
Molecular Phylogeny of Endophytic Fungi from Rattan (Calamus castaneus Griff.) Spines and Their Antagonistic Activities against Plant Pathogenic Fungi

 ,
,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Sample Collection and Isolation of Endophytic Fungi
2.2. DNA Extraction and PCR Amplification
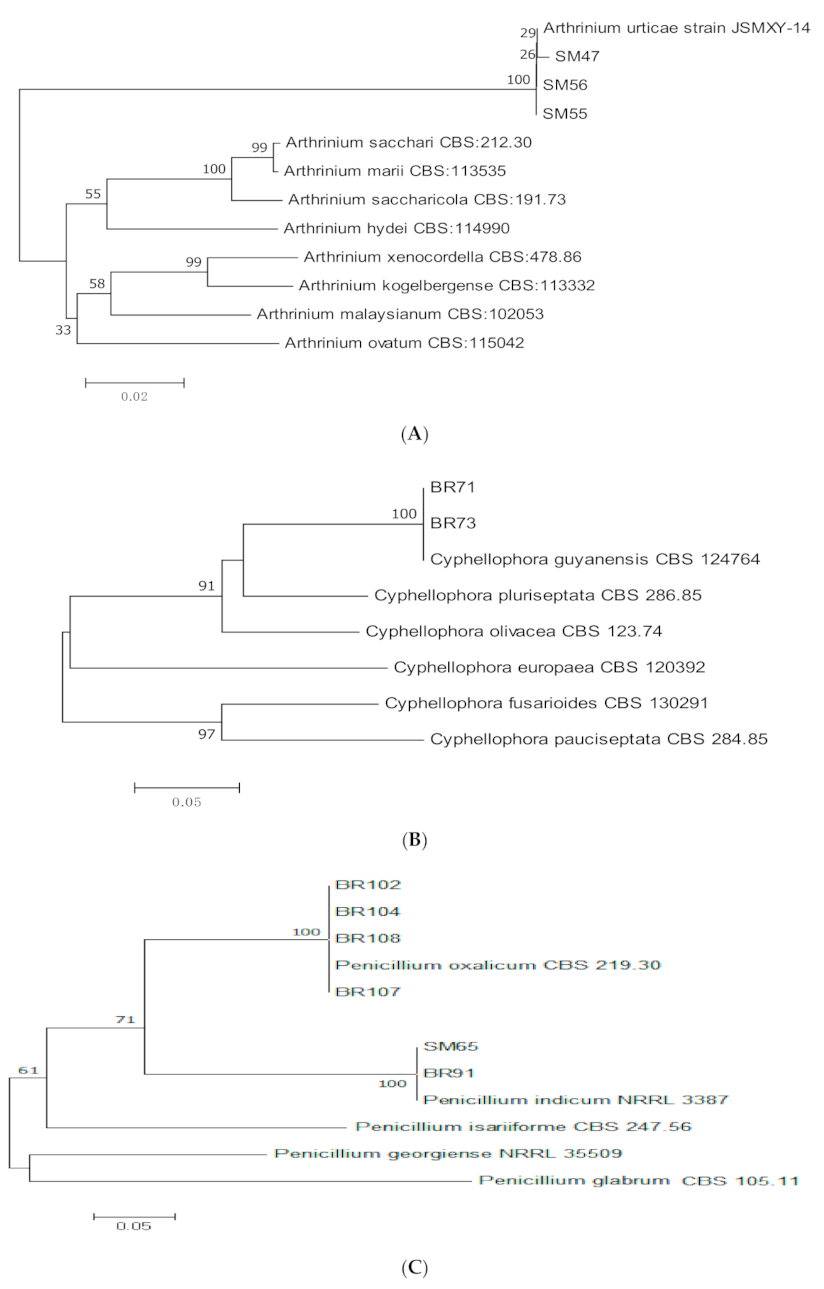
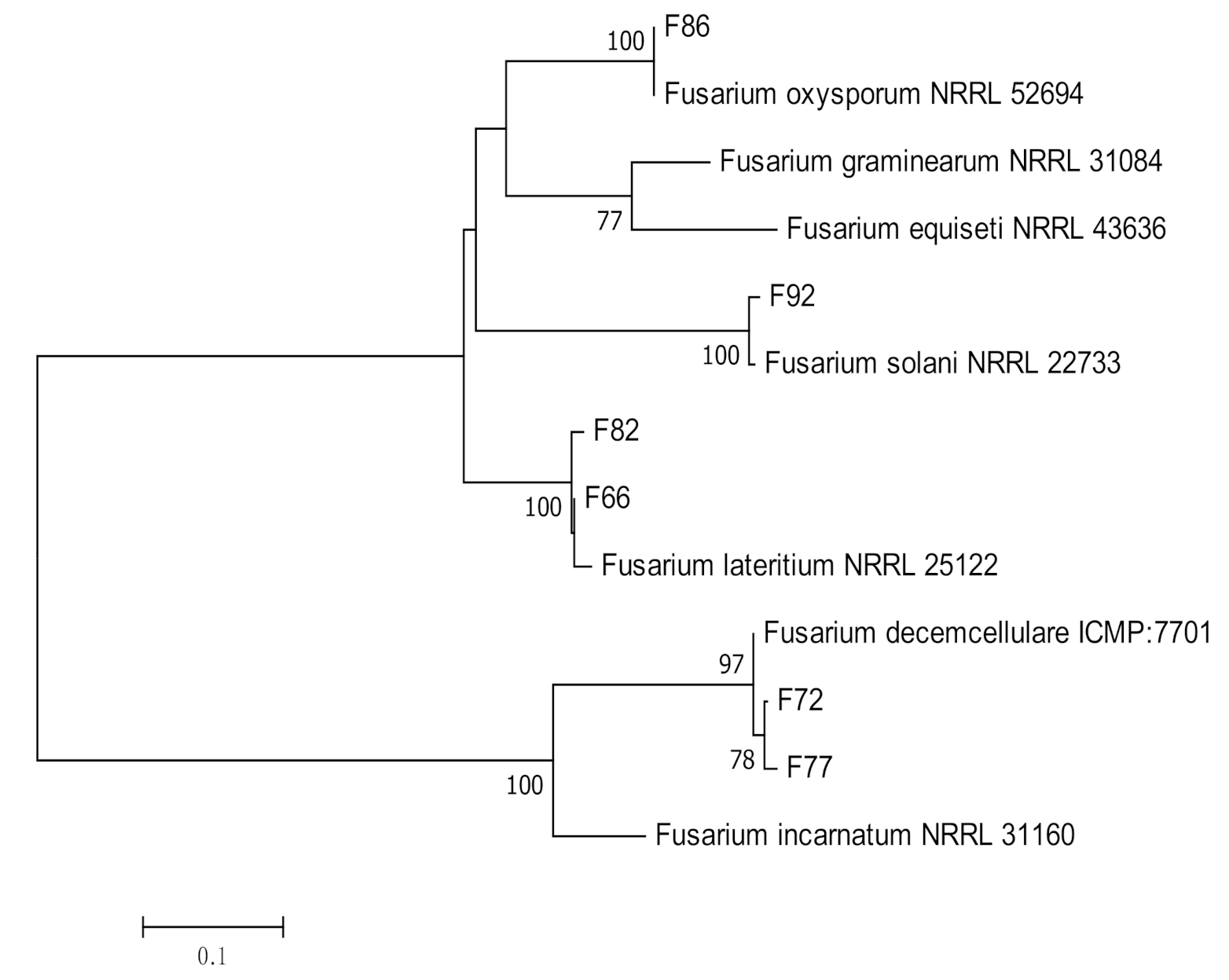
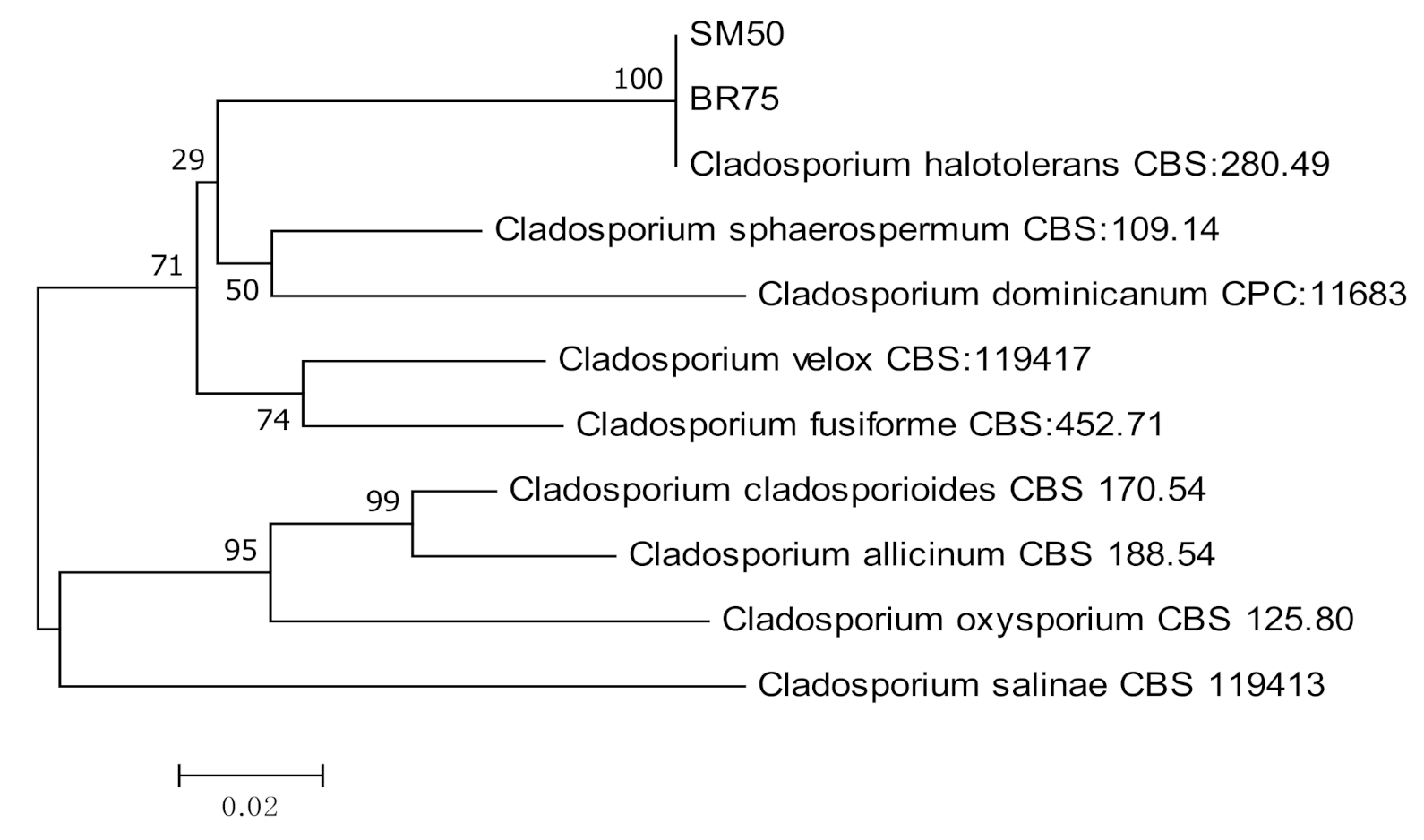
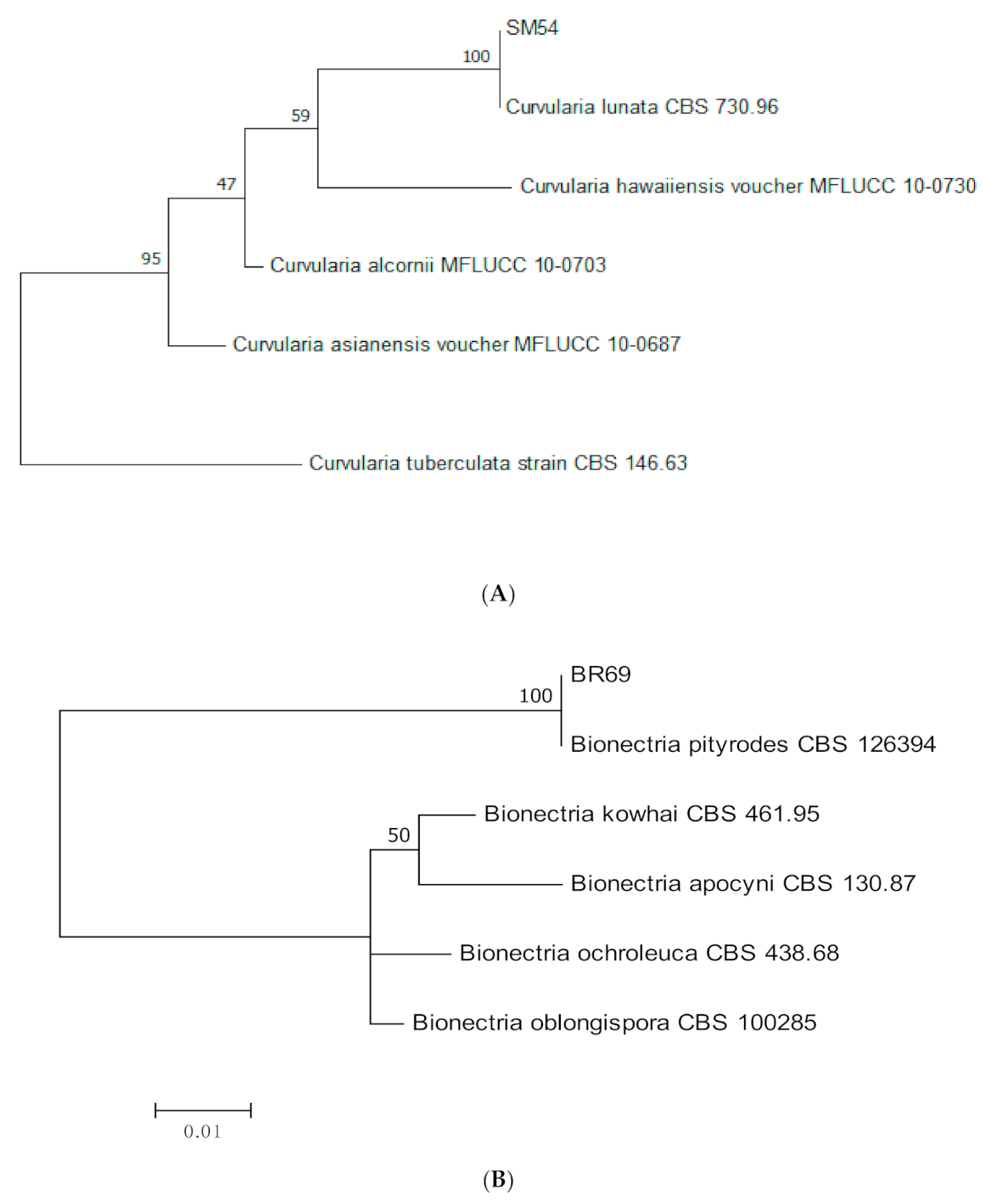
2.3. Molecular Identification and Phylogenetic Analysis
2.4. Antagonistic Activity
- R1—radial growth of plant pathogenic fungi in control plate.
- R2—radial growth of plant pathogenic fungi in dual culture plate.
3. Results
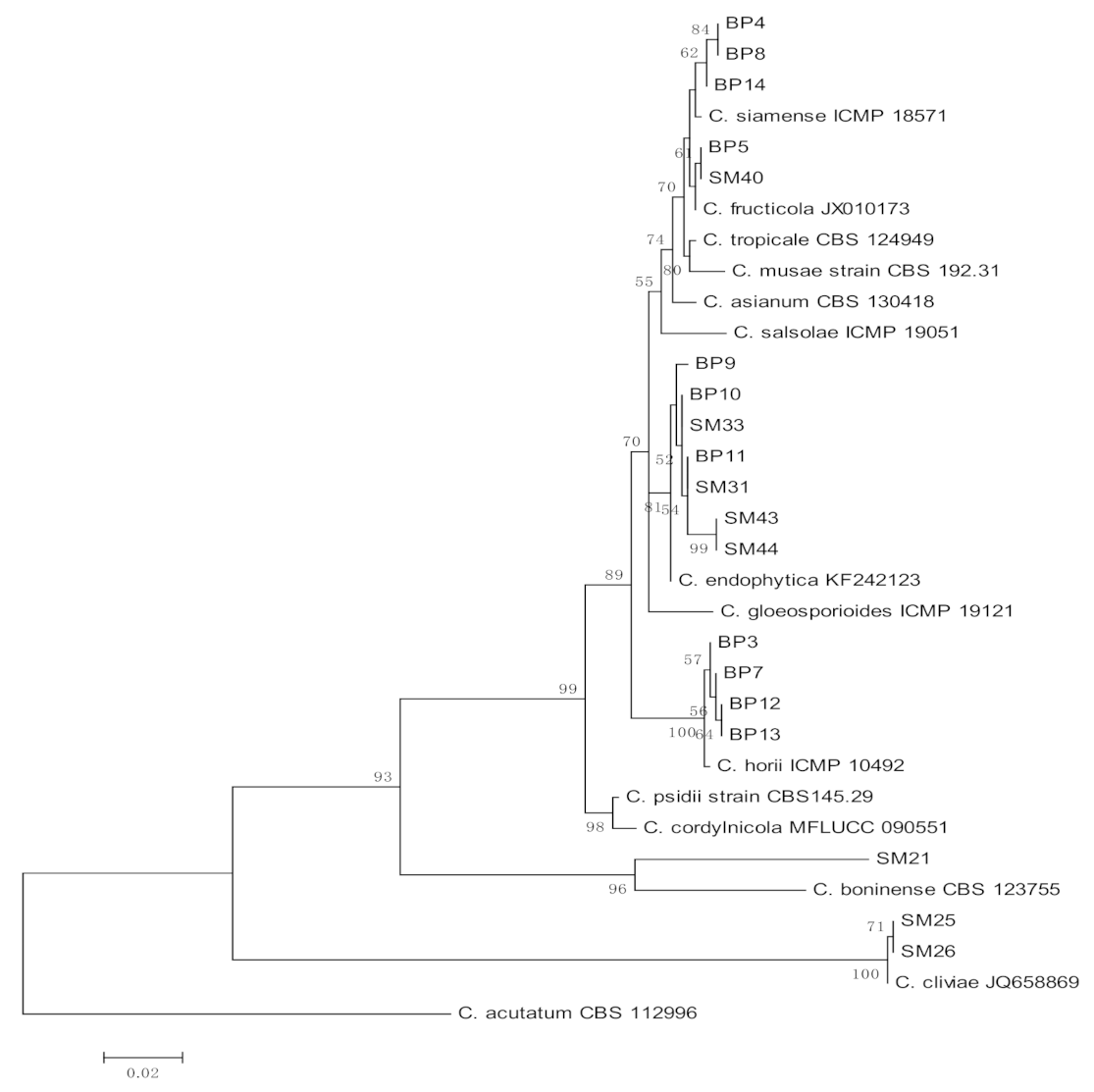
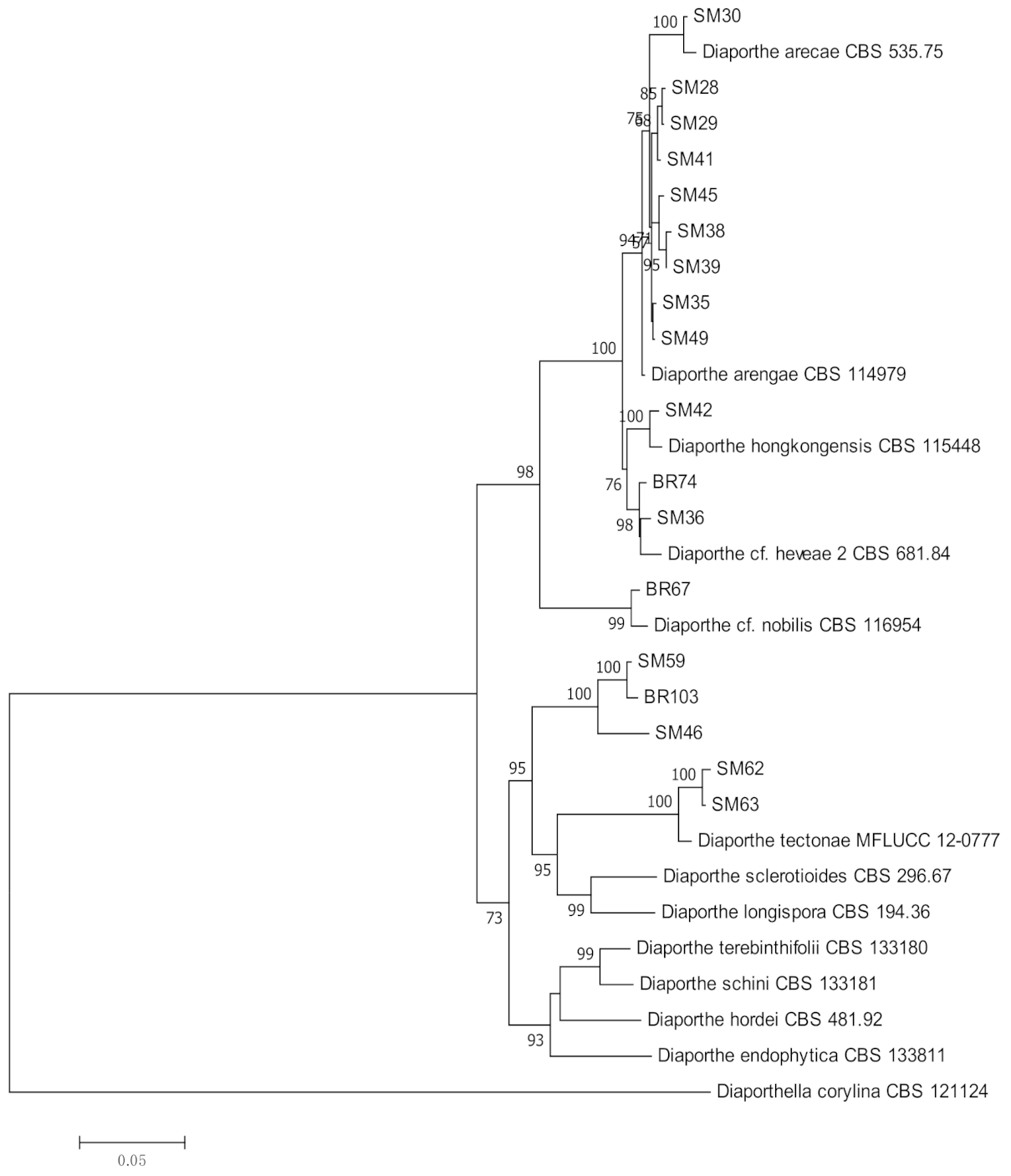
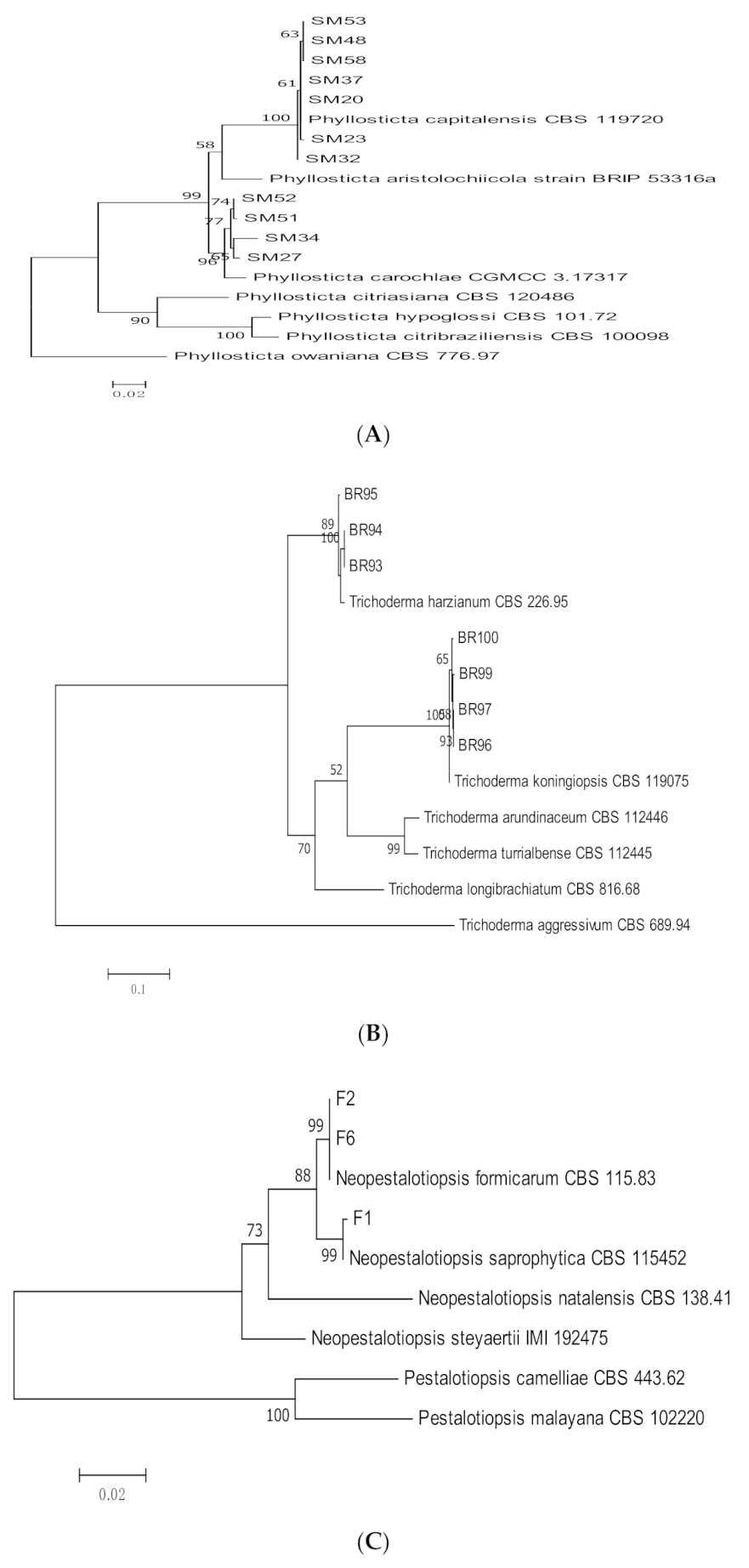
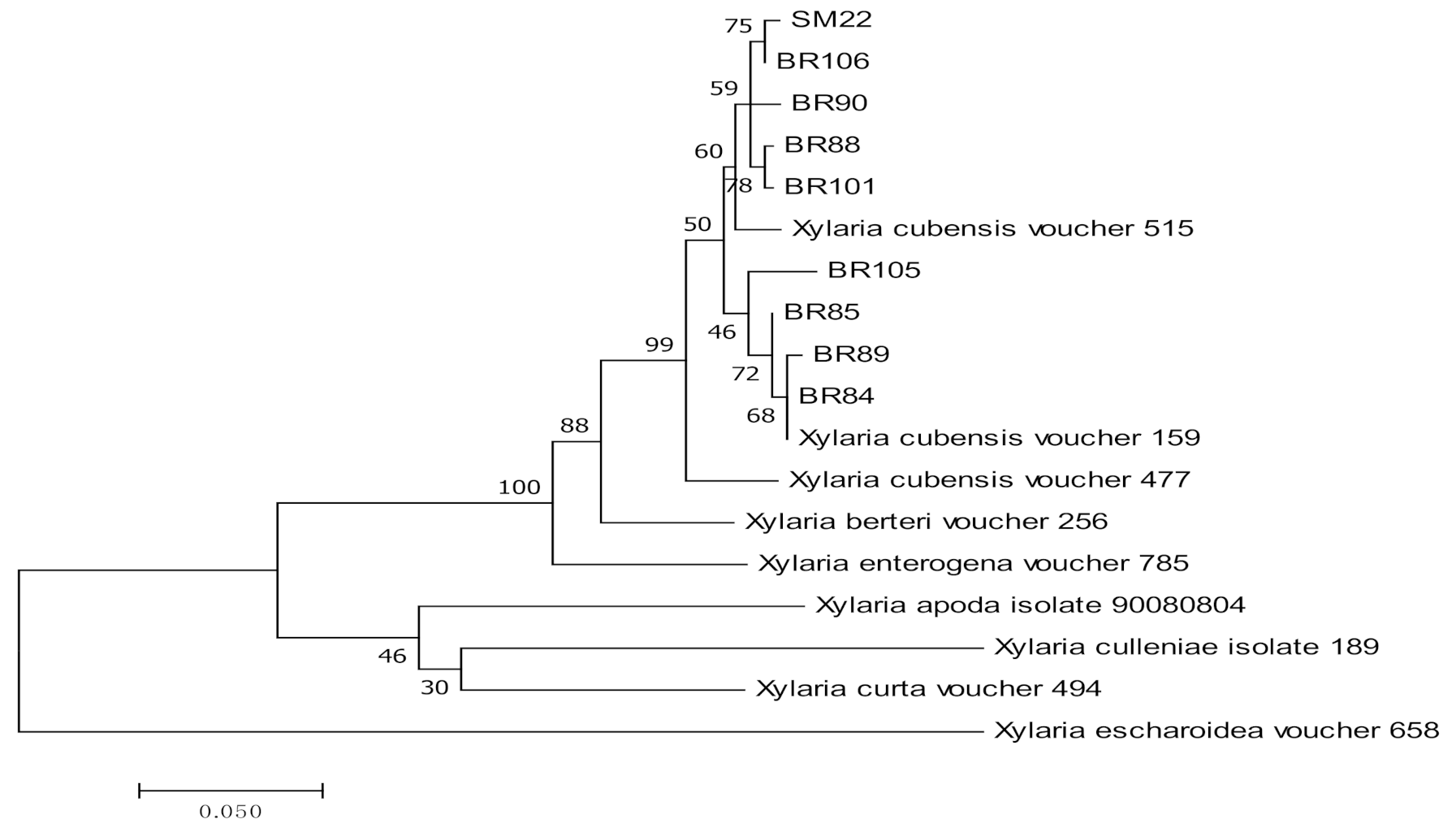
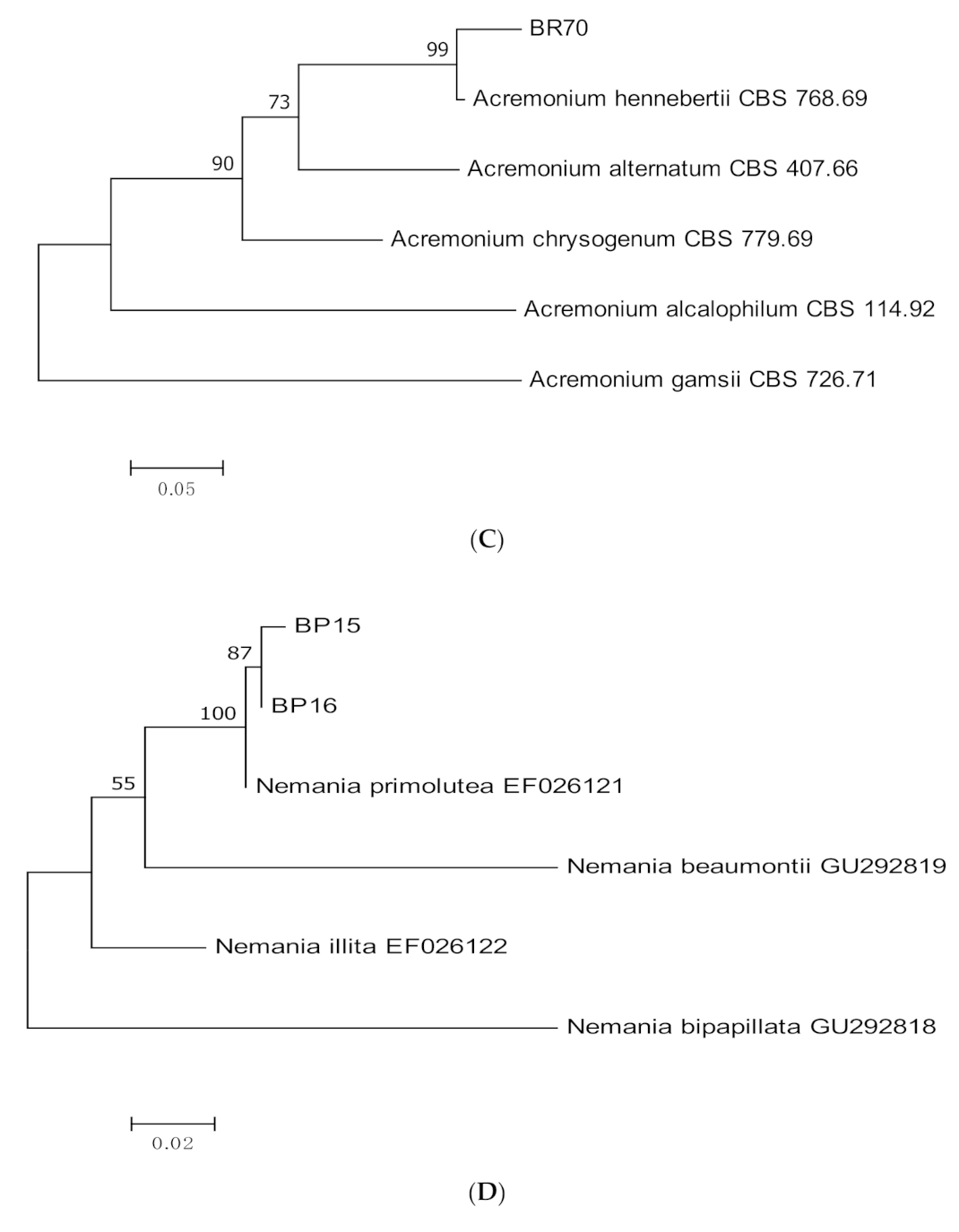
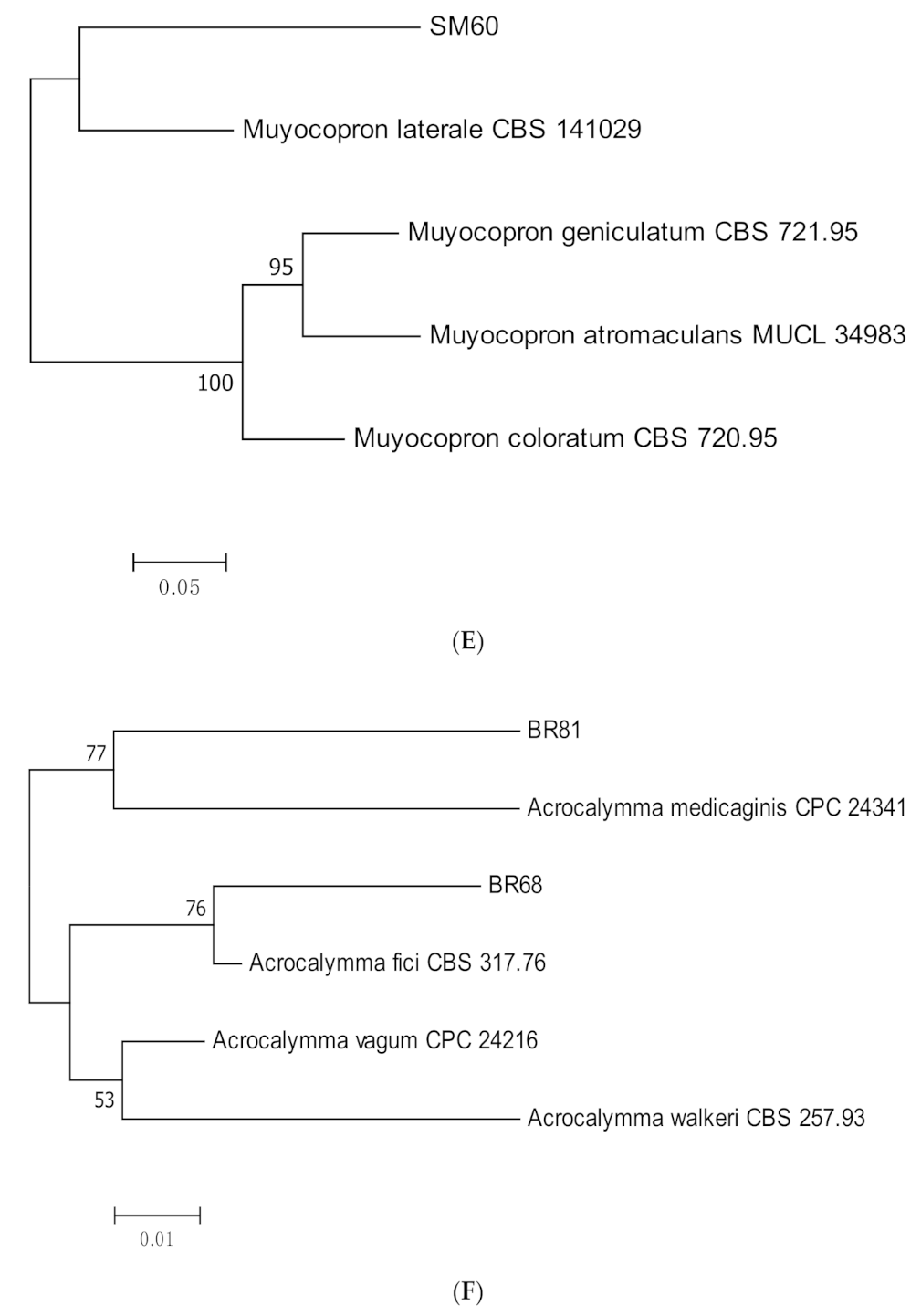
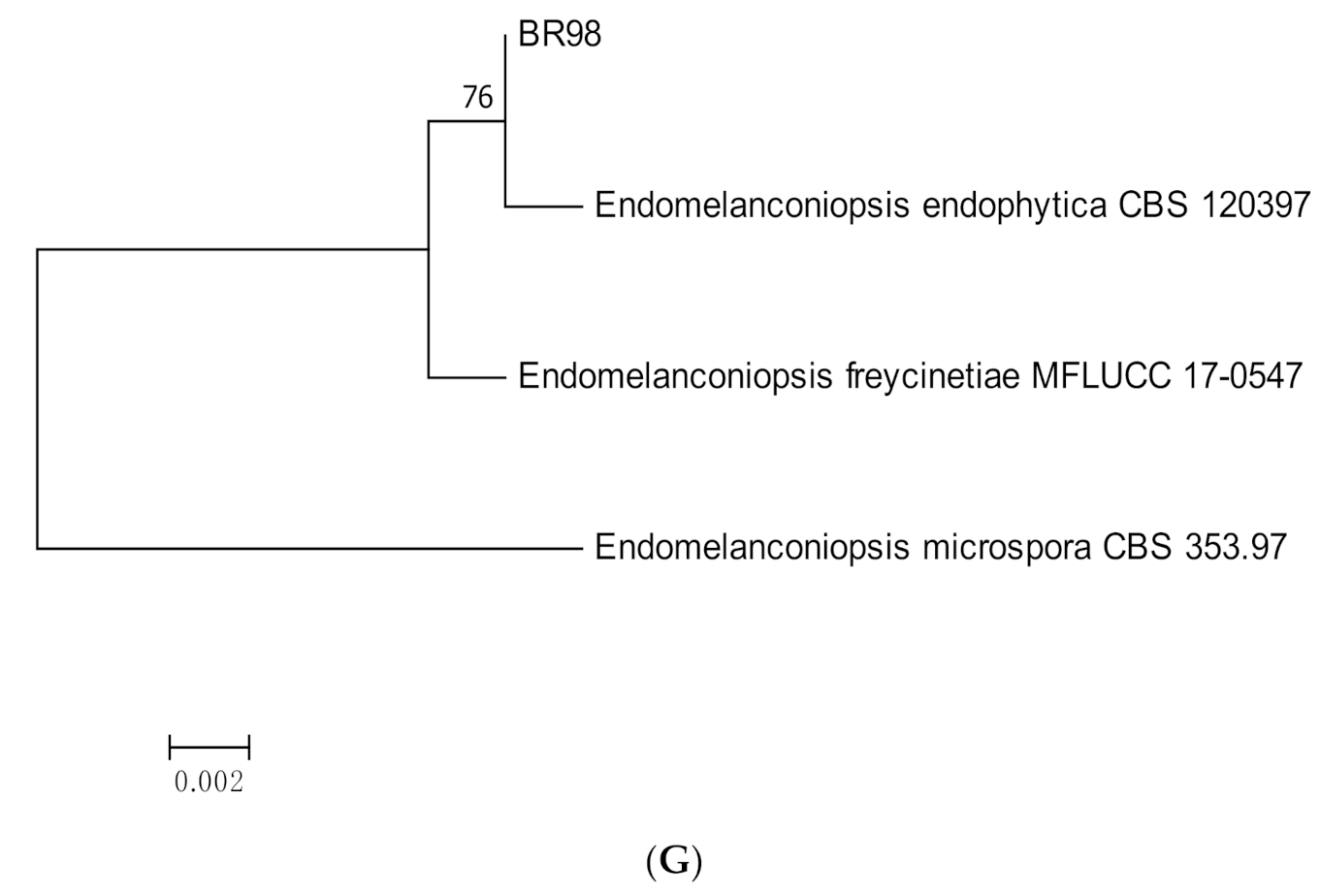
3.1. Molecular Identification
3.2. Antagonistic Activity
4. Discussion
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Schulz, B.; Boyle, C. The endophytic continuum. Mycol. Res. 2005, 109, 661–686. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Backman, P.A.; Sikora, R.A. Endophytes: An emerging tool for biological control. Biol. Control 2008, 46, 1–3. [Google Scholar] [CrossRef]
- Bilal, L.; Asaf, S.; Hamayun, M.; Gul, H.; Iqbal, A.; Ullah, I.; Lee, I.-J.; Hussaim, A. Plant growth promoting endophytic fungi Aspergillus fumigatus TS1 and Fusarium proliferatum BRL1 produce gibberellins and regulates plant endogenous hormones. Symbiosis 2018, 76, 117–127. [Google Scholar] [CrossRef]
- Arnold, A.E.; Mejía, L.C.; Kyllo, D.; Rojas, E.I.; Maynard, Z.; Robbins, N.; Herre, E.A. Fungal endophytes limit pathogen damage in a tropical tree. Proc. Natl. Acad. Sci. USA 2003, 100, 15649–15654. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mejia, L.C.; Rojas, E.I.; Maynard, Z.; Van Bael, S.; Arnold, A.E.; Hebbar, P.; Samuels, G.J.; Robbins, N.; Herre, E.A. Endophytic fungi as biocontrol agents of Theobroma cacao pathogens. Biol. Control 2008, 46, 4–14. [Google Scholar] [CrossRef]
- Gao, F.K.; Dai, C.C.; Liu, X.Z. Mechanisms of fungal endophytes in plant protection against pathogens. Afr. J. Microbiol. Res. 2010, 4, 1346–1351. [Google Scholar]
- Halpern, M.; Raats, D.; Lev-Yadun, S. Plant biological warfare: Thorns inject pathogenic bacteria into herbivores. Environ. Microbiol. 2007, 9, 584–592. [Google Scholar] [CrossRef]
- Dransfield, J.A. Manual of the rattans of the Malay Peninsula. In Malayan Forest Records 29; Forest Department, Ministry of Primary Industries: Kuala Lumpur, Malaysia, 1979. [Google Scholar]
- Halpern, M.; Waissler, A.; Dror, A.; Lev-Yadun, S. Biological warfare of the spiny plant: Introducing pathogenic microorganisms into herbivore’s tissues. In Advances in Applied Microbiology; Laskin, A.I., Bennett, J.W., Gadd, G.M., Eds.; Academic Press: Cambridge, MA, USA, 2011; Volume 74, pp. 97–116. [Google Scholar]
- White, T.; Bruns, T.; Lee, S.; Taylor, J. Amplification and direct sequencing of fungal ribosomal RNA genes for phylogenetics. In PCR Protocols: A Guide to Methods and Applications; Innis, M.A., Gelfand, D.H., Sninsky, J.J., White, T.J., Eds.; Academic Press: San Diego, CA, USA, 1990; pp. 315–322. [Google Scholar]
- Templeton, M.D.; Rikkerink, E.H.A.; Solon, S.L.; Crowhurst, R.N. Cloning and molecular characterization of the glyceraldehyde-3-phosphate dehydrogenase encoding gene and cDNA from the plant pathogenic fungus Glomerella cingulata. Gene 1992, 122, 225–230. [Google Scholar] [CrossRef]
- O’Donnell, K.; Kistlerr, H.C.; Cigelnik, E.; Ploetz, R.C. Multiple evolutionary origins of the fungus causing panama disease of banana: Concordant evidence from nuclear and mitochondrial gene genealogies. Proc. Natl. Acad. Sci. USA 1998, 95, 2044–2049. [Google Scholar] [CrossRef] [Green Version]
- Carbone, I.; Kohn, L.M. A method for designing primer sets for speciation studies infilamentous ascomycetes. Mycologia 1999, 91, 553–556. [Google Scholar] [CrossRef]
- O’Donnell, K.; Cigelnik, E. Two divergent intragenomic rDNA ITS2 types withina monophyletic lineage of the fungus Fusarium are nonorthologous. MoI. Phylogenet. Evol. 1997, 7, 103–116. [Google Scholar] [CrossRef]
- Glass, N.L.; Donaldson, G.C. Development of primer sets designed for use with the PCR to amplify conserved genes from filamentous ascomycetes. Appl. Environ. Microbiol. 1995, 61, 1323–1330. [Google Scholar] [CrossRef] [Green Version]
- Vilgalys, R.; Hester, M. Rapid genetic identification and mapping of enzymatically amplified ribosomal DNA from several Cryptococcus species. J. Bacteriol. 1990, 172, 4238–4246. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- De Hoog, G.S.; Gerrits Van Den Ende, A.H.G. Molecular diagnostics of clinical strains of filamentous Basidiomycetes. Mycoses 1998, 189, 183–189. [Google Scholar] [CrossRef] [PubMed]
- Kumar, S.; Stecher, G.; Tamura, K. MEGA7: Molecular evolutionary genetics analysis version 7.0 for bigger datasets. Mol. Biol. Evol. 2016, 33, 1870–1874. [Google Scholar] [CrossRef] [Green Version]
- Skidmore, A.M.; Dickinson, C.H. Colony interactions and hyphal interference between Septoria nodorum and phylloplane fungi. Trans. Brit. Mycol. Soc. 1976, 66, 57–64. [Google Scholar] [CrossRef]
- Schulz, B.; Wanke, U.; Draeger, S.; Aust, H.-J. Endophytes from herbaceous plants and shrubs: Effectiveness of surface sterilization methods. Mycol. Res. 1993, 97, 1447–1450. [Google Scholar] [CrossRef]
- Sánchez Márquez, S.; Bills, G.F.; Zabalgogeazcoa, I. The endophytic mycobiota of the grass Dactylis glomerata. Fungal Diver. 2007, 27, 171–195. [Google Scholar]
- Cai, L.; Hyde, K.D.; Taylor, P.W.; Weir, B.S.; Waller, J.M.; Abang, M.M.; Zhang, J.Z.; Yang, Y.L.; Phoulivong, S.; Liu, Z.Y.; et al. A polyphasic approach for studying. Colletotrichum Fungal Divers 2009, 39, 183–204. [Google Scholar]
- Weir, B.S.; Johnston, P.R.; Damm, U. The Colletotrichum gloeosporioides species complex. Stud Mycol. 2012, 73, 115–180. [Google Scholar] [CrossRef] [Green Version]
- Schoch, C.L.; Seifert, K.A.; Huhndorf, S.; Robert, V.; Spouge, J.L.; Levesque, C.A.; Chen, W. Fungal Barcoding Consortium. 2012. Nuclear ribosomal internal transcribed spacer (ITS) region as a universal DNA barcode marker for Fungi. Proc. Natl. Acad. Sci. USA 2012, 109, 6241–6246. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Stielow, J.B.; Lévesque, C.A.; Seifert, K.A.; Meyer, W.; Iriny, L.; Smits, D.; Renfurm, R.; Verkley, G.J.; Groenewald, M.; Chaduli, D.; et al. One fungus, which genes? Development and assessment of universal primers for potential secondary fungal DNA barcodes. Persoonia 2015, 35, 242–263. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ferreira, M.C.; de Vieira, M.L.A.; Zani, C.L.; de Alves, T.M.A.; Junior PA, S.; Murta SM, F.; Rosa, L.H. Molecular phylogeny, diversity, symbiosis and discover of bioactive compounds of endophytic fungi associated with the medicinal Amazonian plant Carapa guianensis Aublet (Meliaceae). Biochem. Syst. Ecol. 2015, 59, 36–44. [Google Scholar] [CrossRef]
- Guo, L.D.; Hyde, K.D.; Liew, E.C.Y. Identification of endophytic fungi from Livistona chinensis (Palmae) using morphological and molecular techniques. New Phytol. 2000, 147, 617–630. [Google Scholar] [CrossRef]
- Song, J.; Pongnak, W.; Soytong, K. Isolation and identification of endophytic fungi from 10 species palm trees. J. Agric. Technol. 2016, 12, 349–363. [Google Scholar]
- Prihastuti, H.; Cai, L.; Chen, H.; McKenzie, E.H.C.; Hyde, K.D. Characterization of Colletotrichum species associated with coffee berries in northern Thailand. Fungal Divers. 2009, 39, 89–109. [Google Scholar]
- Zhang, H.W.; Song, Y.C.; Tan, R.X. Biology and chemistry of endophytes. Nat. Prod. Rep. 2006, 23, 753–771. [Google Scholar] [CrossRef] [PubMed]
- Carraro, L.; Maifreni, M.; Bartolomeoli, I.; Martino, M.E.; Novelli, E.; Frigo, F.; Marino, M.; Cardazzo, B. Comparison of culture-dependent and -independent methods for bacterial community monitoring during Montasio cheese manufacturing. Res. Microbiol. 2011, 162, 231–239. [Google Scholar] [CrossRef]
- Nocker, A.; Burr, M.; Camper, K. Genotypic microbial community profiling: A critical technical review. Microb. Ecol. 2007, 54, 276–289. [Google Scholar] [CrossRef]
- Vaz-Moreira, I.; Egas, C.; Nunes, O.C.; Manaia, C.M. Culture-dependent and culture-independent diversity surveys target different bacteria: A case study in a freshwater sample. Anton Leeuw. 2011, 100, 245–257. [Google Scholar] [CrossRef]
- Shokralla, S.; Spall, J.L.; Gibson, J.F.; Hajibabaei, M. Next-generation sequencing technologies for environmental DNA research. Mol. Ecol. 2012, 21, 1794–1805. [Google Scholar] [CrossRef]
- Rajamani, T.; Suryanarayanan, T.S.; Murali, T.S.; Thirunavukkarasu, N. Distribution and diversity of foliar endophytic fungi in the mangroves of Andaman Islands, India. Fungal Ecol. 2018, 36, 109–116. [Google Scholar] [CrossRef]
- Santos, C.M.; Ribeiro, A.S.; Garcia, A.; Polli, A.D.; Polonio, J.C.; Azevedo, J.L.; Pamphile, J.A. Enzymatic and antagonist activity of endophytic fungi from Sapindus saponaria L. (Sapindaceae). Acta Biol. Colomb. 2019, 24, 322–330. [Google Scholar] [CrossRef]
- Sun, X.; Guo, L.D.; Hyde, K.D. Community composition of endophytic fungi in Acer truncatum and their role in decomposition. Fungal Divers. 2011, 47, 85–95. [Google Scholar] [CrossRef]
- Kirilenko, S.T. Pidoplitchkoviella terricola–a new ascomycete. Mikrobiol. Zhurnal 1975, 37, 603–605, [in Ukrainian with English summary]. [Google Scholar]
- Nováková, A. Pidoplitchkoviella terricola–an interesting fungus from the Domica Cave (Slovakia). Int. J. Speleology 2009, 38, 23–26. [Google Scholar] [CrossRef] [Green Version]
- Halpern, M.; Raats, D.; Lev-Yadun, S. The potential anti-herbivory role of microorganisms on plant thorns. Plant Signal. Behav. 2007, 2, 503–504. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lo’pez-Martı´nez, R.; Me´ndez-Tovar, L.J. Chromoblastomycosis. Clin. Dermatol. 2007, 25, 188–194. [Google Scholar] [CrossRef]
- Son, Y.-M.; Kang, H.-K.; Na, S.-Y.; Lee, H.-Y.; Baek, J.-O.; Lee, J.-R.; Roh, J.-Y.; Seo, Y.-H. Chromoblastomycosis caused by Phialophora Richardsiae. Ann. Dermatol. 2010, 22, 362–366. [Google Scholar] [CrossRef]
- Salgado, C.G.; da Silva, J.P.; Diniz, J.A.; da Silva, M.B.; da Costa, P.F.; Teixeira, C.; Salgado, U.I. Isolation of Fonsecaea pedrosoi from thorns of Mimosa pudica, aprobable natural source of chromoblastomycosis. Rev. Inst. Med. Trop. Sa˜o Paulo 2004, 46, 33–36. [Google Scholar] [CrossRef] [Green Version]
- Engle, J.; Desir, J.; Bernstein, J.M. A rose by any other name. Skinmed 2007, 6, 139–141. [Google Scholar] [CrossRef]
- Haldar, N.; Sharma, M.K.; Gugnani, H.C. Sporotrichosis in north-east India. Mycoses 2007, 50, 201–204. [Google Scholar] [CrossRef]
- Arnold, A.E. Understanding the diversity of foliar fungal endophytes: Progress, challenges, and frontiers. Fungal Biol. Rev. 2007, 21, 51–66. [Google Scholar] [CrossRef]
- Vega, F.E.; Simpkins, A.; Aime, M.C.; Posada, F.; Peterson, S.W.; Rehner, S.A.; Infante, F.; Castillo, A.; Arnold, A.E. Fungal endophyte diversity in coffee plants from Colombia, Hawai’i, Mexico and Puerto Rico. Fungal Ecol. 2010, 3, 122–138. [Google Scholar] [CrossRef]
- Porras, M.; Barrau, C.; Romero, F. Biological control of anthracnose with Trichoderma in strawberry fields. Acta Hortic. 2009, 842, 351–354. [Google Scholar] [CrossRef]
- Alvindia, D.G. The antagonistic action of Trichoderma harzianum strain DGA01 against anthracnose-causing pathogen in mango cv.‘Carabao’. Biocontrol Sci. Technol. 2018, 28, 591–602. [Google Scholar] [CrossRef]
- Tsegaye Redda, E.; Ma, J.; Mei, J.; Li, M.; Wu, B.; Jiang, X. Antagonistic potential of different Isolates of Trichoderma against Fusarium oxysporum, Rhizoctonia solani, and Botrytis cinerea. Eur. J. Exp. Biol. 2018, 8, 1–8. [Google Scholar] [CrossRef]
- Kong, P.; Hong, C. Biocontrol of boxwood blight by Trichoderma koningiopsis Mb2. Crop Prot. 2017, 98, 124–127. [Google Scholar] [CrossRef]
- Ghanbarzadeh, B.; Safaie, N.; Goltapeh, E.M. Antagonistic activity and hyphal interactions of Trichoderma spp. against Fusarium proliferatum and F. oxysporum in vitro. Arch. Phytopathol. Pflanzenschutz 2014, 47, 1979–1987. [Google Scholar] [CrossRef]
- Taha Yassin, M.; Abdel-Fattah Mostafa, A.; Al-Askar, A.A.; Sayed, S.R.M.; Mostafa Rady, A. Antagonistic activity of Trichoderma harzianum and Trichoderma viride strains against some fusarial pathogens causing stalk rot disease of maize, in vitro. J. King Saud Univ. Sci. 2021, 33, 101363. [Google Scholar]
- Ben Amira, M.; Lopez, D.; Triki Mohamed, A.; Khouaja, A.; Chaar, H.; Fumanal, B.; Venisse, J.S. Beneficial effect of Trichoderma harzianum strain Ths97 in biocontrolling Fusarium solani causal agent of root rot disease in olive trees. Biol. Control 2017, 110, 70–78. [Google Scholar] [CrossRef]
- Milanesi, P.M.; Blume, E.; Antonioli, Z.I.; Muniz, M.F.B.; Santos, R.F.; dos Finger, G.; Durigon, M.R. Biocontrol of Fusarium spp. with Trichoderma spp. and growth promotion in soybean seedlings. Rev. Ciênc. Agrár. 2013, 36, 347–356. [Google Scholar]
- Verma, M.; Brar, S.K.; Tyagi, R.D.; Surampalli, R.Y.; Valéro, J.R. Antagonistic fungi, Trichoderma spp.: Panoply of biological control. Biochem. Eng. J. 2007, 37, 1–20. [Google Scholar] [CrossRef]
- Sudarma, I.M.; Puspawati, N.M.; Suada, I.K. The potency of endofit fungi in cocoa as biological agent to control cocoa pod disease caused by Phytophthota palmivora (Butler) Butler. Adv. Trop. Biodivers. Environ. Sci. 2017, 1, 6–10. [Google Scholar] [CrossRef] [Green Version]
- Khare, E.; Mishra, J.; Arora, N.K. Multifaceted interactions between endophytes and plant: Developments and prospects. Front. Microbiol. 2018, 9, 1–12. [Google Scholar] [CrossRef] [PubMed]
- Tan, R.X.; Zau, W.X. Endophytes: A rich source of functional metabolites. Nat. Prod. Rep. 2001, 18, 448–459. [Google Scholar] [CrossRef]
- Strobel, G.A.; Daisy, B.; Castillo, U.; Harper, J. Natural products from endophytic fungi. J. Nat. Prod. 2004, 67, 257–268. [Google Scholar] [CrossRef]
- Zhang, P.; Li, X.M.; Liu, H.; Li, X.; Wang, B.G. Two new alkaloids from Penicillium oxalicum EN-201, an endophytic fungus derived from the marine mangrove plant Rhizophora stylosa. Phytochem. Lett. 2015, 13, 160–164. [Google Scholar] [CrossRef]
- Guo, B.; Wang, Y.; Sun, X.; Tang, K. Bioactive natural products from endophytes: A review. Appl. Biochem. Microbiol. 2008, 44, 136–142. [Google Scholar] [CrossRef]
- Chen, L.H.; Cui, Y.Q.; Yang, X.M.; Zhao, D.K.; Shen, Q.R. An antifungal compound from Trichoderma harzianum SQR-T037 effectively controls Fusarium wilt of cucumber in continuously cropped soil. Australas. Plant Pathol. 2012, 41, 239–245. [Google Scholar] [CrossRef]
- Scarselletti, R.; Faull, J.L. In vitro activity of 6-pentyl-a-pyrone, a metabolite of Trichoderma harzianum, in the inhibition of Rhizoctonia solani and Fusarium oxysporum f. sp. lycopersici. Mycol. Res. 1994, 98, 1207–1209. [Google Scholar] [CrossRef]
- Deshmukh, S.K.; Gupta, M.K.; Prakash, V.; Saxena, S. Endophytic Fungi: A source of potential antifungal compounds. J. Fungi 2018, 4, 77. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nisa, H.; Kamili, A.N.; Nawchoo, I.A.; Shafi, S.; Shameem, N.; Bandh, S.A. Fungal endophytes as prolific source of phytochemicals and other bioactive natural products: A review. Microb. Pathog. 2015, 82, 50–59. [Google Scholar] [CrossRef] [PubMed]
- Manganyi, M.C.; Ateba, C.N. Untapped potentials of endophytic fungi: A review of novel bioactive compounds with biological applications. Microorganisms 2020, 8, 1934. [Google Scholar] [CrossRef] [PubMed]
- Xu, T.-C.; Lu, Y.-H.; Wang, J.-F.; Song, Z.-Q.; Hou, Y.-G.; Liu, S.-S.; Liu, C.-S.; Wu, S.-H. Bioactive secondary metabolites of the genus diaporthe and anamorph phomopsis from terrestrial and marine habitats and endophytes: 2010–2019. Microorganisms 2021, 9, 217. [Google Scholar] [CrossRef] [PubMed]















{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Region/Gene | Primers | Sequence (5′-3′) | Fungal Genera | References |
|---|---|---|---|---|
| ITS | ITS 1 | TCC GTA GGT GAA CCT GCG G | All fungal genera | White et al. (1990) [10] |
| ITS 4 | TCC GCT TAT TGA TAT GC | |||
| GAPDH | GDF1 | GCC GTC AAC GAC CCC TTC ATT GA | Colletotrichum spp. | Templeton et al. (1992) [11] |
| GDR2 | GGG TGG AGT CGT ACT TGA GCA TGT | |||
| TEF-1α | EF1 | ATG GGT AAG GAG GAC AAG AC | Fusarium spp. | |
| EF2 | GGA AGT ACC AGT GAT CAT GTT | |||
| EF1-728F | CAT CGA GAA GTT CGA GAA GG | Diaporthe spp. | O’Donnell et al. (1998) [12] | |
| EF1-986R | TAC TTG AAG GAA CCC TTA CC | |||
| EF1-728F | CAT CGA GAA GTT CGA GAA GG | Phyllosticta spp. | Carbone and Kohn (1999) [13] | |
| EF2 | GGA AGT ACC AGT GAT CAT GTT | Arthrinium sp. | ||
| Pestalotiopsis spp. | ||||
| EF1-728F | CAT CGA GAA GTT CGA GAA GG | Trichoderma spp. | ||
| TEF1-rev | GCC ATC CTT GGA GAT ACC AGC | |||
| β-tubulin | T1 | AAC ATG CGT GAG ATT GTA AGT | Xylaria sp. | |
| T22 | TCT GGA TGT TGG GAA TCC | |||
| T1 | AAC ATG CGT GAG ATT GTA AGT | Fusarium spp. | O’Donnell and Cigelnik (1997) [14] | |
| T2 | TAG TGA CCC TTG GCC CAG TTG | |||
| Bt2a | GGT AAC CAA ATC GGT GCT TTC | Penicillium spp. | Glass and Donaldson (1995) [15] | |
| Bt2b | ACC CTC AGT GTA GTG ACC CTT GGC | |||
| T1 | AAC ATG CGT GAG ATT GTA AGT | Cyphellophora sp. | ||
| Bt2b | ACC CTC AGT GTA GTG ACC CTT GGC | Diaporthe spp. | ||
| ACT | ACT-512F | ATG TGC AAG GCC GGT TTC G | Xylaria sp. | Carbone and Kohn (1999) [13] |
| ACT-783R | TAC GAG TCC TTC TGG CCC AT | Cladosporium sp. | ||
| LSU | LROR | ACC CGC TGA ACT TAA GC | Non-sporulating fungi | Vilgalys and Hester (1990) [16] |
| LR5 | TCC TGA GGG AAA CTT CG | |||
| V9G | TTA CGT CCC TGC CCT TTG TA | Corynespora spp. | De Hoog and Gerrits Van Den Ende (1998) [17] | |
| LR5 | TCC TGA GGG AAA CTT CG | Vilgalys and Hester (1990) [16] |
| Endophytic Fungi | |||||||||
|---|---|---|---|---|---|---|---|---|---|
| Plant Pathogenic Fungi | C. endophytica (BP9) | C. siamense (BP14) | C. boninense (SM21) | X. cubensis (SM22) | X. cubensis (BR90) | D. arengae (SM45) | D. tectonae (BR62) | D. cf. nobilis (BR67) | D. cf. heveae (BR74) |
| C.truncatum | √ | √ | √ | √ | √ | √ | √ | √ | √ |
| C. scovellei | √ | √ | √ | √ | √ | √ | √ | √ | √ |
| F. solani | √ | √ | √ | √ | √ | √ | √ | √ | √ |
| F. oxysporum | √ | √ | √ | √ | √ | √ | √ | √ | √ |
| F. proliferatum | √ | √ | √ | √ | √ | √ | √ | √ | √ |
| F. fujikuroi | √ | √ | √ | √ | √ | √ | √ | √ | √ |
| L. theobromae | √ | √ | √ | √ | √ | √ | √ | √ | √ |
| P. mangiferae | √ | √ | √ | √ | √ | √ | √ | √ | √ |
| L. pseudotheobromae | √ | √ | √ | √ | √ | √ | √ | √ | √ |
| D. pascoei | √ | √ | √ | √ | √ | √ | √ | √ | √ |
| Plant Pathogenic Fungi | Endophytic Fungi | ||||||||
| N. saprophytica (BP1) | Pen. indicum (BR91) | T. harzianum (BR94) | T. koningiopsis (BR96) | End. endophytica (BR98) | Pen.oxalicum (BR102) | ||||
| C.truncatum | √ | √ | √ | √ | √ | √ | |||
| C. scovellei | √ | √ | √ | √ | √ | √ | |||
| F. solani | √ | √ | √ | √ | √ | √ | |||
| F. oxysporum | √ | √ | √ | √ | √ | √ | |||
| F. proliferatum | √ | √ | √ | √ | √ | √ | |||
| F. fujikuroi | √ | √ | √ | √ | √ | √ | |||
| L. theobromae | √ | √ | √ | √ | √ | √ | |||
| P. mangiferae | √ | √ | √ | √ | √ | √ | |||
| L. pseudotheobromae | √ | √ | √ | √ | √ | √ | |||
| D. pascoei | √ | √ | √ | √ | √ | √ | |||
| Genbank Accession Number | |||||||
|---|---|---|---|---|---|---|---|
| Isolates | ITS | GAPDH | β-Tubulin | TEF-1α | ACT | LSU | % Similarity |
| Colletotrichum spp. | |||||||
| C. siamense BP4 | MN635697 | MT077122 | - | - | - | - | 99 |
| C. siamense BP8 | MN635698 | MT077123 | - | - | - | - | 99 |
| C. siamense BP14 | MN635699 | MT077124 | - | - | - | - | 99 |
| C. fructicola BP5 | MN635702 | MT077113 | - | - | - | - | 99 |
| C. fructicola SM40 | MN635702 | MT077114 | - | - | - | - | 99 |
| C. endophytica BP9 | MN635726 | MT077115 | - | - | - | - | 99 |
| C. endophytica BP10 | MN635727 | MT077116 | - | - | - | - | 99 |
| C. endophytica BP11 | MN635728 | MT077117 | - | - | - | - | 99 |
| C. endophytica SM31 | MN635729 | MT077118 | - | - | - | - | 99 |
| C. endophytica SM33 | MN635730 | MT077119 | - | - | - | - | 99–100 |
| C. endophytica SM43 | MN635731 | MT077120 | - | - | - | - | 99 |
| C. endophytica SM44 | MN635732 | MT077121 | - | - | - | - | 99 |
| C. horii BP3 | MN635649 | MT077107 | - | - | - | - | 99 |
| C. horii BP7 | MN635650 | MT077108 | - | - | - | - | 99 |
| C. horii BP12 | MN635651 | MT077109 | - | - | - | - | 99 |
| C. horii BP13 | MN635652 | MT077110 | - | - | - | - | 99 |
| C. cliviae SM25 | MN652631 | MT077111 | - | - | - | - | 99 |
| C. cliviae SM26 | MN652632 | MT077112 | - | - | - | - | 99 |
| C. boninense SM21 | MN635733 | MT077106 | - | - | - | - | 99 |
| Diaporthe spp. | |||||||
| D. arengae SM28 | MN651480 | - | MT077062 | MT077093 | - | - | 98–99 |
| D. arengae SM41 | MN651481 | - | MT077064 | MT077095 | - | - | 98–99 |
| D. arengae SM35 | MN651483 | - | MT077068 | MT077099 | - | - | 98–99 |
| D. arengae SM49 | MN651487 | - | MT077069 | MT077089 | - | - | 98–99 |
| D. arengae SM38 | MN651484 | - | MT077066 | MT077097 | - | - | 98–99 |
| D. arengae SM39 | MN651485 | - | MT077067 | MT077098 | - | - | 98–99 |
| D. arengae SM45 | MN635732 | - | MT077065 | MT077096 | - | - | 97–98 |
| D. arengae SM29 | MN651486 | - | MT077063 | MT077094 | - | - | 98–99 |
| D. arecae SM30 | MN651482 | - | MT077061 | MT077090 | - | - | 99 |
| D. hongkongensis SM42 | MN651488 | - | MT077085 | MT077103 | - | - | 97–99 |
| D. cf. heveae SM36 | MN651489 | - | MT077080 | MT077092 | - | - | 96–99 |
| D. cf. heveae BR74 | MN636282 | - | MT077079 | MT077091 | - | - | 96–99 |
| D. cf. nobilis BR67 | MN651491 | - | MT077084 | MT077088 | - | - | 96–98 |
| Diaporthe sp.SM46 | MN651495 | - | MT077083 | MT077100 | - | - | 98–99 |
| Diaporthe sp. SM59 | MN651496 | - | MT077081 | MT077101 | - | - | 95–99 |
| Diaporthe sp. BR103 | MN651497 | - | MT077082 | MT077102 | - | - | 98–99 |
| D. tectonae SM62 | MN651493 | - | MT077086 | MT077104 | - | - | 95–97 |
| D. tectonae SM63 | MN651494 | - | MT077087 | MT077105 | - | - | 95–98 |
| Phyllosticta spp. | |||||||
| P. capitalensis SM20 | MN635748 | - | - | MT118281 | - | - | 99 |
| P. capitalensis SM23 | MN635749 | - | - | MT118282 | - | - | 99 |
| P. capitalensis SM32 | MN635750 | - | - | MT118283 | - | - | 99–100 |
| P. capitalensis SM37 | MN635751 | - | - | MT118284 | - | - | 99–100 |
| P. capitalensis SM48 | MN635752 | - | - | MT118285 | - | - | 99 |
| P. capitalensis SM53 | MN635753 | - | - | MT118286 | - | - | 99 |
| P. capitalensis SM58 | MN635754 | - | - | MT118287 | - | - | 99 |
| P. carochlae SM27 | MN652663 | - | - | MT118272 | - | - | 99 |
| P. carochlae SM34 | MN652664 | - | - | MT118269 | - | - | 95–99 |
| P. carochlae SM51 | MN652665 | - | - | MT118270 | - | - | 97–99 |
| P. carochlae SM52 | MN652666 | - | - | MT118271 | - | - | 97–99 |
| Neopestalatiopsis spp. | |||||||
| N. saprophytica BP1 | MN635619 | - | - | MT264943 | - | - | 99 |
| N. formicarum BP2 | MN635621 | - | - | MT264929 | - | - | 99 |
| N. formicarum BP6 | MN635622 | - | - | MT264930 | - | - | 99 |
| Trichoderma spp. | |||||||
| T. harzianum BR93 | MN636262 | - | - | MT264931 | - | - | 99–100 |
| T. harzianum BR94 | MN636263 | - | - | MT264932 | - | - | 99 |
| T. harzianum BR95 | MN636264 | - | - | MT264933 | - | - | 98–99 |
| T. harzianum BR93 | MN636262 | - | - | MT264931 | - | - | 99–100 |
| T. koningiospsis BR96 | MN636269 | - | - | MT264934 | - | - | 99 |
| T. koningiospsis BR97 | MN636270 | - | - | MT264935 | - | - | 99 |
| T. koningiospsis BR99 | MN636271 | - | - | MT264936 | - | - | 99 |
| T.koningiospsis BR100 | MN636272 | - | - | MT264937 | - | - | 99 |
| Xylaria cubensis | |||||||
| X. cubensis SM22 | - | - | MT118273 | - | MT077070 | - | 99 |
| X. cubensis BR84 | - | - | MT118274 | - | MT077071 | - | 99 |
| X. cubensis BR85 | - | - | MT118275 | - | MT077072 | - | 99 |
| X. cubensis BR88 | - | - | MT118276 | - | MT077073 | - | 99 |
| X. cubensis BR89 | - | - | MT118277 | - | MT077074 | - | 99 |
| X. cubensis BR90 | - | - | MT118278 | - | MT077075 | - | 99 |
| X. cubensis BR101 | - | - | MT118279 | - | MT077076 | - | 99 |
| X. cubensis BR105 | - | - | MT118280 | - | MT077077 | - | 99 |
| X. cubensis BR106 | - | - | - | - | MT077078 | - | 95–99 |
| Pidoplitchkoviella terricola | |||||||
| Pid. terricola SM17 | MN652667 | - | - | - | - | MW338725 | 96 |
| Pid. terricola SM18 | MN652668 | - | - | - | - | MW338726 | 96 |
| Pid. terricola SM19 | MN652669 | - | - | - | - | MW338727 | 96 |
| Pid. terricola SM24 | MN652670 | - | - | - | - | MW338728 | 96 |
| Pid. terricola SM57 | MN652671 | - | - | - | - | MW338729 | 96 |
| Pid. terricola BR79 | MN652672 | - | - | - | - | MW338730 | 96 |
| Helminthosporium spp. | |||||||
| H. endiandrea SM61 | MT279339 | - | - | - | - | MW338667 | 99 |
| H. endiandrea SM64 | MT279340 | - | - | - | - | MW338668 | 99 |
| H. livistonae BR76 | MN652658 | - | - | - | - | MW338703 | 93–97 |
| H. livistonae BR78 | MN652659 | - | - | - | - | MW338704 | 93–98 |
| H. livistonae BR80 | MN652660 | - | - | - | - | MW338705 | 93–99 |
| H. livistonae BR83 | MN652673 | - | - | - | - | MW338706 | 93–99 |
| H. livistonae BR87 | MT279326 | - | - | - | - | MW338669 | 99–100 |
| Cladosporium halotolerans | |||||||
| Cla. halotolerans SM50 | MN636281 | - | - | - | MT264919 | - | 99 |
| Cla. halotolerans BR75 | MN636282 | - | - | - | MT264920 | - | 99 |
| Penicillium spp. | |||||||
| Pen. indicum SM65 | MN635766 | - | MT264923 | - | - | - | 99 |
| Pen. indicum BR91 | MN635767 | - | MT264924 | - | - | - | 99 |
| Pen. oxalicum BR102 | MN636265 | - | MT264925 | - | - | - | 99 |
| Pen. oxalicum BR104 | MN636266 | - | MT264926 | - | - | - | 99 |
| Pen. oxalicum BR107 | MN636267 | - | MT264927 | - | - | - | 99 |
| Pen. oxalicum BR108 | MN636268 | - | MT264928 | - | - | - | 99 |
| Fusarium spp. | |||||||
| F. lateritium BR66 | - | - | MT296784 | MT264940 | - | - | 99–100 |
| F. decemcellulare BR72 | - | - | MT296782 | MT264938 | - | - | 99 |
| F. decemcellulare BR77 | - | - | MT296783 | MT264939 | - | - | 99 |
| F. lateritium BR82 | - | - | MT296785 | MT264941 | - | - | 99 |
| F. oxysporum BR86 | - | - | MT296786 | MT264942 | - | - | 99 |
| F. solani BR92 | - | - | MT296787 | MT264944 | - | - | 99 |
| Cyphellophora guyanensis | |||||||
| Cyp. guyanensis BR71 | MN636279 | - | MT264921 | - | - | - | 99–100 |
| Cyp. guyanensis BR73 | MN636280 | - | MT264922 | - | - | - | 99 |
| Arthrinium urticae | |||||||
| Art. urticae SM47 | MN636276 | - | - | - | - | - | 98–99 |
| Art. urticae SM55 | MN636277 | - | - | - | - | - | 98–99 |
| Art. urticae SM56 | MN636278 | - | - | - | - | - | 99 |
| Nemania primolutea | |||||||
| Nem.primolutea BP15 | MN652661 | - | - | - | - | - | 99 |
| Nem.primolutea BP16 | MN652662 | - | - | - | - | - | 99 |
| Cuvularia. lunata SM54 | MN637803 | - | - | - | - | - | 99 |
| Muyocopron laterale SM60 | MN637806 | - | - | - | - | - | 96 |
| Endomelanconiopsis endophytica BR98 | MN637809 | - | - | - | - | - | 99 |
| Acrocalymma fici BR68 | MN637807 | - | - | - | - | - | 96 |
| Acrocalymma medicaginis BR81 | MN637808 | - | - | - | - | - | 96 |
| Acremonium hennebertii BR70 | MN637805 | - | - | - | - | - | 99 |
| Bionectria pityrodes BR69 | MN637804 | - | - | - | - | - | 99 |
| Endophytic Fungi and PGI Value | |||||||||
|---|---|---|---|---|---|---|---|---|---|
| Plant Pathogenic Fungi | C. endophytica (BP9) | C. siamense (BP14) | C. boninense (SM21) | X. cubensis (SM22) | X. cubensis (BR90) | D. arengae (SM45) | D. tectonae (BR62) | D. cf. nobilis (BR67) | D. cf. heveae(BR74) |
| C. truncatum | 33.33 ± 6.03 cd | 13.33 ± 5.58 ab | 20.46 1.38 bc | 0 ± 0.00 a | 1.11 ± 1.72 a | 15.24 ± 14.53 ab | 45.49 ± 4.04 d | 19.57 ± 0.70 bc | 38.34 ± 2.40 d |
| C. scovellei | 19.52 ± 0.56 abc | 55.85 ± 3.27 cd | 28.10 ± 6.24 bcd | 0.57 ± 1.73 a | 1.33 ± 2.37 a | 57.73 ± 4.05 cd | 70.59 ± 3.51 f | 30.55 ± 0.15 bcd | 60.50 ± 5.47 de |
| F. solani | 35.96 ± 2.15 de | 31.58 ± 1.66 d | 20.18 ± 2.72 bc | 13.16 ± 0.66 a | 13.16 ± 2.35 a | 16.23 ± 1.98 ab | 41.23 ± 2.72 e | 17.54 ± 2.15 ab | 35.53 ± 2.20 de |
| F. oxysporum | 28.47 ± 0.69 abc | 49.65 ± 1.57 ef | 33.33 ± 1.32 bc | 26.39 ± 1.70 ab | 20.49 ± 2.77 a | 60.76 ± 2.05 fg | 61.35 ± 1.66 g | 34.72 ± 2.85 bc | 57.99 ± 4.04 ef |
| F. proliferatum | 28.58 ± 4.01 bc | 16.10 ± 0.86 abc | 4.45 ± 3.01 a | 4.80 ± 3.93 a | 6.85 ± 3.88 ab | 17.11 ± 15.18 abc | 40.45 ± 17.79 cd | 14.97 ± 8.87 abc | 19.50 4.35 abc |
| F. fujikuroi | 41.90 ± 2.76 cd | 33.64 ± 8.95 abc | 28.44 ± 1.64 a | 27.83 ± 1.50 a | 25.99 ± 3.96 a | 48.93 ± 1.80 def | 55.35 ± 6.10 ef | 40.06 ± 2.76 bcd | 45.8 ± 1.53 de |
| L. theobromae | 58.20 ± 5.22 ef | 40.23 ± 2.50 bc | 50.10 ± 1.00 bcde | 55.85 ± 11.90 def | 57.40 ± 4.55 ef | 38.49 ± 2.86 b | 0 ± 0.00 a | 0 ± 0.00 a | 0 ± 0.00 a |
| Pes. mangiferae | 27.78 ± 2.33 de | 31.48 ± 1.67 ef | 22.22 ± 1.99 c | 22.59 ± 1.67 c | 22.22 ± 1.99 c | 27.04 ± 1.67 d | 44.07 ± 1.67 g | 29.63 ± 2.30 def | 33.33 ± 1.99 f |
| L. pseudotheobromae | 43.56 ± 1.38 cd | 42.89 ± 1.00 cd | 40.22 ± 1.00 b | 0.00 ± 0.00 a | 0.00 ± 0.00 a | 43.56 ± 1.38 cd | 56.44 ± 1.38 g | 44.44 ± 1.38 d | 47.78 ± 1.00 e |
| D. pascoei | 39.42 ± 31 abc | 38.00 ± 4.88 abc | 31.30 ± 2.64 ab | 27.82 ± 2.40 a | 29.56 ± 1.46 ab | 32.75 ± 4.35 abc | 38.55 ± 2.38 abc | 36.23 ± 2.38 abc | 39.13 ± 3.65 abc |
| Plant Pathogenic Fungi | Endophytic Fungi and PGI Value | ||||||||
| N. saprophytica (BP1) | Pen. indicum (BR91) | T. harzianum (BR94) | T. koningiopsis (BR96) | End. endophytica (BR98) | Pen. oxalicum (BR102) | ||||
| C. truncatum | 19.44 ± 2.51 bc | 7.22 ± 2.51 ab | 89.33 ± 2.99 e | 80.05 ± 5.75 e | 53.65 ± 10.85 d | 1.34 ± 2.33 a | |||
| C. scovellei | 48.46 ± 8.00 cd | 3.20 ± 4.66 a | 85.80 ± 5.47 e | 89.45 ± 2.55 e | 45.70 ± 7.39 bcd | 8.09 ± 2.13 ab | |||
| F. solani | 35.96 ± 2.15 de | 16.67 ± 11.39 ab | 62.28 ± 2.15 f | 74.56 ± 2.72 g | 24.56 ± 2.72 c | 25.44 ± 2.15 c | |||
| F. oxysporum | 46.88 ± 1.14 de | 30.56 ± 7.65 abc | 76.74 ± 4.45 h | 76.04 ± 1.74 h | 59.03 ± 5.38 fg | 37.85 ± 1.57 cd | |||
| F. proliferatum | 30.18 ± 8.98 bcd | 7.94 ± 7.11 ab | 57.38 ± 17.22 e | 51.63 ± 13.52 de | 23.52 ± 8.66 abc | 11.36 ± 6.34 abc | |||
| F. fujikuroi | 43.43 ± 6.19 cd | 32.42 ± 5.37 abc | 71.25 ± 1.50 g | 59.94 ± 11.16 fg | 46.18 ± 1.50 de | 30.28 ± 1.64 ab | |||
| L. theobromae | 43.07 ± 2.89 abc | 46.83 ± 0.89 abcd | 82.86 ± 1.28 f | 77.62 ± 6.30 f | 63.59 ± 4.83 e | 48.85 ± 3.89 abcd | |||
| Pes. mangiferae | 27.41 ± 2.30 cd | 7.41 ± 3.04 a | 88.89 ± 1.41 g | 60.00 ± 1.99 f | 32.52 ± 1.89 de | 14.07 ± 2.69 b | |||
| L. pseudotheobromae | 52.44 ± 1.09 f | 41.56 ± 1.00 bc | 73.78 ± 1.09 h | 93.56 ± 1.00 i | 53.56 ± 1.00 f | 44.44 ± 1.09 d | |||
| D. pascoei | 53.04 ± 6.22 d | 39.71 ± 1.809 bc | 66.96 ± 1.56 e | 66.67 ± 9.30 e | 44.35 ± 1.10 cd | 39.71 ± 1.809 bc | |||
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Azuddin, N.F.; Mohd, M.H.; Rosely, N.F.N.; Mansor, A.; Zakaria, L. Molecular Phylogeny of Endophytic Fungi from Rattan (Calamus castaneus Griff.) Spines and Their Antagonistic Activities against Plant Pathogenic Fungi. J. Fungi 2021, 7, 301. https://0-doi-org.brum.beds.ac.uk/10.3390/jof7040301
Azuddin NF, Mohd MH, Rosely NFN, Mansor A, Zakaria L. Molecular Phylogeny of Endophytic Fungi from Rattan (Calamus castaneus Griff.) Spines and Their Antagonistic Activities against Plant Pathogenic Fungi. Journal of Fungi. 2021; 7(4):301. https://0-doi-org.brum.beds.ac.uk/10.3390/jof7040301
Chicago/Turabian StyleAzuddin, Nurul Farizah, Masratul Hawa Mohd, Nik Fadzly N. Rosely, Asyraf Mansor, and Latiffah Zakaria. 2021. "Molecular Phylogeny of Endophytic Fungi from Rattan (Calamus castaneus Griff.) Spines and Their Antagonistic Activities against Plant Pathogenic Fungi" Journal of Fungi 7, no. 4: 301. https://0-doi-org.brum.beds.ac.uk/10.3390/jof7040301