 12 April 2021
/
Published: 13 April 2021
(This article belongs to the Special Issue Diversity and Classification of Environmental Fungi)
Abstract
:Truffles (Tuber spp.) are well-known as edible ectomycorrhizal mushrooms, and some species are one of the most expensive foods in the world. During the fruiting process, truffles produce hypogeous ascocarps; a trained pig or dog is needed to locate the ascocarps under the ground. Truffles in northern Thailand have been recorded in association with Betula alnoides and Carpinus poilanei. In this study, we investigated the soil mycobiota diversity of soil samples from both of these truffle host plants in native forests using environmental DNA metabarcoding to target the internal transcribed spacer 1 (ITS1) region of the rDNA gene for the purposes of investigation of truffle diversity and locating truffles during the non-fruiting phase. In this study, a total of 38 soil samples were collected from different locations. Of these, truffles had been found at three of these locations. Subsequently, a total of 1341 putative taxonomic units (OTUs) were obtained. The overall fungal community was dominated by phylum-level sequences assigned to Ascomycota (57.63%), Basidiomycota (37.26%), Blastocladiomycota (0.007%), Chytridiomycota (0.21%), Glomeromycota (0.01%), Kickxellomycota (0.01%), Mortierellomycota (2.08%), Mucoromycota (0.24%), Rozellomycota (0.01%), Zoopagomycota (0.003%), and unidentified (2.54%). The results revealed that six OTUs were determined to be representative and belonged to the genus Tuber. OTU162, OTU187, OTU447, and OTU530 belonged to T. thailandicum, T. lannaense, T. bomiense, and T. magnatum, whereas OTU105 and OTU720 were acknowledged as unrecognized Tuber species. From 38 locations, OTUs of truffles were found in 33 locations (including three previously known truffle locations). Thus, 30 collection sites were considered new locations for T. thailandicum, T. bomiense, and other unrecognized Tuber species. Interestingly, at 16 new locations, mature ascocarps of truffles that were undergoing the fruiting phase were located underground. All 16 truffle samples were identified as T. thailandicum based on morphological characteristics and molecular phylogenetic analysis. However, ascocarps of other truffle species were not found at the new OTUs representative locations. The knowledge gained from this study can be used to lead researchers to a better understanding of the occurrence of truffles using soil mycobiota diversity investigation. The outcomes of this study will be particularly beneficial with respect to the search and hunt for truffles without the need for trained animals. In addition, the findings of this study will be useful for the management and conservation of truffle habitats in northern Thailand.
1. Introduction
Truffles are one of the most expensive edible mushrooms in the world and belong to the genus Tuber within the family Tuberaceae, order Pezizales [1,2]. Generally, the life cycle of a truffle takes place underground by ectomycorrhizal symbiosis, where they produce hypogeous ascocarps [2,3,4]. Ascocarps of truffles are only formed during the fruiting phase. The mature state of ascocarps of truffles leads to the release of volatile compounds that are commonly detected by insects and mammals who consume these ascocarps. This process ultimately results in the dispersal of the truffle’s spores throughout the environment [5,6]. However, it can be difficult for humans to find truffles in native forests. Traditionally, the process of searching for and collecting truffles has required the use of female pigs and trained dogs. With their advanced sense of smell, these animals are able to detect the distinct odor of mature truffles underneath the surface of the soil. Therefore, the involvement of trained animals has always been considered a crucial element in the search for truffles in their natural habitat [2,4,6]. Truffles mostly grow in forests in temperate zones throughout Asia, Europe, North Africa, and North America. Generally, woody trees in five families, namely Betulaceae, Cistaceae, Fagaceae, and Pinaceae, are known to form ectomycorrhizal relationships with most truffle species [2,4,7]. The most popular truffles in the world are the Italian white truffle (T. magnatum), the garlic truffle (T. macrosporum), the Périgord black truffle (T. melanosporum), the summer truffle (T. aestivum), and the Bianchetto white truffle (T. borchii). All of these truffle species are predominantly found in Europe [2,4,8,9]. In America, the Oregon spring white truffle (T. gibbosum) and the Oregon winter white truffle (T. oregonense) are commercially harvested in the northern region of the state [10], while the pecan truffle (T. lyonii) is harvested in the southern region [11]. Additionally, the Chinese black truffle (T. indicum) is the most famous commercial truffle in Asia [12]. Interestingly, Asian truffles have been harvested for many decades, and more than 25 new Tuber species have been discovered in Asia since the 1980s, especially in China, Japan, and Taiwan [13,14,15,16,17,18,19,20]. Previous studies have reported that there is a high degree of diversity of Tuber in Asia but this has still not been fully investigated [17,20]. Thailand is proving to be a hotspot for the discovery of a range of novel fungal species [21]. Evidence of this would include the discovery of two new Tuber species (T. lannaense and T. thailandicum) and the Italian white truffle in Southeast Asia in just the last five years [22,23,24]. Importantly, these three Tuber species have been grown in mycorrhizal association with the plant family Betulaceae, including Betula alnoides and Carpinus poilanei [22,23,24].
Environmental DNA (eDNA) revealed that the genetic material presented in the environmental samples, e.g., air, soil, sediment, and water, included both extracellular and intracellular DNA [25,26,27]. Notably, eDNA has been employed in a range of fundamental research studies in the fields of molecular biology, ecology, environmental science, and paleontology through high-throughput next-generation sequencing [26,27,28]. Recently, eDNA metabarcoding has become an important research tool and is now being commonly used to understand the microbial (including bacteria and fungi) diversity and community structure in a variety of environments [29,30,31,32]. Thus, soil mycobiota diversity has been provided to establish the ecological groups of fungi as mycorrhizal (mutualistic symbiotic association of fungi and plants), saprotrophic (decomposers), and pathogenic (harmful or parasitic to other organisms) [33,34,35,36]. Several previous studies have been conducted to evaluate and monitor soil fungal diversity and fungal communities, as well as to detect rare fungal species in soil samples collected from various forest types during different seasons with the use of eDNA [36,37,38]. In previous decades, species-specific primers for polymerase chain reactions (PCR) were used to detect truffles in the environment [39,40,41]; however, a notable disadvantage of this method was that it requires a large amount of extracted DNA to be present in the samples. Consequently, a number of unexpected truffle species were missed [42,43]. Due to the fact that trained animals have always been critical to the process of truffle hunting, truffle collection in Thailand has been limited. In this study, the soil mycobiota diversity of truffle host plants (B. alnoides and C. poilanei) in northern Thailand were investigated using eDNA metabarcoding (ITS1 region of rDNA). This was done for the purposes of developing the capability of investigating truffle diversity and locating truffles in their natural habitats during the non-fruiting phase (the dry season). The ITS sequence (ITS1 and ITS2 regions) has been used for the fungal metabarcoding [44,45]. However, the ITS1 region has been widely used as a universal fungal barcode for the quick and general analyses of diversity and ITS1 outperforms ITS2 in terms of richness and taxonomic coverage [46,47,48]. Thus, we selected the ITS1 region for this study. According to the information gathered from the DNA of truffles in soil samples taken during the wet to cold seasons, the hunt for truffle ascocarps was conducted in those seasons. Furthermore, the hunting for ascocarps of truffles was conducted during wet to cold seasons. The collected truffle ascocarps were identified by their morphological characteristics combined with a phylogenetic analysis. The results of this study will provide valuable information to researchers and truffle hunters and enable them to gain a better understanding of the distribution of truffles using soil mycobiota diversity investigation. The results could also be used to further develop relevant strategies for indicating truffle locations and searching truffles without trained pigs or dogs.
2. Materials and Methods
2.1. Ethics Statement
The collection of soil samples was permitted by the Department of National Parks, Wildlife and Plant Conservation, Bangkok, Thailand under document number 0907.4/4769.
2.2. Soil Sampling and eDNA Extraction
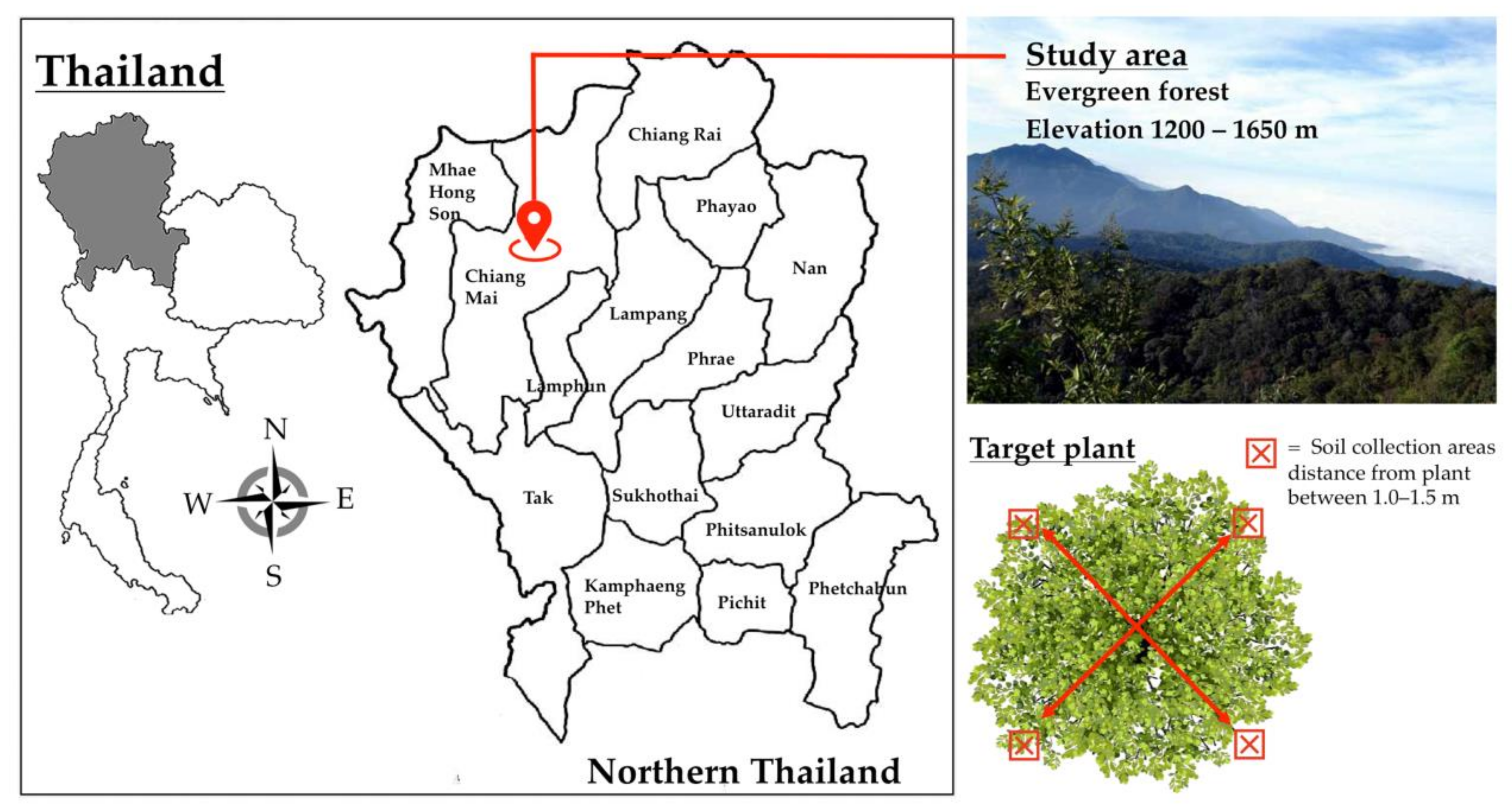
This study was conducted in the native evergreen forests (elevation 1200–1650 m) of Doi Suthep Mountain, Mueang Chiang Mai, Chiang Mai Province, northern Thailand (Figure 1). The study site was located in nature reserves within Doi Suthep-Pui National Park. The dominant tree genera in the study site were Castanopsis, Quercus, and Pinus. Notably, B. alnoides (n = 36) and C. poilanei (n = 2) were found within the study site (Supplementary Table S1). Thus, this study focused on the soil samples of B. alnoides and C. poilanei as host plants of truffles in northern Thailand [22,23,24]. Soil samples (n = 38) were collected from each host plant during the period of February to March, 2017 (the dry season and non-fruiting phase of the truffle) (Figure 1). Soil samples were randomly collected from four positions around target plants (Figure 1). After removing leaf litter, soil samples were aseptically collected at a depth of 5–10 cm from the surface of the soil and at a distance of 1.0–1.5 m from target plants using a soil core sampling tool (8.0 cm in diameter). Soil samples obtained from four positions of each target plant were pooled and kept in well-sealed sterile plastic bags. The samples were stored in an ice box and ultimately transported to the laboratory. All soil samples were air-dried at room temperature (27 ± 2 °C) immediately after reaching the laboratory. Each dried soil sample was ground using a mortar and pestle. The samples were then passed through a 250-µm sieve. All samples were stored at −80 °C until the process of eDNA extraction could be performed. Next, eDNA in the soil samples (each approximately 250 mg) was extracted using a NucleoSpin Soil DNA Isolation Kit (Macherey-Nagel, Germany) according to the manufacturers’ guidelines. All eDNA extracts were kept at −80 °C until further processing.
2.3. PCR Amplification, Amplicon Processing, and Illumina Sequencing
The eDNA extracts were used as templates for polymerase chain reactions (PCR) to amplify amplicons of the fungal ITS1 region of the rDNA gene using ITS5-1737F (5′-GGAAGTAAAAGTCGTAACAAGG-3′) and ITS2-2043R primers (5′-GCTGCGTTCT TCATCGATGC-3′) [44] that were linked to Illumina adapters. The amplification program was conducted with an initial denaturation step at 94 °C for 5 min, followed by 35 cycles of denaturation at 94 °C for 50 s annealing at 55 °C for 30 s and an extension step at 72 °C for 1 min with a final extension step at 72 °C for 5 min. PCR products were purified using a Qiagen Gel Extraction Kit (Qiagen, Hilden, Germany) and then sent to a commercial service provider (Novogene Bioinformatics Technology Co., Ltd., Beijing, China) for illumina sequencing.
2.4. PCR Sequence Analysis and Taxonomical Assignment
FLASH version 1.2.7 was used to assemble the forward and reverse sequences by overlapping paired-end reads [49]. The low-quality sequences (threshold of q30) were filtered out. The quality-filtering consisted of discarding reads with ambiguous sequences; sequence length ranges from 100 bp to 600 bp. Next, sequences that occurred only once (singletons) were discarded. Additionally, only the sequences belonging to the kingdom Fungi were kept for further analyses. Operational taxonomic units (OTUs) with a value greater than or equal to 97% similarity cut-off were clustered using UPARSE version 7.0.1001 [50]. The similarity threshold (≥97%) is commonly used in OTU-based analyses and has been shown to be an optimal threshold when using ITS to identify fungi [51]. The chimeric sequences were identified and removed with the UCHIME algorithm [52]. QIIME 1.7 was used for the purpose of taxonomy assignment with UNITE 7.2 fungal ITS reference training data set [53] and National Center for Biotechnology Information (NCBI) Taxonomy Database (http://0-www-ncbi-nlm-nih-gov.brum.beds.ac.uk/taxonomy, 25 May 2017) via the BLAST algorithm.
2.5. Truffle Hunting and Identification
In this study, truffle hunting was performed during the fruiting phase (the wet to cold seasons beginning from mid-May and extending to September in 2017) without the use of any trained pigs or dogs. Truffle ascocarps were searched and dug in both present and absence of truffle OTUs locations. The collected truffles were identified based on the morphological and molecular criteria. Morphological characteristics were observed for fresh specimens following the established method described in previous studies [15,22,54]. Genomic DNA of fresh specimens (10 mg) was extracted using a FavorPrep™ Tissue Genomic DNA Extraction Mini Kit (FAVORGEN, Taiwan). The ITS region was amplified with ITS4/ITS1F primers, under the following thermal conditions: 94 °C for 2 min; 35 cycles of 95 °C for 30 s, 52 °C for 30 s, and 72 °C for 1 min; and 72 °C for 10 min [22]. PCR products were checked and directly purified using a PCR Clean-up Kit (Macherey-Nagel, Germany). The purified PCR products were then sent to a commercial sequencing provider (1ST BASE Company, Kembangan, Malaysia). The sequences obtained from this study and other reference sequences obtained from GenBank were aligned using MUSCLE [55] and were then manually edited. Furthermore, jModelTest version 2.1.7 [56] was used to search for the selection of the best-fit nucleotide substitution models according to the Akaike Information Criterion. The phylogenetic tree was conducted using maximum likelihood (ML) and Bayesian inference (BI) algorithms and then implemented by RAxML version 7.0.3 [57] and MrBayes version 3.2.6 [58] following the method described in previous studies [3,22]. Bootstrap support (BS) and posterior probabilities (PP) values greater than or equal to 70% and 0.95, respectively, were significantly supported [59,60].
3. Results
3.1. General Soil Fungal Composition
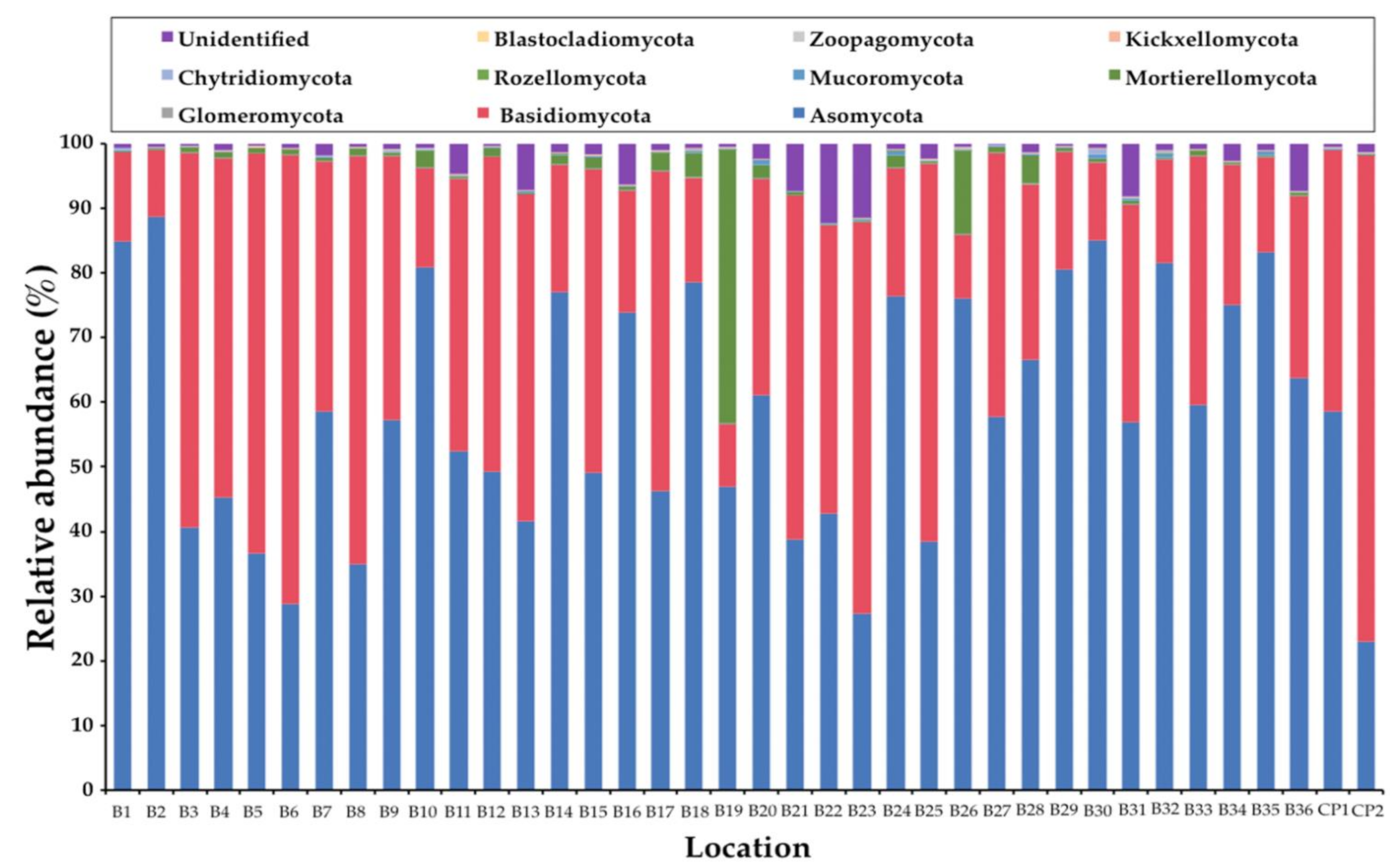
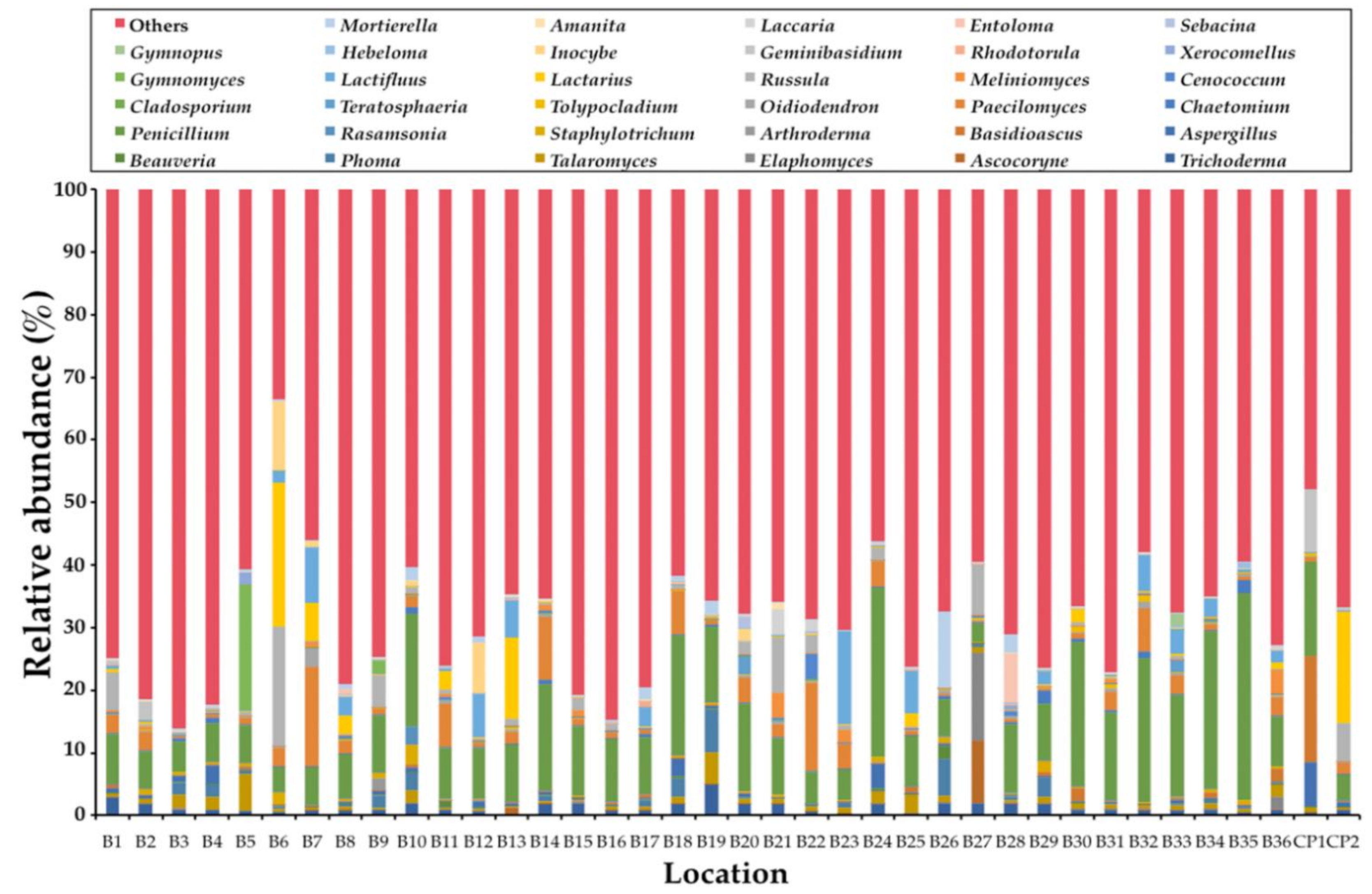
In this study, the obtained ITS1 sequences were not found to be fungal sequences within a range of 3% to 33%. This determination was dependent upon the samples (data not shown). Consequently, only the sequences belonging to the kingdom Fungi were analyzed. A total of 1,448,982 fungal amplicons from 38 soil samples were sequenced (Supplementary Table S2). The overall fungal community was assigned at the phylum-level as shown in Figure 1. The most abundant phylum was found in Ascomycota (57.63%) and followed by Basidiomycota (37.26%), unidentified (2.54%), Mortierellomycota (2.08%), Mucoromycota (0.24%), and Chytridiomycota (0.21%). Glomeromycota, Rozellomycota, and Kickxellomycota were found to be present in the same value at 0.01%. Subsequently, phyla Blastocladiomycota and Zoopagomycota were found at 0.007% and 0.003%, respectively. The abundant fungal phyla for each sample are presented in Figure 2. The relative degrees of abundance at the phylum-level varied in each soil sample. Ascomycota was represented at a range of 27.29% to 88.71% in soil samples of B. alnoides (n = 36) and 22.95% to 58.58% in soil samples of C. poilanei (n = 2). Twenty-three soil samples appear to be dominated by Ascomycota (>50%), of which the soil sample B2 revealed the highest degree of relative abundance. Soil samples of B. alnoides and C. poilanei revealed relative levels of abundance of Basidiomycota at 9.77% to 69.62% (n = 36) and 40.64% to 75.39% (n = 2), respectively. The highest relative abundance of Basidiomycota was found in sample B3. Notably, Chytridiomycota and Mortierellomycota were found in all samples. The highest relative abundance of Chytridiomycota (0.87%), Mortierellomycota (42.36%), and Mucoromycota (0.97%) was observed in soil samples B30, B19, and B20, respectively. Glomeromycota, Rozellomycota, Kickxellomycota, and Zoopagomycota were rarely found in each sample. Blastocladiomycota was only found in samples B5 and B15. However, Blastocladiomycota, Kickxellomycota, and Zoopagomycota were not observed in the soil samples of C. poilanei. Among the representative fungal amplicons, a final 1341 OTUs were obtained. The number of OTUs in Ascomycota, Basidiomycota, Blastocladiomycota, Chytridiomycota, Glomeromycota, Kickxellomycota, Mortierellomycota, Mucoromycota, Rozellomycota, Zoopagomycota, and unidentified were recorded at 811, 425, 2, 28, 8, 1, 19, 23, 14, 2, and 8, respectively (Supplementary Table S3). For deeper taxonomic assignments, a total of 35 classes, 99 orders, 240 families, and 549 genera were identified (Supplementary Table S4). It was found that the fungal genera varied in different soil samples. The relative abundance of the dominant 35 fungal genera among all soil samples is presented in Figure 3. Of the 35 most abundant fungal genera in the soil samples, the most abundant genera of mycorrhizal fungi were Amanita, Cenococcum, Elaphomyces, Gymnomyces, Hebeloma, Inocybe, Lactifluus, Lactarius, Laccaria, Meliniomyces, Russula, and Xerocomellus. The remaining 23 most abundant genera were identified as soil saprotrophs. Interestingly, the expected genus Tuber was also found in most of the soil samples. Additionally, the relative abundance of the genus Tuber in each soil sample is presented in Table 1. It was found that the relative abundance of this genus varied within a range of 0.01% to 4.00%. The highest relative abundance of the genus Tuber was found in sample B20.
3.2. OTUs of Truffles (Tuber Species) in Soil Samples
The results revealed that six OTUs representatively belonged to the genus Tuber (Table 2). OTU162, OTU187, OTU447, and OTU530 belonged to T. thailandicum, T. lannaense, T. bomiense, and T. magnatum, whereas OTU105 and OTU720 were unrecognized Tuber species. From 38 locations, truffle OTUs were not detected in five soil samples (B18, B19, B25, B30, and B35 locations) (Table 2). The remaining 33 locations were determined to be representative locations of truffle OTUs. Theses 33 locations included three previously known truffle locations (B2, B27, and CP2). Tuber thailandicum and T. magnatum were previously found in the locations B2 and CP2, respectively, while T. lannaense was previously found at the locations B27 and CP2 [22,23,24]. Therefore, 30 locations were identified as newly found truffle locations within our study area (Table 1). Twenty-three and three locations of B. alnoides were considered new locations for T. thailandicum and T. bomiense, respectively. Two locations of B. alnoides (B32 and B36) and one location of C. poilanei (CP1) indicated that they were new locations for T. lannaense. However, a new location for T. magnatum was not found. Additionally, eight new locations of B. alnoides were indicated for the unrecognized Tuber species by the presence of OTU105 and OTU720.
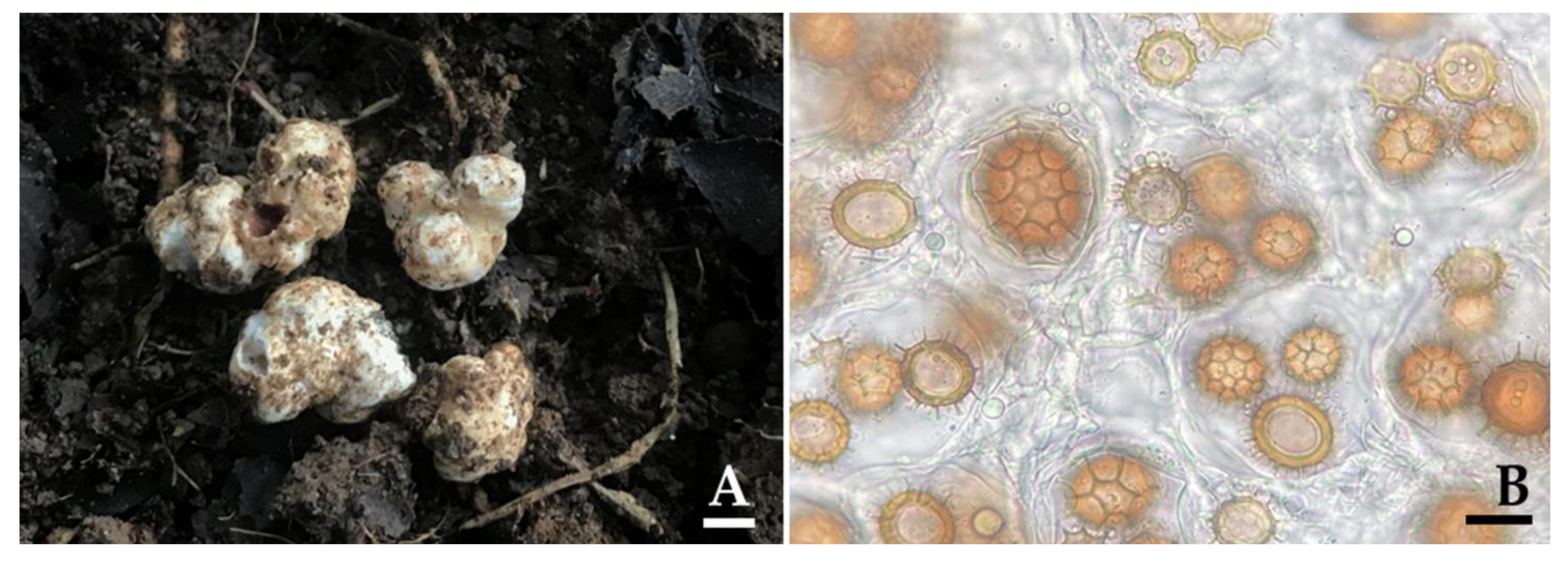
3.3. Truffle Hunting and Identification
Truffle ascocarps were located following the results obtained from the eDNA data (Table 1) during the period from mid-May to September 2017. The results indicated that truffle ascocarps were found at 18 locations from a total of 30 locations, of which 16 locations were determined to be new truffle locations (Figure 4). However, truffle ascocarps were not found in the remaining 14 new locations. Truffle ascocarps were also not found in the absence of truffle OTUs locations. It was found that all 18 truffle ascocarps were obtained from the soil of B. alnoides. The morphological characteristics of 18 ascocarps that were collected from different locations are shown in Table 3. The results indicate that the morphological characteristics of ascocarps at each location were similar. Morphologically, all 18 ascocarps from each location were found to be quite similar to T. thailandicum. Ascocarps were 1.0−4.5 cm in diameter and white to light brown in color. Ascus contained one to four (rarely five) ascospores. Ascospores were globose to subglobose, sometimes broadly ellipsoid, 20–65 × 18–62 µm ornamented with a regular alveolate reticulum. Identification was confirmed by molecular phylogenetic analysis. The ITS sequence from ascocarps in B1, B2, B4, B5, B7, B10, B12, B13, B14, B15, B22, B26, B27, B29, B31, B32, B33, and B36 locations were deposited in GenBank under the numbers MW465642, MW465641, MW471002, MW471000, MW471001, MW470998, MW470999, MW470961, MW470960, MW470969, MW470965, MW470963, MW470962, MW470968, MW470964, MW470966, MW470967, and MW470970, respectively. For the phylogenetic analysis, sequences of 66 Tuber taxa were used, and T. magnatum was used as the outgroup. The alignment was deposited in TreeBASE under the study ID number 27568. A best scoring RAxML tree was established with a final ML optimization likelihood value of −8556.2711. The proportion of invariable sites and gamma distribution were 0.3890 and 0.5670, respectively. The average standard deviation of the split frequencies of the BI analysis is 0.00914.
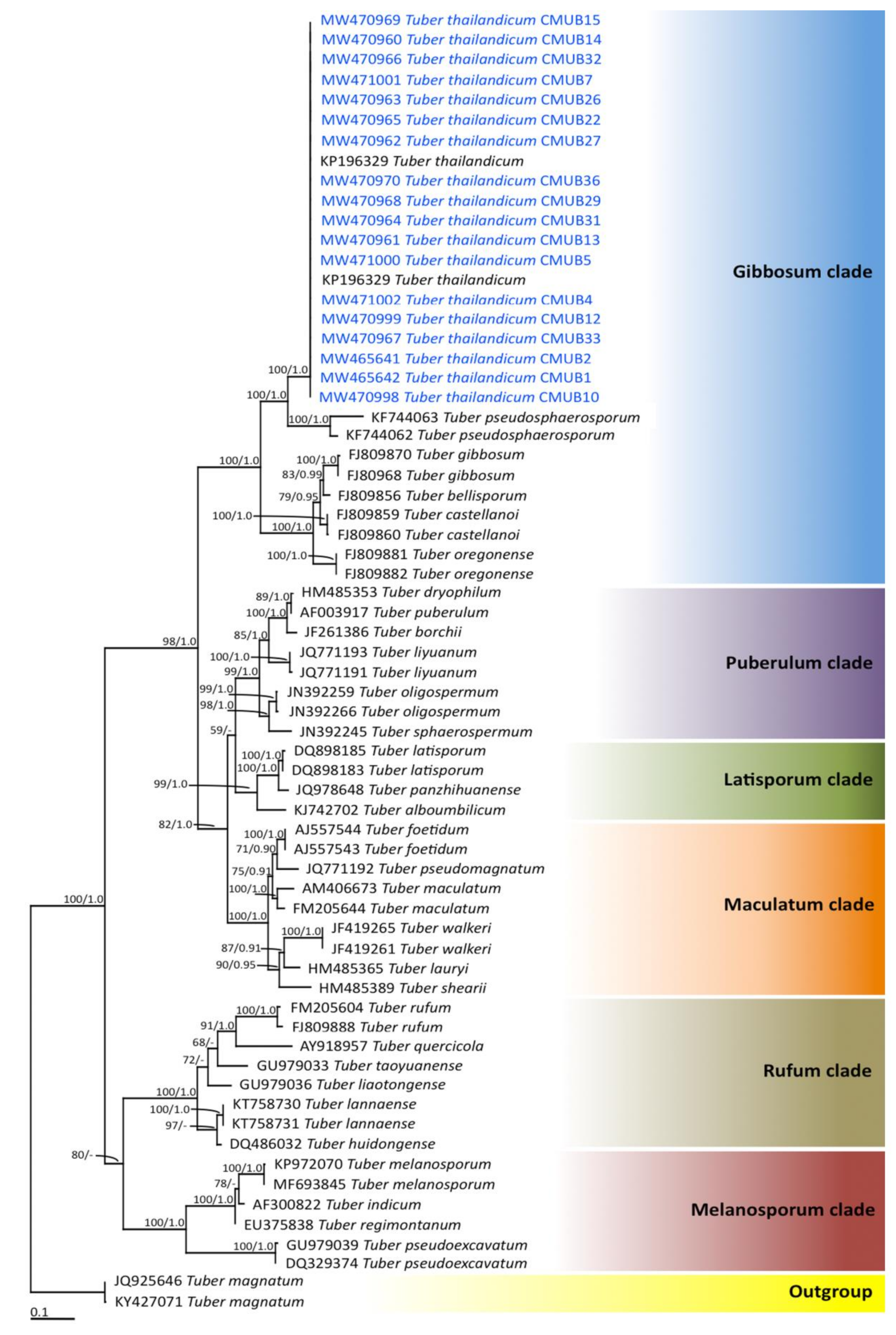
The phylograms of the ML and BI analyses were found to be similar in topology (data not shown). Therefore, we have only presented the phylogram obtained from the ML analysis (Figure 5). Six main clades, namely Gibbosum, Puberulum, Latisporum, Maculatum, Rufum, and Melanosporum, were assigned according to the findings of previous phylogenetic studies [3,61,62]. Our phylogenetic results place all the specimens obtained from each location within the monophyletic clade of T. thailandicum in the Gibbosum clade with high support values (100% BS and 1.0 PP). Therefore, all truffle ascocarps obtained from each location were identified to T. thailandicum based on morphological and molecular data. However, ascocarps of other truffle species were not found at the new representative locations of the OTUs.
4. Discussion
An investigation of soil mycobiota diversity in forest soil using fungal metabarcoding analysis has provided valuable information on the variety of distinct functional groups of fungi, including mycorrhizal, saprotrophic, and pathogenic fungi [33,63,64,65]. We have hypothesized that the soil mycobiota diversity of the host plants of truffles could be used to investigate truffle diversity and to assist in locating truffles during the non-fruiting phase. In this study, soil samples obtained during the non-fruiting phase from known truffle host plants (B. alnoides and C. poilanei) of northern Thailand were collected, while soil mycobiota diversity was investigated. Our results indicate that Ascomycota had the highest overall relative abundance at the phylum level in the soil samples followed by Basidiomycota, Mortierellomycota, and Mucoromycota. Similarly, Ascomycota was found to be the dominant phylum in the soil of evergreen and deciduous forests in northeast Thailand [64]. Moreover, these results were supported by the findings of previous studies, which reported that Ascomycota was associated with high levels of distribution and abundance in global soil samples [66,67,68,69,70]. Our findings are similar to the findings of a number of previous studies [34,70,71,72,73,74,75] that found that Inocybe, Lactarius, Meliniomyces, and Russula are ectomycorrhizal-dominated fungal communities in the soil samples of tropical, temperate and boreal forests. Moreover, Cortinarius, Piloderma, and Suillus were also found to be dominant, especially in boreal forests [68,71,72]. In addition, Meliniomyces have been observed to be able to form both ectomycorrhiza and ericoidmycorrhiza [76,77]. Notably, certain previously published studies have reported that the geographical region, the specific season, the dominant tree species, nutrient availability, and the physical and chemical properties of the soil and soil depth all significantly affected the composition of the fungal community in the soil [37,68,71,78,79].
Metabarcoding studies on fungal diversity in natural environments have provided not only the functional groups of fungal compositions, but deeper insight into the compositions of the fungal species. In this study, six OTUs that were representative of the DNA of truffles included four known species (T. bomiense, T. lannaense, T. magnatum, and T. thailandicum). Additionally, two unrecognized truffle specimens were revealed by an investigation of the soil mycobiota diversity of B. alnoides and C. poilanei. Accordingly, several previous studies have used metagenomic analysis to monitor and detect some expected fungal genus and species in the environment [31,38,39,80,81]. Examples of these include the studies of Bai et al. [82] and Sommermann et al. [83], who used fungal metabarcoding data in agricultural soil to monitor and detect soil fungal pathogens. The distribution of truffle-like taxa (Mesophellia, Hysterangium, and Chondrogaster) in north-east Australian woodlands has been monitored by the mycorrhizal fungal community structure in root and soil samples [75]. Nowadays, several studies have applied the metabarcoding technique in the research and cultivation of truffles, including for the detection of T. aestivum and T. melanosporum in truffle orchards [84], the monitoring of the persistence of mycorrhization of T. melanosporum and T. indicum in the roots of their host plants [85,86], and a greater understanding of the relationship between truffles and other soil microorganisms (bacteria and fungi) for the improvement of the cultivation of several truffle species (T. aestivum, T. indicum, T. pseudoexcavatum, and T. sinoaestivum) along with their ascocarp development [85,87,88,89]. Consequently, the method of detection for the DNA of truffles in soil samples can be used in the investigation of truffle diversity and distribution in their natural habitat, as well as in the hunting of truffles during their fruiting period. As we might expect, the use of this method has resulted in the discovery of new species and new records of truffles in northern Thailand. Thus, the detection of ascocarps will help to prove and confirm the presence of each truffle species. Thirty locations were considered new locations for T. bomiense, T. thailandicum, while other unrecognized Tuber species and ascocarps of T. thailandicum were found that were undergoing the fruiting phase. Our results indicate that the use of the composition of fungal communities in soil samples can offer a new tool for the investigation and hunting of truffles. However, ascocarps of other truffle species were not found at the new OTUs representative locations. We found only the DNA of T. bomiense in the soil samples, but its ascocarps were not detected. Therefore, there is still a need to further hunt truffles in this study area. Furthermore, ascocarps of T. lanaense and T. magnatum were also not detected at previously known locations during our hunting period. Several previous studies have reported that multiple biotic and abiotic factors (e.g., temperature, humidity, soil nutrients, mating type, and soil microbial community) [2,4,9,90,91,92]. The suitable biotic and abiotic factors can influence ascocarp development in truffles. Moreover, seasonal climatic changes that occur during each year significantly affect ascocarp development in truffles [2,4,7,9]. According to the outcomes of our previous studies, ascocarps of T. thailanicum were recorded from mid-May to June [22] and ascocarps of both T. lannaense and T. magnatum were recorded from July to August [23,24], during which the humidity was recorded within a range of 70−75% and temperatures were reported within a range of 28−30 °C. From mid-May to June 2017, rain was observed in the study area and relative humidity was recorded within a range of 72−75%. Furthermore, temperatures were reported within a range of 28−30 °C. These conditions clearly supported the ascocarp development of truffles. However, from July to August, minimal amounts of rain were observed with low relative humidity readings within the range of 62−68% and high temperatures within the range of 29−33 °C (Northern Meteorological Center; http://www.cmmet.tmd.go.th, 30 June 2017). It is possible that during our hunting period, suitable conditions for the ascocarp development of other truffle species did not exist, with the exception of T. thailandicum. Notably, our study had two main limitations: (1) the absence of trained animals for truffle hunting and (2) a short-term period of observation. Consequently, future studies should focus on an evaluation of the efficiency of methods of truffle hunting using fungal community data, trained animals, and a combination of fungal community data and trained animals. Furthermore, long-term observations should be employed to provide an even greater understanding of the discovery and distribution of truffles, as well as to offer an expanded understanding of their fruiting periods within the study area.
5. Conclusions
Truffles form hypogeous ascocarps that are difficult to locate without the use of trained pigs and dogs. This study is the first of its kind to investigate the composition of the fungal community in soil samples of the host plants of truffles (B. alnoides and C. poilanei) in the native forests of northern Thailand. This was done with the objective of investigation of truffle diversity and locating truffles during the non-fruiting phase. From detected soil fungi, six OTUs that are representative of the DNA of truffles were found to include four known species (T. bomiense, T. lannaense, T. magnatum, and T. thailandicum) and two unrecognized truffle species. Thirty locations were considered new truffle locations, while truffle ascocarps were identified underground during the fruiting phase in 16 new locations. However, no truffle ascocarps were found in the remaining 14 new locations during the course of our investigation. All obtained truffles were identified as T. thailandicum based on the morphological and molecular data. There is still a need to further hunt truffles in our study area. Our study demonstrated that soil mycobiota diversity could be offered as a new tool in the investigation of truffle diversity and hunting for truffles without the use of trained animals. To the best of our knowledge, the results of this study can provide valuable information to researchers in terms of establishing a greater understanding of the distribution of truffles in their natural habitats based on soil mycobiota diversity. This would ultimately benefit the management and conservation of truffle habitats in northern Thailand as well as the future cultivation of truffles. Additionally, the outcomes of this study could facilitate a more efficient search for truffles and ultimately indicate their location. Further studies on the search for other truffle species from our describing soil mycobiota diversity are required in conjunction with long-term observations. A combination of using metabarcoding technics and trained animals in the hunt for truffles is needed in the future to evaluate the efficacy of these newly established truffle hunting methods. Moreover, soil mycobiota diversity of different plant species (e.g., chestnut, oak, and pine) should be investigated to fully understand the distribution of truffles in the natural forests of northern Thailand.
Supplementary Materials
The following are available online at https://0-www-mdpi-com.brum.beds.ac.uk/article/10.3390/jof7040293/s1, Table S1: Location of host plant in study site, Table S2: Soil fungal sequence reads from each soil sample organized by phylum-level, Table S3: The OTUs obtained in this study, Table S4: The fungal taxonomic assignments in this study.
Author Contributions
Conceptualization: N.S., J.K. and S.L.; methodology: N.S. and J.K.; software: A.I.-o.; validation: N.S., J.K., A.I.-o. and S.L.; formal analysis: J.K., N.S. and A.I.-o.; investigation: N.S.; resources: N.S. and J.K.; data curation: N.S. and J.K.; writing—original draft: N.S. and J.K.; writing—review and editing: N.S., J.K. and S.L.; supervision: N.S. and S.L. All authors have read and agreed to the published version of the manuscript.
Funding
This work supported by the National Science and Technology Development Agency (FDA-CO-2560-4850-TH), and partially supported by Chiang Mai University, Thailand.
Institutional Review Board Statement
Not applicable.
Informed Consent Statement
Not applicable.
Data Availability Statement
The DNA sequence data obtained from this study have been deposited in GenBank under accession numbers; MW330007, MW326971, MW330257, MW327594, MW326781, MW326083MW465642, MW465641, MW471002, MW471000, MW471001, MW470998, MW470999, MW470961, MW470960, MW470969, MW470965, MW470963, MW470962, MW470968, MW470964, MW470966, MW470967, and MW470970.
Acknowledgments
The authors are grateful to Russell Kirk Hollis for the English correction of this manuscript.
Conflicts of Interest
The authors declare that they have no conflict of interest.
References
- Kirk, P.M.; Cannon, P.F.; Minter, D.W.; Stalpers, J.A. Dictionary of the fungi, 10th ed.; CAB International: Wallingford, UK, 2008. [Google Scholar]
- Hall, I.R.; Brown, G.T.; Zambonelli, A. Taming the Truffle, the History, Lore and Science of the Ultimate Mushroom; Timber Press: Portland, OR, USA, 2007. [Google Scholar]
- Bonito, G.; Grygansyi, A.P.; Trappe, J.M.; Vilgalys, R. A global metaanalysis of Tuber ITS rDNA sequences: Species diversity, host associations and long-distance dispersal. Mol. Ecol. 2010, 19, 4994–5008. [Google Scholar] [CrossRef] [PubMed]
- Zambonelli, A.; Iotti, M.; Murat, C. True Truffle (Tuber spp.) in the World, 1st ed.; Springer International Publishing AG: Basel, Switzerland, 2016. [Google Scholar]
- Splivallo, R.; Valdez, N.; Kirchhoff, N.; Ona, M.C.; Schmidt, J.P.; Feussner, I.; Karlovsky, P. Intraspecific genotypic variability determines concentrations of key truffle volatiles. New Phytol. 2012, 194, 823–835. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Splivallo, R.; Ottonello, S.; Mello, A.; Karlovsky, P. Truffle volatiles: From chemical ecology to aroma biosynthesis. New Phyt. 2011, 189, 688–699. [Google Scholar] [CrossRef] [PubMed]
- Trappe, J.M.; Molina, R.; Luoma, D.L.; Cázares, E.; Pilz, D.; Smith, J.E.; Castellano, M.A.; Miller, S.L.; Trappe, M.J. Diversity, ecology, and conservation of truffle fungi in forests of the pacific northwest; Timber Press: Portland, OR, USA, 2009. [Google Scholar]
- Stobbe, U.; Egli, S.; Tegel, W.; Peter, M.; Sproll, L.; Büntgen, U. Potential and limitation of Burgundy truffle cultivation. Appl. Microbiol. Biotechnol. 2013, 97, 5215–5224. [Google Scholar] [CrossRef]
- Hall, I.R.; Yun, W.; Amicucci, A. Cultivation of edible ectomycorrhizal mushrooms. Trends Biotechnol. 2013, 21, 433–438. [Google Scholar] [CrossRef]
- Bonito, G.; Trappe, J.M.; Rawlimson, P.; Vilgalys, R. Improved resolution of major clades whitish Tuber and taxonomy of species within the Tuber gibbosum complex. Mycologia 2010, 102, 1042–1057. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Smith, M.E.; Bonito, G.; Sharma, J.; Long, J.; Davis-Long, B.; Brenneman, T. Pecan Truffles (Tuber lyonii) What We Know and What We Need to Know. Georgia Pecan Magazine 2012, 52–58. [Google Scholar]
- Mortimer, P.E.; Karunarathna, S.C.; Li, Q.; Gui, H.; Yang, X.; Yang, X.; He, J.; Ye, L.; Guo, J.; Li, H.; et al. Prized edible Asian mushrooms: Ecology, conservation and sustainability. Fungal Divers. 2012, 56, 31–47. [Google Scholar] [CrossRef]
- Chen, Y.L.; Gong, M.Q. Truffles Tuber spp.: Biodiversity and geological distribution. Edible Fungi of China 2000, 19, 25–26. [Google Scholar]
- Chen, J.; Liu, P.G.; Wang, Y. Tuber umbilicatum, a new species from China, with a key to the spinose-reticulate spored Tuber species. Mycotaxon 2005, 94, 1–6. [Google Scholar]
- Deng, X.J.; Liu, P.G.; Liu, C.Y.; Wang, Y. A new white truffle species, Tuber panzhihuanense from China. Mycol. Prog. 2013, 12, 557–561. [Google Scholar] [CrossRef]
- Fan, L.; Cao, J.Z. Two new species of white truffle from China. Mycotaxon 2012, 121, 297–304. [Google Scholar] [CrossRef]
- García-Montero, L.G.; Díaz, P.; Massimo, G.D.; García-Abril, A. A review of research on Chinese Tuber species. Mycol. Prog. 2010, 9, 315–335. [Google Scholar] [CrossRef]
- Kinoshima, A.; Sasaki, H.; Nara, K. Two new truffle species, Tuber japonicum and Tuber flavidosporum spp. nov. found from Japan. Mycoscience 2016, 57, 366–373. [Google Scholar] [CrossRef]
- Fan, L.; Liu, X.; Cao, J. Tuber turmericum sp. nov., a Chinese truffle species based on morphological and molecular data. Mycol. Prog. 2015, 14, 111. [Google Scholar] [CrossRef]
- Kinoshita, A.; Sasaki, H.; Nara, K. Phylogeny and diversity of Japanese truffles (Tuber spp.) inferred from sequences of four nuclear loci. Mycologia 2011, 103, 779–794. [Google Scholar] [CrossRef] [PubMed]
- Hyde, K.D.; Norphanphoun, C.; Chen, J.; Dissanayake, A.J.; Doilom, M.; Hongsanan, S.; Jayawardena, R.S.; Jeewon, R.; Perera, R.H.; Thongbai, B.; et al. Thailand’s amazing diversity–up to 96% of fungi in northern Thailand are novel. Fungal Divers. 2018, 93, 215–239. [Google Scholar] [CrossRef]
- Suwannarach, N.; Kumla, J.; Lumyong, S. A new whitish truffle, Tuber thailandicum from northern Thailand and its ectomycorrhizal association. Mycol. Prog. 2015, 14, 83. [Google Scholar] [CrossRef]
- Suwannarach, N.; Kumla, J.; Vadthanarat, S.; Raspé, O.; Lumyong, S. Morphology and molecular evidence support a new truffle. Tuber lannaense, from Thailand. Mycol. Prog. 2016, 15, 827–834. [Google Scholar]
- Suwannarach, N.; Kumla, J.; Lumyong, S. Tuber magnatum in Thailand, a first report from Asia. Mycotaxon 2017, 132, 635–642. [Google Scholar] [CrossRef]
- Ficetola, G.F.; Thuiller, W.; Miaud, C. Prediction and validation of the potential global distribution of a problematic alien invasive species-the American bullfrog. Divers. Distrib. 2007, 13, 476–485. [Google Scholar] [CrossRef]
- Nagler, M.; Insam, H.; Pietramellara, G.; Ascher-Jenull, J. Extracellular DNA in natural environments: Features, relevance and applications. Appl. Microbiol. Biotechnol. 2018, 102, 6343–6356. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Thomsen, P.F.; Willerslev, E. Environmental DNA—an emerging tool in conservation for monitoring past and present biodiversity. Biol. Conserv. 2015, 183, 4–18. [Google Scholar] [CrossRef]
- Taberlet, P.; Coissac, E.; Hajibabaei, M.; Rieseberg, L.H. Environmental DNA. Mol. Ecol. 2012, 21, 1789–1793. [Google Scholar] [CrossRef] [PubMed]
- Epp, L.S.; Boessenkool, S.; Bellemain, E.P.; Haile, J.; Esposito, A.; Riaz, T.; Erseus, C.; Gusarov, V.I.; Edwards, M.E.; Johnsen, A. New environmental metabarcodes for analysing soil DNA: Potential for studying past and present ecosystems. Mol. Ecol. 2012, 21, 1821–1833. [Google Scholar] [CrossRef] [PubMed]
- Ruppert, K.M.; Kline, R.J.; Rahman, M.S. Past, present, and future perspectives of environmental DNA (eDNA) metabarcoding: A systematic review in methods, monitoring, and applications of global eDNA. Glob. Ecol. Conserv. 2019, 17, e00547. [Google Scholar] [CrossRef]
- Soliman, T.; Yang, S.Y.; Yamazaki, T.; Jenke-Kodama, H. Profiling soil microbial communities with next-generation sequencing: The influence of DNA kit selection and technician technical expertise. PeerJ. 2017, 5, e4178. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Deiner, K.; Walser, J.; Machler, E.; Altermatt, F. Choice of capture and extraction methods affect detection of freshwater biodiversity from environmental DNA. Biol. Conserv. 2015, 183, 53–63. [Google Scholar] [CrossRef]
- Uroz, S.; Oger, P.; Tisserand, E.; Cebron, A.; Turpault, M.P.; Buee, M.; de Boer, W.; Leveau, J.H.; Frey-Klett, P. Specific impacts of beech and Norway spruce on the structure and diversity of the rhizosphere and soil microbial communities. Sci. Rep. 2016, 6, 27756. [Google Scholar] [CrossRef] [PubMed]
- Wu, D.; Zhang, M.; Peng, M.; Sui, X.; Li, W.; Sun, G. Variations in soil functional fungal community structure associated with pure and mixed plantations in typical temperate forests of China. Front. Microbiol. 2019, 10, 1636. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tedersoo, L.; Bahram, M.; Cajthaml, T.; Polme, S.; Hiiesualu, I.; Anslan, S.; Harend, H.; Buegger, F.; Pritsch, K.; Koricheva, J.; et al. Tree diversity and species identity effects on soil fungi, protists and animals are context dependent. ISME J. 2016, 10, 346–362. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Landinez-Torres, A.; Panelli, S.; Picco, A.M.; Comandatore, F.; Tosi, S.; Capelli, E. A meta-barcoding analysis of soil mycobiota of the upper Andean Colombian agro-environment. Sci. Rep. 2019, 9, 10085. [Google Scholar] [CrossRef] [Green Version]
- O’Brien, H.E.; Parrent, J.L.; Jackson, J.A.; Moncalvo, J.M.; Vilgalys, R. Fungal community analysis by large-scale sequencing of environmental samples. Appl. Environ. Microbiol. 2005, 71, 5544–5550. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lindahl, B.D.; Nilsson, R.H.; Tedersoo, L.; Abarenkov, K.; Carlsen, T.; Kjøller, R.; Kõljalg, U.; Pennanen, T.; Rosendahl, S.; Stenlid, J.; et al. Fungal community analysis by high-throughput sequencing of amplified markers--a user’s guide. New Phytol. 2013, 199, 288–299. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Iotti, M.; Amicucci, A.; Bonito, G.; Bonuso, E.; Stocchi, V.; Zambonelli, A. Selection of a set of specific primers for the identification of Tuber rufum: A truffle species with high genetic variability. FEMS Microbiol. Lett. 2007, 277, 223–231. [Google Scholar] [CrossRef] [PubMed]
- Sejalon-Delmas, N.; Roux, C.; Martins, M.; Kulifaj, M.; Becard, G.; Dargent. Molecular tools for the identification of Tuber melanosporum in agroindustry. J. Agric. Food Chem. 2000, 48, 2608–2613. [Google Scholar] [CrossRef] [PubMed]
- Gryndler, M.; Hršelová, H.; Soukupová, L.; Streiblová, E.; Valda, S.; Borovička, J.; Gryndlerová, H.; Gažo, J.; Miko, M. Detection of summer truffle (Tuber aestivum Vittad.) in ectomycorrhizae and in soil using specific primers. FEMS Microbiol. Lett. 2011, 318, 84–91. [Google Scholar] [CrossRef]
- Herder, J.; Valentin, A.; Bellemain, E.; Dejean, T.; van Delft, J.; Thomsen, P.F.; Taberlet, P. Environmental DNA a Review of the Possible Applications for the Detection of (Invasive) Species; Stichting RAVON: Nijmegen, The Netherlands, 2014. [Google Scholar]
- Horisawa, S.; Sakuma, Y.; Doi, S. Qualitative and quantitative PCR methods using species-specific primer for detection and identification of wood rot fungi. J. Wood Sci. 2009, 55, 133–138. [Google Scholar] [CrossRef]
- Bellemain, E.; Carlsen, T.; Brochmann, C.; Coissac, E.; Taberlet, P.; Kauserud, H. ITS as an environmental DNA barcode for fungi: An in silico approach reveals potential PCR biases. BMC Microbiol. 2010, 10, 189. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Schoch, C.L.; Seifert, K.A.; Huhndorf, S.; Robert, V.; Spouge, J.L.; Levesque, C.A. Nuclear ribosomal internal transcribed spacer (ITS) region as a universal DNA barcode marker for Fungi. Proc. Natl. Acad. Sci. USA 2012, 109, 6241–6246. [Google Scholar] [CrossRef] [Green Version]
- Mbareche, H.; Veillette, M.; Bilodeau, G.; Duchaine, C. Comparison of the performance of ITS1 and ITS2 as barcodes in amplicon-based sequencing of bioaerosols. PeerJ 2020, 8, e8523. [Google Scholar] [CrossRef] [Green Version]
- Wang, X.C.; Liu, C.; Huang, L.; Bengtsson-Palme, J.; Chen, H.; Zhang, J.H.; Cai, D.; Li, J.Q. ITS1: A DNA barcode better than ITS2 in eukaryotes? Mol. Ecol. Resour. 2015, 15, 573–586. [Google Scholar] [CrossRef]
- Nilsson, R.H.; Kristiansson, E.; Ryberg, M.; Hallenberg, N.; Larsson, K.-H. Intraspecific ITS variability in the kingdom Fungi as expressed in the international sequence databases and its implications for molecular species identification. Evol. Bioinform. 2008, 4, EBO–S653. [Google Scholar] [CrossRef]
- Magoč, T.; Salzberg, S.L. FLASH: Fast length adjustment of short reads to improve genome assemblies. Bioinformatics 2011, 27, 2957–2963. [Google Scholar] [CrossRef]
- Edgar, R.C. UPARSE: Highly accurate OTU sequences from microbial amplicon reads. Nat. Methods. 2013, 10, 996–998. [Google Scholar] [CrossRef] [PubMed]
- Koljalg, U.; Nilsson, R.H.; Abarenkov, K.; Tedersoo, L.; Taylor, A.F.S.; Bahram, M.; Bates, S.T.; Bruns, T.D.; Bengtsson-Palme, J.; Callaghan, T.M.; et al. Towards a unified paradigm for sequence-based identification of fungi. Mol. Ecol. 2013, 22, 5271–5277. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Edgar, R.C.; Haas, B.J.; Clemente, J.C.; Quince, C.; Knight, R. UCHIME improves sensitivity and speed of chimera detection. Bioinformatics 2011, 27, 2194–2200. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Caporaso, J.G.; Kuczynski, J.; Stombaugh, J.; Bittinger, K.; Bushman, F.D.; Costello, E.K.; Fierer, N.; Peña, A.G.; Goodrich, J.K.; Gordon, J.I.; et al. QIIME allows analysis of high-throughput community sequencing data. Nat. Methods. 2010, 7, 335–336. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, S.H.; Zheng, L.Y.; Liu, C.Y.; Wang, Y.; Li, L.; Zhao, Y.C.; Zhang, X.L.; Yang, M.; Xiong, H.K.; Qing, Y.; et al.; et al. Two new truffles species, Tuber alboumbilicum and Tuber pseudobrumale from China. Mycol. Prog. 2014, 13, 1157–1163. [Google Scholar] [CrossRef]
- Edgar, R.C. MUSCLE: Multiple sequence alignment with high accuracy and high throughput. Nucleic Acids Res. 2004, 32, 1792–1797. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Darriba, D.; Taboada, G.L.; Doallo, R.; Posada, D. jModelTest 2: More models, new heuristics and parallel computing. Nat. Methods 2012, 9, 772. [Google Scholar] [CrossRef] [Green Version]
- Stamatakis, A. RAxML-VI-HPC: Maximum likelihood-based phylogenetic analyses with thousands of taxa and mixed models. Bioinformatics 2006, 22, 2688–2690. [Google Scholar] [CrossRef]
- Ronquist, F.; Teslenko, M.; van der Mark, P.; Ayres, D.L.; Darling, A.; Höhna, S.; Larget, B.; Liu, L.; Suchard, M.A.; Huelsenbeck, J.P. MrBayes 3.2: Efficient Bayesian phylogenetic inference and modelchoice across a large model space. Sys. Biol. 2012, 61, 539–542. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hillis, D.M.; Bull, J.J. An empirical test of bootstrapping as a method for assessing confidence in phylogenetic analysis. Syst. Biol. 1993, 42, 182–192. [Google Scholar] [CrossRef]
- Alfaro, M.E.; Zoller, S.; Lutzoni, F. Bayes or bootstrap? A simulation study comparing the performance of Bayesian Markov Chain Monte Carlo sampling and bootstrapping in assessing phylogenetic confidence. Mol. Biol. Evol. 2003, 20, 255–266. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bonito, G.; Smith, M.E.; Nowak, M.; Healy, R.A.; Guevara, G.; Cázares, E.; Kinoshita, A.; Nouhra, E.R.; Domínguez, L.S.; Tedersoo, L.; et al. Historical biogeography and diversification of truffles in the Tuberaceae and their newly identified southern hemisphere sister lineage. PLoS ONE 2013, 8, e52765. [Google Scholar] [CrossRef]
- Polemis, E.; Konstantinidis, G.; Fryssouli, V.; Slavova, M.; Tsampazis, T.; Nakkas, V.; Assyov, B.; Kaounas, V.; Zervakis, G.I. Tuber pulchrosporum sp. nov., a black truffle of the Aestivum clade (Tuberaceae, Pezizales) from the Balkan peninsula. MycoKeys. 2019, 20, 35–51. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wubet, T.; Christ, S.; Schöning, I.; Boch, S.; Gawlich, M.; Schnabel, B.; Fischer, M.; Buscot, F. Differences in soil fungal communities between European beech (Fagus sylvatica L.) dominated forests are related to soil and understory vegetation. PLoS ONE 2012, 7, e47500. [Google Scholar] [CrossRef] [PubMed]
- Coince, A.; Caël, O.; Bach, C.; Lengellé, J.; Cruaud, C.; Gavory, F. Below-ground fine-scale distribution and soil versus fine root detection of fungal and soil oomycete communities in a French beech forest. Fungal Ecol. 2013, 6, 223–235. [Google Scholar] [CrossRef] [Green Version]
- Osona, T. Ecology of ligninolytic fungi associated with leaf litter decomposition. Ecol. Res. 2007, 22, 955–974. [Google Scholar] [CrossRef]
- Maestre, F.T.; Delgado-Baquerizo, M.; Jeffries, T.C.; Eldridge, D.J.; Ochoa, V.; Gozalo, B.; Quero, J.L.; García-Gómez, M.; Gallardo, A.; Ulrich, W.; et al. Increasing aridity reduces soil microbial diversity and abundance in global drylands. Proc. Natl. Acad. Sci. USA 2015, 112, 15684–15689. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jia, T.; Guo, T.; Yao, Y.; Wang, R.; Chai, B. Seasonal microbial community characteristic and its driving factors in a copper tailings dam in the Chinese Loess Plateau. Front. Microbiol. 2020, 11, 1574. [Google Scholar] [CrossRef] [PubMed]
- Nagati, M.; Roy, M.; Manzi, S.; Richard, F.; Desrocjers, A.; Gardes, M.; Bergeron, Y. Impact of local forest composition on soil fungal communities in a mixed boreal forest. Plant Soil 2018, 432, 345–357. [Google Scholar] [CrossRef]
- Egidi, E.; Delgado-Baquerizo, M.; Plett, J.M.; Wang, J.; Eldridge, D.J.; Bardgett, R.D.; Maestre, F.T.; Singh, B.K. A few Ascomycota taxa dominate soil fungal communities worldwide. Nat. Commun 2019, 10, 2369. [Google Scholar] [CrossRef] [Green Version]
- Brinkmann, N.; Schneider, D.; Sahner, J.; Ballauff, J.; Edy, N.; Barus, H.; Irawan, B.; Wilarso, S.; Qaim, M.; Daniel, R.; et al. Intensive tropical land use massively shifts soil fungal communities. Sci. Rep. 2019, 9, 3403. [Google Scholar] [CrossRef] [Green Version]
- Buée, M.; Reich, M.; Murat, C.; Morin, E.; Nilsson, R.H.; Uroz, S.; Martin, F. 454 Pyrosequencing analyses of forest soils reveal an unexpectedly high fungal diversity. New Phytol. 2009, 184, 449–456. [Google Scholar] [CrossRef] [PubMed]
- Sun, H.; Terhonen, E.; Kovalchuk, A.; Tuovila, H.; Chen, H.; Oghenekaro, A.O.; Heinonsala, J.; Kohler, A.; Kasanen, R.; Vasander, H.; et al. Dominant tree species and soil type affect the fungal community structure in a boreal peatland forest. Appl. Environ. Microbiol. 2016, 82, 2632–2643. [Google Scholar] [CrossRef] [Green Version]
- Männistö, M.; Vuosku, J.; Stark, S.; Saravesi, K.; Suokas, M.; Markkola, A.; Martz, F.; Rautio, P. Bacterial and fungal communities in boreal forest soil are insensitive to changes in snow cover conditions. FEMS Microbiol. Ecol. 2018, 94, 1–14. [Google Scholar]
- Shi, L.; Dossa, G.G.O.; Paudel, E.; Zang, H.; Xu, J.; Harrison, R.D. Changes in fungal communities across a forest disturbance gradient. Appl. Environ. Microbiol. 2019, 85, e00080-19. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nuske, S.J.; Anslan, S.; Tedersoo, L.; Congdon, B.C.; Abell, S.E. Ectomycorrhizal fungal communities are dominated by mammalian dispersed truffle-like taxa in north-east Australian woodlands. Mycrorrhiza 2019, 29, 181–193. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vuong, T.M.D.; Zeng, J.Y.; Man, X.L. Soil fungal and bacterial communities in southern boreal forests of the Greater Khingan Mountains and their relationship with soil properties. Sci. Rep. 2020, 10, 22025. [Google Scholar] [CrossRef]
- Hambleton, S.; Sigler, L. Meliniomyces, a new anamorph genus for root-associated fungi with phylogenetic affinities to Rhizoscyphus ericae (≡ Hymenoscyphus ericae), Leotiomycetes. Stud. Mycol. 2005, 53, 1–27. [Google Scholar] [CrossRef] [Green Version]
- Villarreal-Ruiz, L.; Neri-Luna, C.; Anderson, I.C.; Alexander, I.J. In vitro interactions between ectomycorrhizal fungi and ericaceous plants. Symbiosis 2012, 56, 67–75. [Google Scholar] [CrossRef]
- Gómez-Hernández, M.; Williams-Linera, G.; Guevara, R.; Lodge, D.J. Patterns of macromycete community assemblage along an elevation gradient: Options for fungal gradient and metacommunity analyse. Biodivers. Conserv. 2012, 21, 2247. [Google Scholar] [CrossRef]
- Cuadros-Orellana, S.; Leite, L.R.; Smith, A.; Medeiros, J.D.; Badotti, F.; Fonseca, P.L.C.; Vaz, A.B.; Oliveira, G.; Goes-Neta, G. Assessment of fungal diversity in the environment using Metagenomics: A decade in review. Fungal Genom. Biol. 2013, 3, 2. [Google Scholar]
- Wozniak, M.; Gradziel, J.; Galazka, A.; Frac, M. Metagaenomic analysis of bacteria and fungi community composition associated with Paulownia elongate × Paulownia fortune. BioResources 2019, 14, 8511–8529. [Google Scholar] [CrossRef]
- Bai, L.; Sun, H.; Zhang, X.; Cai, B. Next-generation sequencing of root fungal communities in continuous cropping soybean. Chil. J. Agric. Res. 2018, 78, 528–538. [Google Scholar] [CrossRef] [Green Version]
- Sommermann, L.; Geistlinger, J.; Wibberg, D.; Deubel, A.; Zwanzig, J.; Babin, D.; Schluter, A.; Schellenbern, I. Fungal community profiles in agricultural soils of a long-term field trial under different tillage, fertilization and crop rotation conditions analyzed by high-throughput ITS-amplicon sequencing. PLoS One 2018, 13, e0195345. [Google Scholar] [CrossRef] [PubMed]
- Schneider-Maunoury, L.; Deveau, A.; Moreno, M.; Todesco, F.; Belmondo, S.; Murat, C.; Courty, P.E.; Jąkalski, M.; Selosse, M.A. Two ectomycorrhizal truffles, Tuber melanosporum and T. aestivum, endophytically colonise roots of non-ectomycorrhizal plants in natural environments. New Phytol. 2020, 225, 2542–2556. [Google Scholar] [CrossRef]
- Zhang, X.; Li, X.; Ye, L.; Huang, Y.; Kang, Z.; Zhang, B. Colonization by Tuber melanosporum and Tuber indicum affects the growth of Pinus armandii and phoD alkaline phosphatase encoding bacterial community in the rhizosphere. Microbiol. Res. 2020, 239, 126520. [Google Scholar] [CrossRef] [PubMed]
- Zhang, X.; Ye, L.; Kang, Z.; Zou, J.; Zhang, X.; Li, X. Mycorrhization of Quercus acutissima with Chinese black truffle significantly altered the host physiology and root-associated microbiomes. Peer J. 2019, 7, e6421. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fu, Y.; Li, X.; Li, Q.; Wu, H.; Xiong, C.; Geng, Q.; Sun, H.; Sun, Q. Soil microbial communities of three major Chinese truffles in southwest China. Can. J. Microbiol. 2016, 62, 970–979. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Monaco, P.; Toumi, M.; Sferra, G.; Toth, E.; Naclerio, G.; Bucci, A. The bacterial communities of Tuber aestivum: Preliminary investigations in Molise region, southern Italy. Ann. Microbiol. 2010, 70, 37. [Google Scholar] [CrossRef]
- Splivallo, R.; Vahdatzadeh, M.; Maciá-Vicente, J.G.; Molinier, V.; Peter, M.; Egli, S.; Uroz, S.; Paolocci, F.; Deveau, A. Orchard Conditions and fruiting body characteristics drive the microbiome of the black truffle Tuber aestivum. Front. Microbiol. 2019, 10, 1437. [Google Scholar] [CrossRef] [PubMed]
- Antony-Babu, S.; Deveau, A.; Nostrand, J.D.V.; Zhou, J.; Tacon, F.L.; Robin, C.; Frey-Klett, P.; Uroz, S. Black truffle-associated bacterial communities during the development and maturation of Tuber melanosporum ascocarps and putative functional roles. Environ. Microbol. 2014, 16, 2831–2847. [Google Scholar] [CrossRef]
- Deveau, A.; Antony-Babu, S.; Tacon, L.F.; Robin, C.; Frey-Klett, P.; Uroz, S. Temporal changes of bacterial communities in the Tuber melanosporum ectomycorrhizosphere during ascocarp development. Mycorrhiza 2016, 26, 389–399. [Google Scholar] [CrossRef] [PubMed]
- Ye, L.; Li, Q.; Fu, Y.; Sourzat, P.; Tan, H.; Zou, J.; Zhang, B.; Li, X. Host species effects on bacterial communities associated with the ascocarps of Tuber species from the Sichuan Province in southwest China. Mycol. Prog. 2018, 17, 833–840. [Google Scholar] [CrossRef]
Figure 1.
Map showing the study area and position of soil sample collection of each target plant.

Figure 2.
Soil fungal composition of each sample at the phylum level.

Figure 3.
Soil fungal composition of each sample with the dominant of 35 fungal genera.

Figure 4.
Ascocarps (A) and ascospores (B) of Tuber thailadicum collected from the representative locations of the truffle OTU (B14). Scale bar: A = 1 cm and B = 25 µm.
Figure 4.
Ascocarps (A) and ascospores (B) of Tuber thailadicum collected from the representative locations of the truffle OTU (B14). Scale bar: A = 1 cm and B = 25 µm.

Figure 5.
Phylogram derived from the maximum likelihood tree revealed by RAxML from an analysis of an ITS sequence of 66 taxa. Tuber magnatum was used as outgroup. The numbers above the branches represent Bootstrap support (BS) (left) and PP (right). Only BS values ≥ 70% and PP ≥ 0.90 are shown. “-” indicates a value of BS and PP of less than 70% and 0.90, respectively. The scale bar represents the expected number of nucleotide substitutions per site. Sequences obtained in this study are in blue.
Figure 5.
Phylogram derived from the maximum likelihood tree revealed by RAxML from an analysis of an ITS sequence of 66 taxa. Tuber magnatum was used as outgroup. The numbers above the branches represent Bootstrap support (BS) (left) and PP (right). Only BS values ≥ 70% and PP ≥ 0.90 are shown. “-” indicates a value of BS and PP of less than 70% and 0.90, respectively. The scale bar represents the expected number of nucleotide substitutions per site. Sequences obtained in this study are in blue.


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Table 1.
Data of DNA sequences of Tuber species and fruiting body obtained from each soil sample.
| Sample No. * | Relative Abundant (%) † | OTU Number | Observation of Fruiting Body | |||||
|---|---|---|---|---|---|---|---|---|
| 105 | 162 | 187 | 447 | 530 | 720 | |||
| B1 | 0.02 | – | + | – | – | – | – | + |
| B2 * | 0.08 | – | + | – | – | – | – | + |
| B3 | 0.04 | – | + | – | – | – | + | – |
| B4 | 0.10 | – | + | – | – | – | – | + |
| B5 | 0.05 | – | + | – | – | – | – | + |
| B6 | 0.10 | – | + | – | – | – | – | – |
| B7 | 0.20 | + | + | – | – | – | – | + |
| B8 | 0.60 | – | – | – | – | – | + | – |
| B9 | 0.07 | + | – | – | – | – | – | – |
| B10 | 1.00 | – | + | – | – | – | – | + |
| B11 | 0.03 | – | + | – | – | – | – | – |
| B12 | 0.20 | – | + | – | + | – | – | + |
| B13 | 0.06 | – | + | – | – | – | – | + |
| B14 | 0.30 | – | + | – | – | – | – | + |
| B15 | 0.03 | – | + | – | – | – | – | + |
| B16 | 0.02 | – | + | – | – | – | – | – |
| B17 | 0.02 | + | – | – | – | – | – | – |
| B18 | 0.00 | – | – | – | – | – | – | – |
| B19 | 0.00 | – | – | – | – | – | – | – |
| B20 | 4.00 | – | + | – | + | – | – | – |
| B21 | 0.02 | – | – | – | – | – | + | – |
| B22 | 0.09 | – | + | – | – | – | – | + |
| B23 | 0.20 | – | + | – | – | – | – | – |
| B24 | 0.07 | – | – | – | – | – | + | – |
| B25 | 0.00 | – | – | – | – | – | – | – |
| B26 | 0.20 | – | + | – | – | – | – | + |
| B27 * | 0.90 | – | + | + | – | – | – | + |
| B28 | 0.03 | + | – | – | – | – | – | – |
| B29 | 0.04 | – | + | – | – | – | – | + |
| B30 | 0.00 | – | – | – | – | – | – | – |
| B31 | 0.03 | – | + | – | – | – | – | + |
| B32 | 0.10 | – | + | + | – | – | – | + |
| B33 | 0.03 | – | + | – | – | – | – | + |
| B34 | 0.02 | – | – | – | + | – | – | – |
| B35 | 0.00 | – | – | – | – | – | – | – |
| B36 | 0.80 | – | + | + | – | – | – | + |
| CP1 | 0.01 | – | – | + | – | – | – | – |
| CP2 * | 0.20 | – | – | + | – | + | – | – |
| Total found | 4 | 24 | 5 | 3 | 1 | 4 | 18 | |
* The previous known location. † Relative abundant of Tuber DNA was compared with the total fungal sequences in each sample. OTU162 = T. thailandicum, OTU187 = T. lannaense, OTU447 = T. bomiense, OTU530 = T. magnatum, OTU105 and OTU720 = unrecognized Tuber species. B = soil sample from Betula alnoides and CP = soil sample from Carpinus poilanei. “+” = presence/found and “–” = absence/not found.
Table 2.
Detail of the DNA sequences of Tuber species obtained from soil using metabarcoding.
| OTU Number | Length (bp) | GenBank Accession Number | Closeted Species/Accession Number | Similarity (%) |
|---|---|---|---|---|
| 105 | 182 | MW330007 | Tuber thailandicum CMU-MTUF001/KP196331 | 97.52 |
| 162 | 238 | MW326971 | Tuber thailandicum CMU-MTUF001/KP196332 | 100 |
| 187 | 228 | MW330257 | Tuber lanaense CMU-MTUF007/KT758731 | 100 |
| 447 | 211 | MW327594 | Tuber bomiense SKM101/KC517480 | 100 |
| 530 | 164 | MW326781 | Tuber magnatum Tmag_Ca_09/MG992596 | 100 |
| 720 | 168 | MW326083 | Tuber sp. isolate JT9769/HM485407 | 100 |
Table 3.
Morphological characteristics of truffles collected from each truffle OTU’s representative location in this study.
Table 3.
Morphological characteristics of truffles collected from each truffle OTU’s representative location in this study.
| Location | Macromorphological Characteristics | Micromorphological Characteristics | ||||||
|---|---|---|---|---|---|---|---|---|
| Ascomata | Hair-like Structure in Peridium Surface | Number of Ascospore/Ascus | Ascospores | |||||
| Size (cm in Diam) | Color | Gleba | Shape | Ornamentation | Size (μm) | |||
| B1 | 1.5–3.5 | White | Brown | + | 1–4 | Globose to subglobose | Alveolate-reticulate | 23–40 × 22–35 |
| B2 * | 2.2–4.3 | White to pale yellow | Brown | + | 1–4 | Globose to broadly ellipsoid | Alveolate-reticulate | 25–45 × 22–40 |
| B4 | 2.0 | White | Brown | + | 1–4 | Subglobose to broadly ellipsoid | Alveolate-reticulate | 20–35 × 18–28 |
| B5 | 1.0–3.0 | Pale yellow to light brown | Brown to dark brown | + | 1–4 | Subglobose to ellipsoid | Alveolate-reticulate | 25–55 × 22–48 |
| B7 | 2.5–3.0 | White to pale yellow | Brown | + | 1–4 | Globose to subglobose | Alveolate-reticulate | 28–40 × 25–38 |
| B10 | 1.5–3.5 | White to pale yellow | Brown | + | 1–4 | Subglobose | Alveolate-reticulate | 25–60 × 22–55 |
| B12 | 2.5–4.5 | Pale yellow to light brown | Dark brown | + | 1–4 | Subglobose to broadly ellipsoid | Alveolate-reticulate | 25–43 × 22–35 |
| B13 | 2.0 | Pale yellow | Brown | + | 1–4 | Globose to broadly ellipsoid | Alveolate-reticulate | 25–45 × 25–35 |
| B14 | 1.0–3.2 | White to pale yellow | Brown | + | 1–4 | Globose to subglobose | Alveolate-reticulate | 26–42 × 25–38 |
| B15 | 2.5 | White | Brown | + | 1–4 | Subglobose | Alveolate-reticulate | 20–50 × 18–47 |
| B22 | 2.5–4.0 | Pale yellow to light brown | Dark brown | + | 1–4 | Globose to subglobose | Alveolate-reticulate | 20–65 × 18–62 |
| B26 | 1.4–3.8 | White | Brown | + | 1–4 | Globose to subglobose | Alveolate-reticulate | 27–50 × 25–48 |
| B27 * | 1.0–3.2 | White to pale yellow | Dark brown | + | 1–4 | Globose to broadly ellipsoid | Alveolate-reticulate | 25–43 × 25–35 |
| B29 | 3.5 | White | Brown | + | 1–4 | Gubglobose to ellipsoid | Alveolate-reticulate | 25–50 × 22–40 |
| B31 | 2.0–4.1 | White to pale yellow | Brown | + | 1–5 | Globose to broadly ellipsoid | Alveolate-reticulate | 22–45 × 20–42 |
| B32 | 2.0–3.0 | Pale yellow to light brown | Brown to dark brown | + | 1–4 | Globose to subglobose | Alveolate-reticulate | 25–45 × 20–35 |
| B33 | 2.4–3.5 | White to pale yellow | Brown | + | 1–4 | Subglobose to ellipsoid | Alveolate-reticulate | 20–55 × 15–50 |
| B36 | 1.5–3.0 | White, pale yellow to light brown | Brown to dark brown | + | 1–4 | Subglobose to ellipsoid | Alveolate-reticulate | 30–50 × 22–45 |
* The previous known location. B = soil sample from Betula alnoides. “+” = presence.
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Suwannarach, N.; Kumla, J.; In-on, A.; Lumyong, S. Soil Metabarcoding Offers a New Tool for the Investigation and Hunting of Truffles in Northern Thailand. J. Fungi 2021, 7, 293. https://0-doi-org.brum.beds.ac.uk/10.3390/jof7040293
AMA Style
Suwannarach N, Kumla J, In-on A, Lumyong S. Soil Metabarcoding Offers a New Tool for the Investigation and Hunting of Truffles in Northern Thailand. Journal of Fungi. 2021; 7(4):293. https://0-doi-org.brum.beds.ac.uk/10.3390/jof7040293
Chicago/Turabian StyleSuwannarach, Nakarin, Jaturong Kumla, Ammarin In-on, and Saisamorn Lumyong. 2021. "Soil Metabarcoding Offers a New Tool for the Investigation and Hunting of Truffles in Northern Thailand" Journal of Fungi 7, no. 4: 293. https://0-doi-org.brum.beds.ac.uk/10.3390/jof7040293
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.